
The G Protein-Coupled Glutamate Receptors as Novel Molecular Targets in Schizophrenia Treatment—A Narrative Review



Abstract
1. Introduction
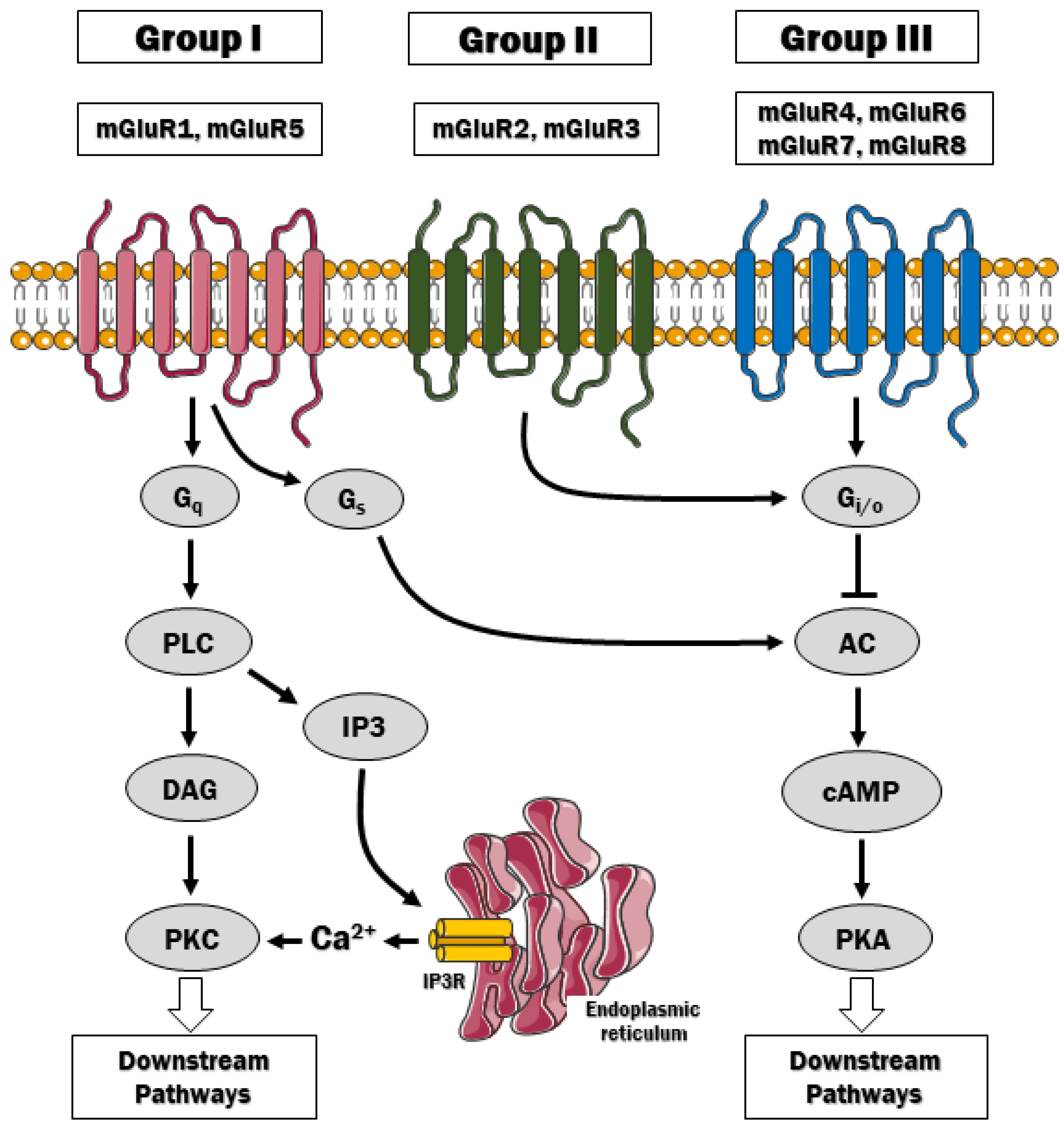
2. Metabotropic Glutamate Receptors (mGluRs)—An Overview
3. Group I mGluRs
3.1. Biology of Group I mGluRs
3.2. mGluR1
3.3. mGluR5
4. Group II mGluRs
4.1. General Characteristics
4.2. mGluR2/3 Function in Schizophrenia
4.3. Targeting mGluR2/3 in Schizophrenia Treatment
4.3.1. Preclinical Studies
4.3.2. Clinical Trials
4.3.3. Advantages and Disadvantages of Clinical Use of mGluR2/3 Agonists
5. Group III mGluRs
5.1. General Characteristics
5.2. mGluR4
Targeting mGluR4 in Preclinical Models of Schizophrenia
5.3. mGluR7
Targeting mGluR7 in Preclinical Models of Schizophrenia
5.4. mGluR8
Targeting mGluR8 in Preclinical Models of Schizophrenia
6. Concluding Remarks
Author Contributions
Funding
Conflicts of Interest
References
- Charlson, F.J.; Ferrari, A.J.; Santomauro, D.F.; Diminic, S.; Stockings, E.; Scott, J.G.; McGrath, J.J.; Whiteford, H.A. Global Epidemiology and Burden of Schizophrenia: Findings from the Global Burden of Disease Study 2016. Schizophr. Bull. 2018, 44, 1195–1203. (In English) [Google Scholar] [CrossRef] [PubMed]
- Keefe, R.S.; Eesley, C.E.; Poe, M.P. Defining a cognitive function decrement in schizophrenia. Biol. Psychiatry 2005, 57, 688–691. (In English) [Google Scholar] [CrossRef] [PubMed]
- Lally, J.; MacCabe, J.H. Antipsychotic medication in schizophrenia: A review. Br. Med. Bull. 2015, 114, 169–179. (In English) [Google Scholar] [CrossRef]
- Huhn, M.; Nikolakopoulou, A.; Schneider-Thoma, J.; Krause, M.; Samara, M.; Peter, N.; Arndt, T.; Bäckers, L.; Rothe, P.; Cipriani, A.; et al. Comparative efficacy and tolerability of 32 oral antipsychotics for the acute treatment of adults with multi-episode schizophrenia: A systematic review and network meta-analysis. Lancet 2019, 394, 939–951. (In English) [Google Scholar] [CrossRef]
- Chong, J.W.X.; Tan, E.H.-J.; Chong, C.E.; Ng, Y.; Wijesinghe, R. Atypical antipsychotics: A review on the prevalence, monitoring, and management of their metabolic and cardiovascular side effects. Ment. Health Clin. 2016, 6, 178–184. (In English) [Google Scholar] [CrossRef] [PubMed]
- Zhou, Y.; Danbolt, N.C. Glutamate as a neurotransmitter in the healthy brain. J. Neural Transm. 2014, 121, 799–817. (In English) [Google Scholar] [CrossRef] [PubMed]
- Farber, N.B. The NMDA receptor hypofunction model of psychosis. Ann. N. Y. Acad. Sci. 2003, 1003, 119–130. (In English) [Google Scholar] [CrossRef] [PubMed]
- Cadinu, D.; Grayson, B.; Podda, G.; Harte, M.K.; Doostdar, N.; Neill, J.C. NMDA receptor antagonist rodent models for cognition in schizophrenia and identification of novel drug treatments, an update. Neuropharmacology 2018, 142, 41–62. (In English) [Google Scholar] [CrossRef]
- Adell, A. Brain NMDA Receptors in Schizophrenia and Depression. Biomolecules 2020, 10, 947. (In English) [Google Scholar] [CrossRef]
- Goff, D. The Therapeutic Role of d-Cycloserine in Schizophrenia. Adv. Pharmacol. 2016, 76, 39–66. (In English) [Google Scholar] [CrossRef]
- MacKay, M.B.; Kravtsenyuk, M.; Thomas, R.; Mitchell, N.D.; Dursun, S.M.; Baker, G.B. D-Serine: Potential Therapeutic Agent and/or Biomarker in Schizophrenia and Depression? Front. Psychiatry 2019, 10, 25. (In English) [Google Scholar] [CrossRef] [PubMed]
- Ebrahimi-Ghiri, M.; Khakpai, F.; Zarrindast, M.R. Combined treatment of scopolamine and group III mGluR antagonist, CPPG, exerts antidepressant activity without affecting anxiety-related behaviors. Physiol. Behav. 2020, 224, 113034. (In English) [Google Scholar] [CrossRef] [PubMed]
- Chaki, S. mGlu2/3 receptor antagonists. Adv. Pharmacol. 2019, 86, 97–120. [Google Scholar] [CrossRef]
- Kim, J.H.; Marton, J.; Ametamey, S.M.; Cumming, P. A Review of Molecular Imaging of Glutamate Receptors. Molecules 2020, 25, 4749. (In English) [Google Scholar] [CrossRef]
- Niswender, C.M.; Conn, P.J. Metabotropic glutamate receptors: Physiology, pharmacology, and disease. Annu. Rev. Pharmacol. Toxicol. 2010, 50, 295–322. (In English) [Google Scholar] [CrossRef] [PubMed]
- Mazzitelli, M.; Palazzo, E.; Maione, S.; Neugebauer, V. Group II Metabotropic Glutamate Receptors: Role in Pain Mechanisms and Pain Modulation. Front. Mol. Neurosci. 2018, 11, 383. [Google Scholar] [CrossRef] [PubMed]
- Iacovelli, L.; Bruno, V.; Salvatore, L.; Melchiorri, D.; Gradini, R.; Caricasole, A.; Barletta, E.; De Blasi, A.; Nicoletti, F. Native group-III metabotropic glutamate receptors are coupled to the mitogen-activated protein kinase/phosphatidylinositol-3-kinase pathways. J. Neurochem. 2002, 82, 216–223. (In English) [Google Scholar] [CrossRef] [PubMed]
- Nawy, S. The metabotropic receptor mGluR6 may signal through G(o), but not phosphodiesterase, in retinal bipolar cells. J. Neurosci. 1999, 19, 2938–2944. (In English) [Google Scholar] [CrossRef]
- Chu, Z.; Hablitz, J.J. Quisqualate induces an inward current via mGluR activation in neocortical pyramidal neurons. Brain Res. 2000, 879, 88–92. (In English) [Google Scholar] [CrossRef]
- Hermans, E.; Challiss, R.A. Structural, signalling and regulatory properties of the group I metabotropic glutamate receptors: Prototypic family C G-protein-coupled receptors. Biochem. J. 2001, 359, 465–484. (In English) [Google Scholar] [CrossRef]
- Eng, A.G.; Kelver, D.A.; Hedrick, T.P.; Swanson, G.T. Transduction of group I mGluR-mediated synaptic plasticity by β-arrestin2 signalling. Nat. Commun. 2016, 7, 13571. (In English) [Google Scholar] [CrossRef]
- Reiner, A.; Levitz, J. Glutamatergic Signaling in the Central Nervous System: Ionotropic and Metabotropic Receptors in Concert. Neuron 2018, 98, 1080–1098. (In English) [Google Scholar] [CrossRef] [PubMed]
- Valenti, O.; Conn, P.J.; Marino, M.J. Distinct physiological roles of the Gq-coupled metabotropic glutamate receptors Co-expressed in the same neuronal populations. J. Cell. Physiol. 2002, 191, 125–137. (In English) [Google Scholar] [CrossRef] [PubMed]
- Liu, F.; Ma, X.-H.; Ule, J.; Bibb, J.A.; Nishi, A.; DeMaggio, A.J.; Yan, Z.; Nairn, A.C.; Greengard, P. Regulation of cyclin-dependent kinase 5 and casein kinase 1 by metabotropic glutamate receptors. Proc. Natl. Acad. Sci. USA 2001, 98, 11062–11068. (In English) [Google Scholar] [CrossRef] [PubMed]
- Stevenson, R.A.; Hoffman, J.L.; Maldonado-Devincci, A.M.; Faccidomo, S.; Hodge, C.W. MGluR5 activity is required for the induction of ethanol behavioral sensitization and associated changes in ERK MAP kinase phosphorylation in the nucleus accumbens shell and lateral habenula. Behav. Brain Res. 2019, 367, 19–27. (In English) [Google Scholar] [CrossRef] [PubMed]
- Schmit, T.L.; Dowell, J.A.; Maes, M.E.; Wilhelm, M. c-Jun N-terminal kinase regulates mGluR-dependent expression of post-synaptic FMRP target proteins. J. Neurochem. 2013, 127, 772–781. (In English) [Google Scholar] [CrossRef] [PubMed]
- Liu, F.; Virshup, D.M.; Nairn, A.C.; Greengard, P. Mechanism of regulation of casein kinase I activity by group I metabotropic glutamate receptors. J. Biol. Chem. 2002, 277, 45393–45399. (In English) [Google Scholar] [CrossRef]
- Pellegrini-Giampietro, D.E.; Torregrossa, S.A.; Moroni, F. Pharmacological characterization of metabotropic glutamate receptors coupled to phospholipase D in the rat hippocampus. Br. J. Pharmacol. 1996, 118, 1035–1043. (In English) [Google Scholar] [CrossRef] [PubMed]
- Abd-Elrahman, K.S.; Ferguson, S.S.G. Modulation of mTOR and CREB pathways following mGluR5 blockade contribute to improved Huntington’s pathology in zQ175 mice. Mol. Brain 2019, 12, 35. (In English) [Google Scholar] [CrossRef]
- Maksymetz, J.; Moran, S.P.; Conn, P.J. Targeting metabotropic glutamate receptors for novel treatments of schizophrenia. Mol. Brain 2017, 10, 15. (In English) [Google Scholar] [CrossRef] [PubMed]
- Correa, A.M.B.; Guimarães, J.D.S.; Alhadas, E.D.S.E.; Kushmerick, C. Control of neuronal excitability by Group I metabotropic glutamate receptors. Biophys. Rev. 2017, 9, 835–845. (In English) [Google Scholar] [CrossRef] [PubMed]
- Gerber, U.; Gee, C.E.; Benquet, P. Metabotropic glutamate receptors: Intracellular signaling pathways. Curr. Opin. Pharmacol. 2007, 7, 56–61. (In English) [Google Scholar] [CrossRef] [PubMed]
- Volk, D.W.; Eggan, S.M.; Lewis, D.A. Alterations in metabotropic glutamate receptor 1α and regulator of G protein signaling 4 in the prefrontal cortex in schizophrenia. Am. J. Psychiatry 2010, 167, 1489–1498. (In English) [Google Scholar] [CrossRef] [PubMed]
- Brody, S.A.; Conquet, F.; Geyer, M.A. Disruption of prepulse inhibition in mice lacking mGluR1. Eur. J. Neurosci. 2003, 18, 3361–3366. (In English) [Google Scholar] [CrossRef] [PubMed]
- Hikichi, H.; Nishino, M.; Fukushima, M.; Satow, A.; Maehara, S.; Kawamoto, H.; Ohta, H. Pharmacological effects of metabotropic glutamate receptor ligands on prepulse inhibition in DBA/2J mice. Eur. J. Pharmacol. 2010, 639, 99–105. (In English) [Google Scholar] [CrossRef]
- Braff, D.L.; Grillon, C.; Geyer, M.A. Gating and habituation of the startle reflex in schizophrenic patients. Arch. Gen. Psychiatry 1992, 49, 206–215. (In English) [Google Scholar] [CrossRef] [PubMed]
- Frank, R.A.W.; McRae, A.F.; Pocklington, A.J.; Van De Lagemaat, L.N.; Navarro, P.; Croning, M.D.R.; Komiyama, N.H.; Bradley, S.J.; Challiss, R.A.J.; Armstrong, J.D.; et al. Clustered coding variants in the glutamate receptor complexes of individuals with schizophrenia and bipolar disorder. PLoS ONE 2011, 6, e19011. (In English) [Google Scholar] [CrossRef] [PubMed]
- Ayoub, M.A.; Angelicheva, D.; Vile, D.; Chandler, D.; Morar, B.; Cavanaugh, J.A.; Visscher, P.M.; Jablensky, A.; Pfleger, K.D.G.; Kalaydjieva, L. Deleterious GRM1 mutations in schizophrenia. PLoS ONE 2012, 7, e32849. (In English) [Google Scholar] [CrossRef] [PubMed]
- Cho, H.P.; Garcia-Barrantes, P.M.; Brogan, J.T.; Hopkins, C.R.; Niswender, C.M.; Rodriguez, A.L.; Venable, D.F.; Morrison, R.D.; Bubser, M.; Daniels, J.S.; et al. Chemical modulation of mutant mGlu1 receptors derived from deleterious GRM1 mutations found in schizophrenics. ACS Chem. Biol. 2014, 9, 2334–2346. (In English) [Google Scholar] [CrossRef]
- Lesage, A.; Steckler, T. Metabotropic glutamate mGlu1 receptor stimulation and blockade: Therapeutic opportunities in psychiatric illness. Eur. J. Pharmacol. 2010, 639, 2–16. (In English) [Google Scholar] [CrossRef]
- Zoicas, I.; Kornhuber, J. The Role of Metabotropic Glutamate Receptors in Social Behavior in Rodents. Int. J. Mol. Sci. 2019, 20, 1412. (In English) [Google Scholar] [CrossRef]
- Foster, D.J.; Conn, P.J. Allosteric Modulation of GPCRs: New Insights and Potential Utility for Treatment of Schizophrenia and Other CNS Disorders. Neuron 2017, 94, 431–446. (In English) [Google Scholar] [CrossRef]
- Satow, A.; Maehara, S.; Ise, S.; Hikichi, H.; Fukushima, M.; Suzuki, G.; Kimura, T.; Tanaka, T.; Ito, S.; Kawamoto, H.; et al. Pharmacological effects of the metabotropic glutamate receptor 1 antagonist compared with those of the metabotropic glutamate receptor 5 antagonist and metabotropic glutamate receptor 2/3 agonist in rodents: Detailed investigations with a selective allosteric metabotropic glutamate receptor 1 antagonist, FTIDC [4-[1-(2-fluoropyridine-3-yl)-5-methyl-1H-1,2,3-triazol-4-yl]-N-isopropyl-N-methyl-3,6-dihydropyridine-1(2H)-carboxamide]. J. Pharmacol. Exp. Ther. 2008, 326, 577–586. (In English) [Google Scholar] [CrossRef]
- Conn, P.J.; Lindsley, C.W.; Jones, C.K. Activation of metabotropic glutamate receptors as a novel approach for the treatment of schizophrenia. Trends Pharmacol. Sci. 2009, 30, 25–31. (In English) [Google Scholar] [CrossRef] [PubMed]
- Satow, A.; Suzuki, G.; Maehara, S.; Hikichi, H.; Murai, T.; Murai, T.; Kawagoe-Takaki, H.; Hata, M.; Ito, S.; Ozaki, S.; et al. Unique antipsychotic activities of the selective metabotropic glutamate receptor 1 allosteric antagonist 2-cyclopropyl-5-[1-(2-fluoro-3-pyridinyl)-5-methyl-1H-1,2,3-triazol-4-yl]-2,3-dihydro-1H-isoindol-1-one. J. Pharmacol. Exp. Ther. 2009, 330, 179–190. (In English) [Google Scholar] [CrossRef] [PubMed]
- Kinney, G.G.; Burno, M.; Campbell, U.C.; Hernandez, L.M.; Rodriguez, D.; Bristow, L.J.; Conn, P.J. Metabotropic glutamate subtype 5 receptors modulate locomotor activity and sensorimotor gating in rodents. J. Pharmacol. Exp. Ther. 2003, 306, 116–123. (In English) [Google Scholar] [CrossRef]
- Vales, K.; Svoboda, J.; Benkovicova, K.; Bubenikova-Valesova, V.; Stuchlik, A. The difference in effect of mGlu2/3 and mGlu5 receptor agonists on cognitive impairment induced by MK-801. Eur. J. Pharmacol. 2010, 639, 91–98. (In English) [Google Scholar] [CrossRef] [PubMed]
- Chan, M.H.; Chiu, P.H.; Sou, J.H.; Chen, H.H. Attenuation of ketamine-evoked behavioral responses by mGluR5 positive modulators in mice. Psychopharmacology 2008, 198, 141–148. (In English) [Google Scholar] [CrossRef] [PubMed]
- Kinney, G.G.; O’Brien, J.A.; Lemaire, W.; Burno, M.; Bickel, D.J.; Clements, M.K.; Chen, T.-B.; Wisnoski, D.D.; Lindsley, C.W.; Tiller, P.R.; et al. A novel selective positive allosteric modulator of metabotropic glutamate receptor subtype 5 has in vivo activity and antipsychotic-like effects in rat behavioral models. J. Pharmacol. Exp. Ther. 2004, 313, 199–206. (In English) [Google Scholar] [CrossRef]
- Vardigan, J.D.; Huszar, S.L.; McNaughton, C.H.; Hutson, P.H.; Uslaner, J.M. MK-801 produces a deficit in sucrose preference that is reversed by clozapine, D-serine, and the metabotropic glutamate 5 receptor positive allosteric modulator CDPPB: Relevance to negative symptoms associated with schizophrenia? Pharmacol. Biochem. Behav. 2010, 95, 223–229. (In English) [Google Scholar] [CrossRef]
- Stefani, M.R.; Moghaddam, B. Activation of type 5 metabotropic glutamate receptors attenuates deficits in cognitive flexibility induced by NMDA receptor blockade. Eur. J. Pharmacol. 2010, 639, 26–32. (In English) [Google Scholar] [CrossRef] [PubMed]
- Liu, F.; Grauer, S.; Kelley, C.; Navarra, R.L.; Graf, R.; Zhang, G.; Atkinson, P.J.; Popiolek, M.; Wantuch, C.M.; Khawaja, X.; et al. ADX47273 [S-(4-fluoro-phenyl)-{3-[3-(4-fluoro-phenyl)-(1,2,4)-oxadiazol-5-yl]-piperidin-1-yl}-methanone]: A novel metabotropic glutamate receptor 5-selective positive allosteric modulator with preclinical antipsychotic-like and procognitive activities. J. Pharmacol. Exp. Ther. 2008, 327, 827–839. (In English) [Google Scholar] [CrossRef] [PubMed]
- Rook, J.M.; Xiang, Z.; Lv, X.; Ghoshal, A.; Dickerson, J.W.; Bridges, T.M.; Johnson, K.A.; Foster, D.J.; Gregory, K.J.; Vinson, P.N.; et al. Biased mGlu5-Positive Allosteric Modulators Provide In Vivo Efficacy without Potentiating mGlu5 Modulation of NMDAR Currents. Neuron 2015, 86, 1029–1040. [Google Scholar] [CrossRef] [PubMed]
- Parmentier-Batteur, S.; Hutson, P.H.; Menzel, K.; Uslaner, J.M.; Mattson, B.A.; O’Brien, J.A.; Magliaro, B.C.; Forest, T.; Stump, C.A.; Tynebor, R.M.; et al. Mechanism based neurotoxicity of mGlu5 positive allosteric modulators—Development challenges for a promising novel antipsychotic target. Neuropharmacology 2014, 82, 161–173. (In English) [Google Scholar] [CrossRef]
- Balu, D.T.; Li, Y.; Takagi, S.; Presti, K.T.; Ramikie, T.S.; Rook, J.M.; Jones, C.K.; Lindsley, C.W.; Conn, P.J.; Bolshakov, V.Y.; et al. An mGlu5-Positive Allosteric Modulator Rescues the Neuroplasticity Deficits in a Genetic Model of NMDA Receptor Hypofunction in Schizophrenia. Neuropsychopharmacology 2016, 41, 2052–2061. (In English) [Google Scholar] [CrossRef]
- Crupi, R.; Impellizzeri, D.; Cuzzocrea, S. Role of Metabotropic Glutamate Receptors in Neurological Disorders. Front. Mol. Neurosci. 2019, 12, 20. (In English) [Google Scholar] [CrossRef]
- Ohishi, H.; Shigemoto, R.; Nakanishi, S.; Mizuno, N. Distribution of the mRNA for a metabotropic glutamate receptor (mGluR3) in the rat brain: An in situ hybridization study. J. Comp. Neurol. 1993, 335, 252–266. (In English) [Google Scholar] [CrossRef]
- Ferraguti, F.; Shigemoto, R. Metabotropic glutamate receptors. Cell Tissue Res. 2006, 326, 483–504. (In English) [Google Scholar] [CrossRef] [PubMed]
- Yin, S.; Noetzel, M.J.; Johnson, K.A.; Zamorano, R.; Jalan-Sakrikar, N.; Gregory, K.J.; Conn, P.J.; Niswender, C.M. Selective actions of novel allosteric modulators reveal functional heteromers of metabotropic glutamate receptors in the CNS. J. Neurosci. 2014, 34, 79–94. (In English) [Google Scholar] [CrossRef] [PubMed]
- Di Menna, L.; Joffe, M.E.; Iacovelli, L.; Orlando, R.; Lindsley, C.W.; Mairesse, J.; Gressèns, P.; Cannella, M.; Caraci, F.; Copani, A.; et al. Functional partnership between mGlu3 and mGlu5 metabotropic glutamate receptors in the central nervous system. Neuropharmacology 2018, 128, 301–313. (In English) [Google Scholar] [CrossRef] [PubMed]
- Pehrson, A.L.; Moghaddam, B. Impact of metabotropic glutamate 2/3 receptor stimulation on activated dopamine release and locomotion. Psychopharmacology 2010, 211, 443–455. (In English) [Google Scholar] [CrossRef] [PubMed]
- Moghaddam, B.; Adams, B.W. Reversal of phencyclidine effects by a group II metabotropic glutamate receptor agonist in rats. Science 1998, 281, 1349–1352. (In English) [Google Scholar] [CrossRef] [PubMed]
- Uslaner, J.M.; Smith, S.M.; Huszar, S.L.; Pachmerhiwala, R.; Hinchliffe, R.M.; Vardigan, J.D.; Hutson, P.H. Combined administration of an mGlu2/3 receptor agonist and a 5-HT 2A receptor antagonist markedly attenuate the psychomotor-activating and neurochemical effects of psychostimulants. Psychopharmacology 2009, 206, 641–651. (In English) [Google Scholar] [CrossRef] [PubMed]
- Krystal, J.H.; Abi-Saab, W.; Perry, E.; D’Souza, D.C.; Liu, N.; Gueorguieva, R.; McDougall, L.; Hunsberger, T.; Belger, A.; Levine, L.; et al. Preliminary evidence of attenuation of the disruptive effects of the NMDA glutamate receptor antagonist, ketamine, on working memory by pretreatment with the group II metabotropic glutamate receptor agonist, LY354740, in healthy human subjects. Psychopharmacology 2004, 179, 303–309. (In English) [Google Scholar] [CrossRef] [PubMed]
- Lorrain, D.S.; Baccei, C.S.; Bristow, L.J.; Anderson, J.J.; Varney, M.A. Effects of ketamine and N-methyl-D-aspartate on glutamate and dopamine release in the rat prefrontal cortex: Modulation by a group II selective metabotropic glutamate receptor agonist LY379268. Neuroscience 2003, 117, 697–706. (In English) [Google Scholar] [CrossRef]
- Spampinato, S.F.; Copani, A.; Nicoletti, F.; Sortino, M.A.; Caraci, F. Metabotropic Glutamate Receptors in Glial Cells: A New Potential Target for Neuroprotection? Front. Mol. Neurosci. 2018, 11, 414. (In English) [Google Scholar] [CrossRef] [PubMed]
- Ster, J.; Mateos, J.M.; Grewe, B.F.; Coiret, G.; Corti, C.; Corsi, M.; Helmchen, F.; Gerber, U. Enhancement of CA3 hippocampal network activity by activation of group II metabotropic glutamate receptors. Proc. Natl. Acad. Sci. USA 2011, 108, 9993–9997. (In English) [Google Scholar] [CrossRef]
- González-Maeso, J.; Ang, R.L.; Yuen, T.; Chan, P.; Weisstaub, N.V.; López-Giménez, J.F.; Zhou, M.; Okawa, Y.; Callado, L.F.; Milligan, G.; et al. Identification of a serotonin/glutamate receptor complex implicated in psychosis. Nature 2008, 452, 93–97. (In English) [Google Scholar] [CrossRef]
- Moreno, J.L.; Holloway, T.; Albizu, L.; Sealfon, S.C.; González-Maeso, J. Metabotropic glutamate mGlu2 receptor is necessary for the pharmacological and behavioral effects induced by hallucinogenic 5-HT2A receptor agonists. Neurosci. Lett. 2011, 493, 76–79. (In English) [Google Scholar] [CrossRef]
- Kurita, M.; Holloway, T.; García-Bea, A.; Kozlenkov, A.; Friedman, A.K.; Moreno, J.L.; Heshmati, M.; Golden, S.A.; Kennedy, P.J.; Takahashi, N.; et al. HDAC2 regulates atypical antipsychotic responses through the modulation of mGlu2 promoter activity. Nat. Neurosci. 2012, 15, 1245–1254. (In English) [Google Scholar] [CrossRef] [PubMed]
- Woolley, M.L.; Pemberton, D.J.; Bate, S.; Corti, C.; Jones, D.N. The mGlu2 but not the mGlu3 receptor mediates the actions of the mGluR2/3 agonist, LY379268, in mouse models predictive of antipsychotic activity. Psychopharmacology 2008, 196, 431–440. (In English) [Google Scholar] [CrossRef] [PubMed]
- Cartmell, J.; Monn, J.A.; Schoepp, D.D. The metabotropic glutamate 2/3 receptor agonists LY354740 and LY379268 selectively attenuate phencyclidine versus d-amphetamine motor behaviors in rats. J. Pharmacol. Exp. Ther. 1999, 291, 161–170. (In English) [Google Scholar]
- Cartmell, J.; Monn, J.A.; Schoepp, D.D. The mGlu(2/3) receptor agonist LY379268 selectively blocks amphetamine ambulations and rearing. Eur. J. Pharmacol. 2000, 400, 221–224. [Google Scholar] [CrossRef]
- Cartmell, J.; Monn, J.A.; Schoepp, D.D. Attenuation of specific PCP-evoked behaviors by the potent mGlu2/3 receptor agonist, LY379268 and comparison with the atypical antipsychotic, clozapine. Psychopharmacology 2000, 148, 423–429. (In English) [Google Scholar] [CrossRef]
- Galici, R.; Echemendia, N.G.; Rodriguez, A.L.; Conn, P.J. A selective allosteric potentiator of metabotropic glutamate (mGlu) 2 receptors has effects similar to an orthosteric mGlu2/3 receptor agonist in mouse models predictive of antipsychotic activity. J. Pharmacol. Exp. Ther. 2005, 315, 1181–1187. (In English) [Google Scholar] [CrossRef]
- Schlumberger, C.; Schäfer, D.; Barberi, C.; Morè, L.; Nagel, J.; Pietraszek, M.; Schmidt, W.J.; Danysz, W. Effects of a metabotropic glutamate receptor group II agonist LY354740 in animal models of positive schizophrenia symptoms and cognition. Behav. Pharmacol. 2009, 20, 56–66. (In English) [Google Scholar] [CrossRef]
- Profaci, C.P.; Krolikowski, K.A.; Olszewski, R.T.; Neale, J.H. Group II mGluR agonist LY354740 and NAAG peptidase inhibitor effects on prepulse inhibition in PCP and D-amphetamine models of schizophrenia. Psychopharmacology 2011, 216, 235–243. (In English) [Google Scholar] [CrossRef] [PubMed]
- Harich, S.; Gross, G.; Bespalov, A. Stimulation of the metabotropic glutamate 2/3 receptor attenuates social novelty discrimination deficits induced by neonatal phencyclidine treatment. Psychopharmacology 2007, 192, 511–519. (In English) [Google Scholar] [CrossRef] [PubMed]
- Chartoff, E.H.; Heusner, C.L.; Palmiter, R.D. Dopamine is not required for the hyperlocomotor response to NMDA receptor antagonists. Neuropsychopharmacology 2005, 30, 1324–1333. (In English) [Google Scholar] [CrossRef]
- Rorick-Kehn, L.M.; Perkins, E.J.; Knitowski, K.M.; Hart, J.C.; Johnson, B.G.; Schoepp, D.D.; McKinzie, D.L. Improved bioavailability of the mGlu2/3 receptor agonist LY354740 using a prodrug strategy: In vivo pharmacology of LY544344. J. Pharmacol. Exp. Ther. 2005, 316, 905–913. (In English) [Google Scholar] [CrossRef] [PubMed]
- Imre, G.; Salomons, A.; Jongsma, M.; Fokkema, D.S.; den Boer, J.A.; Horst, G.J.T. Effects of the mGluR2/3 agonist LY379268 on ketamine-evoked behaviours and neurochemical changes in the dentate gyrus of the rat. Pharmacol. Biochem. Behav. 2006, 84, 392–399. (In English) [Google Scholar] [CrossRef] [PubMed]
- Xi, D.; Li, Y.-C.; Snyder, M.A.; Gao, R.Y.; Adelman, A.E.; Zhang, W.; Shumsky, J.S.; Gao, W.-J. Group II metabotropic glutamate receptor agonist ameliorates MK801-induced dysfunction of NMDA receptors via the Akt/GSK-3β pathway in adult rat prefrontal cortex. Neuropsychopharmacology 2011, 36, 1260–1274. (In English) [Google Scholar] [CrossRef] [PubMed]
- Winter, J.C.; Eckler, J.R.; Rabin, R.A. Serotonergic/glutamatergic interactions: The effects of mGlu2/3 receptor ligands in rats trained with LSD and PCP as discriminative stimuli. Psychopharmacology 2004, 172, 233–240. (In English) [Google Scholar] [CrossRef]
- Lorrain, D.S.; Schaffhauser, H.; Campbell, U.C.; Baccei, C.S.; Correa, L.D.; Rowe, B.; Rodriguez, D.E.; Anderson, J.J.; Varney, M.A.; Pinkerton, A.B.; et al. Group II mGlu receptor activation suppresses norepinephrine release in the ventral hippocampus and locomotor responses to acute ketamine challenge. Neuropsychopharmacology 2003, 28, 1622–1632. (In English) [Google Scholar] [CrossRef]
- Swanson, C.J.; Schoepp, D.D. The group II metabotropic glutamate receptor agonist (-)-2-oxa-4-aminobicyclo (3.1.0.)hexane-4,6-dicarboxylate (LY379268) and clozapine reverse phencyclidine-induced behaviors in monoamine-depleted rats. J. Pharmacol. Exp. Ther. 2002, 303, 919–927. (In English) [Google Scholar] [CrossRef] [PubMed]
- Engel, M.; Snikeris, P.; Matosin, N.; Newell, K.A.; Huang, X.F.; Frank, E. mGluR2/3 agonist LY379268 rescues NMDA and GABAA receptor level deficits induced in a two-hit mouse model of schizophrenia. Psychopharmacology 2016, 233, 1349–1359. (In English) [Google Scholar] [CrossRef] [PubMed]
- Sokolenko, E.; Hudson, M.R.; Nithianantharajah, J.; Jones, N.C. The mGluR. J. Psychopharmacol. 2019, 33, 1588–1599. (In English) [Google Scholar] [CrossRef] [PubMed]
- Jones, C.A.; Brown, A.M.; Auer, D.P.; Fone, K.C. The mGluR2/3 agonist LY379268 reverses post-weaning social isolation-induced recognition memory deficits in the rat. Psychopharmacology 2011, 214, 269–283. (In English) [Google Scholar] [CrossRef] [PubMed]
- Matrisciano, F.; Dong, E.; Nicoletti, F.; Guidotti, A. Epigenetic Alterations in Prenatal Stress Mice as an Endophenotype Model for Schizophrenia: Role of Metabotropic Glutamate 2/3 Receptors. Front. Mol. Neurosci. 2018, 11, 423. (In English) [Google Scholar] [CrossRef] [PubMed]
- Holloway, T.; Moreno, J.L.; Umali, A.; Rayannavar, V.; Hodes, G.E.; Russo, S.J.; González-Maeso, J. Prenatal stress induces schizophrenia-like alterations of serotonin 2A and metabotropic glutamate 2 receptors in the adult offspring: Role of maternal immune system. J. Neurosci. 2013, 33, 1088–1098. (In English) [Google Scholar] [CrossRef] [PubMed]
- Xing, B.; Han, G.; Wang, M.J.; Snyder, M.A.; Gao, W.J. Juvenile treatment with mGluR2/3 agonist prevents schizophrenia-like phenotypes in adult by acting through GSK3β. Neuropharmacology 2018, 137, 359–371. (In English) [Google Scholar] [CrossRef]
- Li, M.-L.; Gulchina, Y.; Monaco, S.A.; Xing, B.; Ferguson, B.R.; Li, Y.-C.; Li, F.; Hu, X.-Q.; Gao, W.-J. Juvenile treatment with a novel mGluR2 agonist/mGluR3 antagonist compound, LY395756, reverses learning deficits and cognitive flexibility impairments in adults in a neurodevelopmental model of schizophrenia. Neurobiol. Learn. Mem. 2017, 140, 52–61. (In English) [Google Scholar] [CrossRef] [PubMed]
- Riegel, A.C.; Ali, S.F.; French, E.D. Toluene-induced locomotor activity is blocked by 6-hydroxydopamine lesions of the nucleus accumbens and the mGluR2/3 agonist LY379268. Neuropsychopharmacology 2003, 28, 1440–1447. (In English) [Google Scholar] [CrossRef] [PubMed]
- Cartmell, J.; Monn, J.A.; Schoepp, D.D. Tolerance to the motor impairment, but not to the reversal of PCP-induced motor activities by oral administration of the mGlu2/3 receptor agonist, LY379268. Naunyn Schmiedeberg Arch. Pharmacol. 2000, 361, 39–46. (In English) [Google Scholar] [CrossRef] [PubMed]
- Schoepp, D.; Marek, G. Preclinical pharmacology of mGlu2/3 receptor agonists: Novel agents for schizophrenia? Curr. Drug Target CNS Neurol. Disord. 2002, 1, 215–225. (In English) [Google Scholar] [CrossRef] [PubMed]
- Burgos-Aguilar, C.; Ferris, M.J.; Sexton, L.L.; Sun, H.; Xiao, R.; Chen, R.; Childers, S.R.; Howlett, A.C. Metabotropic glutamate 2,3 receptor stimulation desensitizes agonist activation of G-protein signaling and alters transcription regulators in mesocorticolimbic brain regions. Synapse 2021, 75, e22190. (In English) [Google Scholar] [CrossRef] [PubMed]
- Nikiforuk, A.; Popik, P.; Drescher, K.U.; Van Gaalen, M.; Relo, A.-L.; Mezler, M.; Marek, G.; Schoemaker, H.; Gross, G.; Bespalov, A. Effects of a positive allosteric modulator of group II metabotropic glutamate receptors, LY487379, on cognitive flexibility and impulsive-like responding in rats. J. Pharmacol. Exp. Ther. 2010, 335, 665–673. (In English) [Google Scholar] [CrossRef] [PubMed]
- Hikichi, H.; Kaku, A.; Karasawa, J.; Chaki, S. Stimulation of metabotropic glutamate (mGlu) 2 receptor and blockade of mGlu1 receptor improve social memory impairment elicited by MK-801 in rats. J. Pharmacol. Sci. 2013, 122, 10–16. (In English) [Google Scholar] [CrossRef]
- Benneyworth, M.A.; Xiang, Z.; Smith, R.L.; Garcia, E.E.; Conn, P.J.; Sanders-Bush, E. A selective positive allosteric modulator of metabotropic glutamate receptor subtype 2 blocks a hallucinogenic drug model of psychosis. Mol. Pharmacol. 2007, 72, 477–484. (In English) [Google Scholar] [CrossRef]
- Lavreysen, H.; Ahnaou, A.; Drinkenburg, W.; Langlois, X.; Mackie, C.; Pype, S.; Lutjens, R.; Le Poul, E.; Trabanco, A.A.; Nuñez, J.M.C. Pharmacological and pharmacokinetic properties of JNJ-40411813, a positive allosteric modulator of the mGlu2 receptor. Pharmacol. Res. Perspect. 2015, 3, e00096. (In English) [Google Scholar] [CrossRef] [PubMed]
- Griebel, G.; Pichat, P.; Boulay, D.; Naimoli, V.; Potestio, L.; Featherstone, R.; Sahni, S.; DeFex, H.; Desvignes, C.; Slowinski, F.; et al. The mGluR2 positive allosteric modulator, SAR218645, improves memory and attention deficits in translational models of cognitive symptoms associated with schizophrenia. Sci. Rep. 2016, 6, 35320. (In English) [Google Scholar] [CrossRef] [PubMed]
- Patil, S.T.; Zhang, L.; Martenyi, F.; Lowe, S.L.; Jackson, K.A.; Andreev, B.V.; Avedisova, A.S.; Bardenstein, L.M.; Gurovich, I.Y.; Morozova, M.A.; et al. Activation of mGlu2/3 receptors as a new approach to treat schizophrenia: A randomized Phase 2 clinical trial. Nat. Med. 2007, 13, 1102–1107. (In English) [Google Scholar] [CrossRef] [PubMed]
- Kinon, B.J.; Gómez, J.C. Clinical development of pomaglumetad methionil: A non-dopaminergic treatment for schizophrenia. Neuropharmacology 2013, 66, 82–86. (In English) [Google Scholar] [CrossRef] [PubMed]
- Adams, D.H.; Kinon, B.J.; Baygani, S.; Millen, B.A.; Velona, I.; Kollack-Walker, S.; Walling, D.P. A long-term, phase 2, multicenter, randomized, open-label, comparative safety study of pomaglumetad methionil (LY2140023 monohydrate) versus atypical antipsychotic standard of care in pa tients with schizophrenia. BMC Psychiatry 2013, 13, 143. (In English) [Google Scholar] [CrossRef] [PubMed]
- Li, M.L.; Hu, X.Q.; Li, F.; Gao, W.J. Perspectives on the mGluR2/3 agonists as a therapeutic target for schizophrenia: Still promising or a dead end? Prog. Neuro Psychopharmacol. Biol. Psychiatry 2015, 60, 66–76. (In English) [Google Scholar] [CrossRef]
- Hopkins, C.R. Is there a path forward for mGlu(2) positive allosteric modulators for the treatment of schizophrenia? ACS Chem. Neurosci. 2012, 4, 211–213. (In English) [Google Scholar] [CrossRef] [PubMed][Green Version]
- Salih, H.; Anghelescu, I.; Kezic, I.; Sinha, V.; Hoeben, E.; Van Nueten, L.; De Smedt, H.; De Boer, P. Pharmacokinetic and pharmacodynamic characterisation of JNJ-40411813, a positive allosteric modulator of mGluR2, in two randomised, double-blind phase-I studies. J. Psychopharmacol. 2014, 29, 414–425. (In English) [Google Scholar] [CrossRef]
- Litman, R.E.; Smith, M.A.; Doherty, J.J.; Cross, A.; Raines, S.; Gertsik, L.; Zukin, S.R. AZD8529, a positive allosteric modulator at the mGluR2 receptor, does not improve symptoms in schizophrenia: A proof of principle study. Schizophr. Res. 2016, 172, 152–157. (In English) [Google Scholar] [CrossRef]
- Javitt, D.C.; Carter, C.S.; Krystal, J.H.; Kantrowitz, J.T.; Girgis, R.R.; Kegeles, L.S.; Ragland, J.D.; Maddock, R.J.; Lesh, T.A.; Tanase, C.; et al. Utility of Imaging-Based Biomarkers for Glutamate-Targeted Drug Development in Psychotic Disorders: A Randomized Clinical Trial. JAMA Psychiatry 2018, 75, 11–19. (In English) [Google Scholar] [CrossRef] [PubMed]
- Kantrowitz, J.T.; Grinband, J.; Goff, D.C.; Lahti, A.C.; Marder, S.R.; Kegeles, L.S.; Girgis, R.R.; Sobeih, T.; Wall, M.M.; Choo, T.-H.; et al. Proof of mechanism and target engagement of glutamatergic drugs for the treatment of schizophrenia: RCTs of pomaglumetad and TS-134 on ketamine-induced psychotic symptoms and pharmacoBOLD in healthy volunteers. Neuropsychopharmacology 2020, 45, 1842–1850. (In English) [Google Scholar] [CrossRef] [PubMed]
- Tandon, R.; Keshavan, M.S.; Nasrallah, H.A. Schizophrenia, “Just the Facts”: What we know in 2008 part 1: Overview. Schizophr. Res. 2008, 100, 4–19. (In English) [Google Scholar] [CrossRef] [PubMed]
- Miyamoto, S.; Duncan, G.E.; Marx, C.E.; Lieberman, J.A. Treatments for schizophrenia: A critical review of pharmacology and mechanisms of action of antipsychotic drugs. Mol. Psychiatry 2004, 10, 79–104. (In English) [Google Scholar] [CrossRef]
- Ayan-Oshodi, M.; Wondmagegnehu, E.T.; Lowe, S.L.; Kryzhanovskaya, L.; Walker, D.J.; Kinon, B.J. Adverse events in healthy subjects exposed to single and multiple doses of LY2140023 monohydrate: Pooled results from 10 phase 1 studies. J. Clin. Psychopharmacol. 2012, 32, 408–411. (In English) [Google Scholar] [CrossRef]
- Stauffer, V.L.; Baygani, S.K.; Kinon, B.J.; Krikke-Workel, J.O. A short-term, multicenter, placebo-controlled, randomized withdrawal study of a metabotropic glutamate 2/3 receptor agonist using an electronic patient-reported outcome device in patients with schizophrenia. J Clin Psychopharmacol. J. Clin. Psychopharmacol. 2014, 34, 552–558. (In English) [Google Scholar] [CrossRef] [PubMed]
- Chue, P.; Lalonde, J.K. Addressing the unmet needs of patients with persistent negative symptoms of schizophrenia: Emerging pharmacological treatment options. Neuropsychiatr. Dis. Treat. 2014, 10, 777–789. (In English) [Google Scholar] [CrossRef][Green Version]
- Adams, D.H.; Zhang, L.; Millen, B.A.; Kinon, B.J.; Gomez, J.C. Pomaglumetad Methionil (LY2140023 Monohydrate) and Aripiprazole in Patients with Schizophrenia: A Phase 3, Multicenter, Double-Blind Comparison. Schizophr. Res. Treat. 2014, 2014, 758212. (In English) [Google Scholar] [CrossRef] [PubMed]
- Cartmell, J.; Schoepp, D.D. Regulation of neurotransmitter release by metabotropic glutamate receptors. J. Neurochem. 2002, 75, 889–907. (In English) [Google Scholar] [CrossRef]
- Lorez, M.; Humbel, U.; Pflimlin, M.C.; Kew, J.N. Group III metabotropic glutamate receptors as autoreceptors in the cerebellar cortex. Br. J. Pharmacol. 2003, 138, 614–625. (In English) [Google Scholar] [CrossRef]
- Pekhletski, R.; Gerlai, R.; Overstreet, L.S.; Huang, X.-P.; Agopyan, N.; Slater, N.T.; Abramow-Newerly, W.; Roder, J.C.; Hampson, D.R. Impaired cerebellar synaptic plasticity and motor performance in mice lacking the mGluR4 subtype of metabotropic glutamate receptor. J. Neurosci. 1996, 16, 6364–6373. (In English) [Google Scholar] [CrossRef]
- Nakajima, Y.; Iwakabe, H.; Akazawa, C.; Nawa, H.; Shigemoto, R.; Mizuno, N.; Nakanishi, S. Molecular characterization of a novel retinal metabotropic glutamate receptor mGluR6 with a high agonist selectivity for L-2-amino-4-phosphonobutyrate. J. Biol. Chem. 1993, 268, 11868–11873. (In English) [Google Scholar] [CrossRef]
- Corti, C.; Aldegheri, L.; Somogyi, P.; Ferraguti, F. Distribution and synaptic localisation of the metabotropic glutamate receptor 4 (mGluR4) in the rodent CNS. Neuroscience 2002, 110, 403–420. (In English) [Google Scholar] [CrossRef]
- Daniel, H.; Crepel, F. Control of Ca (2+) influx by cannabinoid and metabotropic glutamate receptors in rat cerebellar cortex requires K (+) channels. J. Physiol. 2001, 537, 793–800. (In English) [Google Scholar] [CrossRef]
- Abitbol, K.; Acher, F.; Daniel, H. Depression of excitatory transmission at PF-PC synapse by group III metabotropic glutamate receptors is provided exclusively by mGluR4 in the rodent cerebellar cortex. J. Neurochem. 2008, 105, 2069–2079. (In English) [Google Scholar] [CrossRef] [PubMed]
- Weber, M.; Schnitzler, H.U.; Schmid, S. Synaptic plasticity in the acoustic startle pathway: The neuronal basis for short-term habituation? Eur. J. Neurosci. 2002, 16, 1325–1332. (In English) [Google Scholar] [CrossRef] [PubMed]
- Goddyn, H.; Callaerts-Vegh, Z.; D’Hooge, R. Functional Dissociation of Group III Metabotropic Glutamate Receptors Revealed by Direct Comparison between the Behavioral Profiles of Knockout Mouse Lines. Int. J. Neuropsychopharmacol. 2015, 18, pyv053. (In English) [Google Scholar] [CrossRef] [PubMed]
- Bruno, V.; Battaglia, G.; Książek, I.; Van Der Putten, H.; Catania, M.V.; Giuffrida, R.; Lukic, S.; Leonhardt, T.; Inderbitzin, W.; Gasparini, F.; et al. Selective activation of mGlu4 metabotropic glutamate receptors is protective against excitotoxic neuronal death. J. Neurosci. 2000, 20, 6413–6420. (In English) [Google Scholar] [CrossRef] [PubMed]
- Senter, R.K.; Ghoshal, A.; Walker, A.G.; Xiang, Z.; Niswender, C.M.; Conn, P.J. The Role of mGlu Receptors in Hippocampal Plasticity Deficits in Neurological and Psychiatric Disorders: Implications for Allosteric Modulators as Novel Therapeutic Strategies. Curr. Neuropharmacol. 2016, 14, 455–473. (In English) [Google Scholar] [CrossRef]
- Mena, A.; Ruiz-Salas, J.C.; Puentes, A.; Dorado, I.; Ruiz-Veguilla, M.; de la Casa, L.G. Reduced Prepulse Inhibition as a Biomarker of Schizophrenia. Front. Behav. Neurosci. 2016, 10, 202. (In English) [Google Scholar] [CrossRef] [PubMed]
- Swerdlow, N.R.; Weber, M.; Qu, Y.; Light, G.A.; Braff, D.L. Realistic expectations of prepulse inhibition in translational models for schizophrenia research. Psychopharmacology 2008, 199, 331–388. (In English) [Google Scholar] [CrossRef]
- Beurrier, C.; Lopez, S.; Révy, D.; Selvam, C.; Goudet, C.; Lhérondel, M.; Gubellini, P.; Kerkerian-Legoff, L.; Acher, F.; Pin, J.-P.; et al. Electrophysiological and behavioral evidence that modulation of metabotropic glutamate receptor 4 with a new agonist reverses experimental parkinsonism. FASEB J. 2009, 23, 3619–3628. (In English) [Google Scholar] [CrossRef] [PubMed]
- Wierońska, J.M.; Stachowicz, K.; Acher, F.; Lech, T.; Pilc, A. Opposing efficacy of group III mGlu receptor activators, LSP1-2111 and AMN082, in animal models of positive symptoms of schizophrenia. Psychopharmacology 2012, 220, 481–494. (In English) [Google Scholar] [CrossRef] [PubMed]
- Wierońska, J.M.; Acher, F.C.; Sławińska, A.; Gruca, P.; Łasoń-Tyburkiewicz, M.; Papp, M.; Pilc, A. The antipsychotic-like effects of the mGlu group III orthosteric agonist, LSP1-2111, involves 5-HT₁A signalling. Psychopharmacology 2013, 227, 711–725. (In English) [Google Scholar] [CrossRef] [PubMed]
- Woźniak, M.; Gołembiowska, K.; Noworyta-Sokołowska, K.; Acher, F.; Cieślik, P.; Kusek, M.; Tokarski, K.; Pilc, A.; Wierońska, J.M. Neurochemical and behavioral studies on the 5-HT. Neuropharmacology 2017, 115, 149–165. (In English) [Google Scholar] [CrossRef] [PubMed]
- Woźniak, M.; Acher, F.; Marciniak, M.; Lasoń-Tyburkiewicz, M.; Gruca, P.; Papp, M.; Pilc, A.; Wierońska, J.M. Involvement of GABAB Receptor Signaling in Antipsychotic-like Action of the Novel Orthosteric Agonist of the mGlu4 Receptor, LSP4-2022. Curr. Neuropharmacol. 2016, 14, 413–426. (In English) [Google Scholar] [CrossRef] [PubMed]
- Cieślik, P.; Woźniak, M.; Rook, J.M.; Tantawy, M.N.; Conn, P.J.; Acher, F.; Tokarski, K.; Kusek, M.; Pilc, A.; Wierońska, J.M. Mutual activation of glutamatergic mGlu. Psychopharmacology 2018, 235, 2897–2913. (In English) [Google Scholar] [CrossRef] [PubMed]
- Sławińska, A.; Wierońska, J.M.; Stachowicz, K.; Marciniak, M.; Łasoń-Tyburkiewicz, M.; Gruca, P.; Papp, M.; Kusek, M.; Tokarski, K.; Doller, D.; et al. The antipsychotic-like effects of positive allosteric modulators of metabotropic glutamate mGlu4 receptors in rodents. Br. J. Pharmacol. 2013, 169, 1824–1839. (In English) [Google Scholar] [CrossRef]
- Kalinichev, M.; Le Poul, E.; Boléa, C.; Girard, F.; Campo, B.; Fonsi, M.; Royer-Urios, I.; Browne, S.E.; Uslaner, J.M.; Davis, M.J.; et al. Characterization of the novel positive allosteric modulator of the metabotropic glutamate receptor 4 ADX88178 in rodent models of neuropsychiatric disorders. J. Pharmacol. Exp. Ther. 2014, 350, 495–505. (In English) [Google Scholar] [CrossRef]
- Fazio, F.; Lionetto, L.; Molinaro, G.; Bertrand, H.O.; Acher, F.; Ngomba, R.T.; Notartomaso, S.; Curini, M.; Rosati, O.; Scarselli, P.; et al. Cinnabarinic acid, an endogenous metabolite of the kynurenine pathway, activates type 4 metabotropic glutamate receptors. Mol. Pharmacol. 2012, 81, 643–656. (In English) [Google Scholar] [CrossRef]
- Ulivieri, M.; Wierońska, J.M.; Lionetto, L.; Martinello, K.; Cieslik, P.; Chocyk, A.; Curto, M.; Di Menna, L.; Iacovelli, L.; Traficante, A.; et al. The Trace Kynurenine, Cinnabarinic Acid, Displays Potent Antipsychotic-Like Activity in Mice and Its Levels Are Reduced in the Prefrontal Cortex of Individuals Affected by Schizophrenia. Schizophr. Bull. 2020, 46, 1471–1481. (In English) [Google Scholar] [CrossRef]
- Cieślik, P.; Woźniak, M.; Kaczorowska, K.; Brański, P.; Burnat, G.; Chocyk, A.; Bobula, B.; Gruca, P.; Litwa, E.; Pałucha-Poniewiera, A.; et al. Negative Allosteric Modulators of mGlu. Front. Mol. Neurosci. 2018, 11, 316. (In English) [Google Scholar] [CrossRef]
- Ossowska, K.; Pietraszek, M.; Wardas, J.; Wolfarth, S. Potential antipsychotic and extrapyramidal effects of (R,S)-3,4-dicarboxyphenylglycine [(R,S)-3,4-DCPG], a mixed AMPA antagonist/mGluR8 agonist. Pol. J. Pharmacol. 2004, 56, 295–304. (In English) [Google Scholar] [PubMed]
- Pinheiro, P.S.; Mulle, C. Presynaptic glutamate receptors: Physiological functions and mechanisms of action. Nat. Rev. Neurosci. 2008, 9, 423–436. (In English) [Google Scholar] [CrossRef]
- Summa, M.; di Prisco, S.; Grilli, M.; Usai, C.; Marchi, M.; Pittaluga, A. Presynaptic mGlu7 receptors control GABA release in mouse hippocampus. Neuropharmacology 2013, 66, 215–224. (In English) [Google Scholar] [CrossRef] [PubMed]
- Kwon, H.B.; Castillo, P.E. Role of glutamate autoreceptors at hippocampal mossy fiber synapses. Neuron 2008, 60, 1082–1094. (In English) [Google Scholar] [CrossRef]
- Somogyi, P.; Dalezios, Y.; Luján, R.; Roberts, J.D.; Watanabe, M.; Shigemoto, R. High level of mGluR7 in the presynaptic active zones of select populations of GABAergic terminals innervating interneurons in the rat hippocampus. Eur. J. Neurosci. 2003, 17, 2503–2520. (In English) [Google Scholar] [CrossRef] [PubMed]
- Palazzo, E.; Marabese, I.; de Novellis, V.; Rossi, F.; Maione, S. Metabotropic Glutamate Receptor 7: From Synaptic Function to Therapeutic Implications. Curr. Neuropharmacol. 2016, 14, 504–513. (In English) [Google Scholar] [CrossRef]
- Schoepp, D.D.; Jane, D.E.; Monn, J.A. Pharmacological agents acting at subtypes of metabotropic glutamate receptors. Neuropharmacology 1999, 38, 1431–1476. (In English) [Google Scholar] [CrossRef]
- Dunn, H.A.; Patil, D.N.; Cao, Y.; Orlandi, C.; Martemyanov, K.A. Synaptic adhesion protein ELFN1 is a selective allosteric modulator of group III metabotropic glutamate receptors. Proc. Natl. Acad. Sci. USA 2018, 115, 5022–5027. (In English) [Google Scholar] [CrossRef] [PubMed]
- Stachniak, T.J.; Sylwestrak, E.L.; Scheiffele, P.; Hall, B.J.; Ghosh, A. Elfn1-Induced Constitutive Activation of mGluR7 Determines Frequency-Dependent Recruitment of Somatostatin Interneurons. J. Neurosci. 2019, 39, 4461–4474. (In English) [Google Scholar] [CrossRef]
- O’Connor, R.M.; Finger, B.C.; Flor, P.J.; Cryan, J.F. Metabotropic glutamate receptor 7: At the interface of cognition and emotion. Eur. J. Pharmacol. 2010, 639, 123–131. (In English) [Google Scholar] [CrossRef] [PubMed]
- Callaerts-Vegh, Z.; Beckers, T.; Ball, S.M.; Baeyens, F.; Callaerts, P.F.; Cryan, J.F.; Molnar, E.; D’Hooge, R. Concomitant deficits in working memory and fear extinction are functionally dissociated from reduced anxiety in metabotropic glutamate receptor 7-deficient mice. J. Neurosci. 2006, 26, 6573–6582. (In English) [Google Scholar] [CrossRef] [PubMed]
- Masugi, M.; Yokoi, M.; Shigemoto, R.; Muguruma, K.; Watanabe, Y.; Sansig, G.; Van Der Putten, H.; Nakanishi, S. Metabotropic glutamate receptor subtype 7 ablation causes deficit in fear response and conditioned taste aversion. J. Neurosci. 1999, 19, 955–963. (In English) [Google Scholar] [CrossRef]
- Stachowicz, K.; Brañski, P.; Kłak, K.; van der Putten, H.; Cryan, J.F.; Flor, P.J.; Andrzej, P. Selective activation of metabotropic G-protein-coupled glutamate 7 receptor elicits anxiolytic-like effects in mice by modulating GABAergic neurotransmission. Behav. Pharmacol. 2008, 19, 597–603. (In English) [Google Scholar] [CrossRef] [PubMed]
- Tassin, V.; Girard, B.; Chotte, A.; Fontanaud, P.; Rigault, D.; Kalinichev, M.; Perroy, J.; Acher, F.; Fagni, L.; Bertaso, F. Phasic and Tonic mGlu7 Receptor Activity Modulates the Thalamocortical Network. Front. Neural Circuits 2016, 10, 31. (In English) [Google Scholar] [CrossRef] [PubMed]
- Mitsukawa, K.; Yamamoto, R.; Ofner, S.; Nozulak, J.; Pescott, O.; Lukic, S.; Stoehr, N.; Mombereau, C.; Kuhn, R.; McAllister, K.H.; et al. A selective metabotropic glutamate receptor 7 agonist: Activation of receptor signaling via an allosteric site modulates stress parameters in vivo. Proc. Natl. Acad. Sci. USA 2005, 102, 18712–18717. (In English) [Google Scholar] [CrossRef] [PubMed]
- Darmani, N.A.; Martin, B.R.; Pandey, U.; Glennon, R.A. Do functional relationships exist between 5-HT1A and 5-HT2 receptors? Pharmacol. Biochem. Behav. 1990, 36, 901–906. (In English) [Google Scholar] [CrossRef]
- Suzuki, G.; Tsukamoto, N.; Fushiki, H.; Kawagishi, A.; Nakamura, M.; Kurihara, H.; Mitsuya, M.; Ohkubo, M.; Ohta, H. In vitro pharmacological characterization of novel isoxazolopyridone derivatives as allosteric metabotropic glutamate receptor 7 antagonists. J. Pharmacol. Exp. Ther. 2007, 323, 147–156. (In English) [Google Scholar] [CrossRef] [PubMed]
- Kalinichev, M.; Rouillier, M.; Girard, F.; Royer-Urios, I.; Bournique, B.; Finn, T.; Charvin, D.; Campo, B.; Le Poul, E.; Mutel, V.; et al. ADX71743, a potent and selective negative allosteric modulator of metabotropic glutamate receptor 7: In vitro and in vivo characterization. J. Pharmacol. Exp. Ther. 2012, 344, 624–636. (In English) [Google Scholar] [CrossRef]
- Duvoisin, R.M.; Zhang, C.; Ramonell, K. A novel metabotropic glutamate receptor expressed in the retina and olfactory bulb. J. Neurosci. 1995, 15, 3075–3083. (In English) [Google Scholar] [CrossRef]
- Robbins, M.J.; Starr, K.R.; Honey, A.; Soffin, E.M.; Rourke, C.; Jones, G.A.; Kelly, F.M.; Strum, J.; Melarange, R.A.; Harris, A.J.; et al. Evaluation of the mGlu8 receptor as a putative therapeutic target in schizophrenia. Brain Res. 2007, 1152, 215–227. (In English) [Google Scholar] [CrossRef] [PubMed]
- Shigemoto, R.; Kinoshita, A.; Wada, E.; Nomura, S.; Ohishi, H.; Takada, M.; Flor, P.J.; Neki, A.; Abe, T.; Nakanishi, S.; et al. Differential presynaptic localization of metabotropic glutamate receptor subtypes in the rat hippocampus. J. Neurosci. 1997, 17, 7503–7522. (In English) [Google Scholar] [CrossRef] [PubMed]
- Bushell, T.J.; Jane, D.E.; Tse, H.W.; Watkins, J.C.; Garthwaite, J.; Collingridge, G.L. Pharmacological antagonism of the actions of group II and III mGluR agonists in the lateral perforant path of rat hippocampal slices. Br. J. Pharmacol. 1996, 117, 1457–1462. (In English) [Google Scholar] [CrossRef]
- Gerlai, R.; Adams, B.; Fitch, T.; Chaney, S.; Baez, M. Performance deficits of mGluR8 knockout mice in learning tasks: The effects of null mutation and the background genotype. Neuropharmacology 2002, 43, 235–249. (In English) [Google Scholar] [CrossRef]
- Davis, M.J.; Duvoisin, R.M.; Raber, J. Related functions of mGlu4 and mGlu8. Pharmacol. Biochem. Behav. 2013, 111, 11–16. (In English) [Google Scholar] [CrossRef] [PubMed][Green Version]
- Fendt, M.; Bürki, H.; Imobersteg, S.; Van Der Putten, H.; McAllister, K.; Leslie, J.C.; Shaw, D.; Hölscher, C. The effect of mGlu8 deficiency in animal models of psychiatric diseases. Genes Brain Behav. 2010, 9, 33–44. (In English) [Google Scholar] [CrossRef] [PubMed]
- Duvoisin, R.M.; Zhang, C.; Pfankuch, T.F.; O’Connor, H.; Quraishi, S.; Raber, J.; Gayet-Primo, J. Increased measures of anxiety and weight gain in mice lacking the group III metabotropic glutamate receptor mGluR8. Eur. J. Neurosci. 2005, 22, 425–436. (In English) [Google Scholar] [CrossRef]
- Gasparini, F.; Bruno, V.; Battaglia, G.; Lukic, S.; Leonhardt, T.; Inderbitzin, W.; Laurie, D.; Sommer, B.; A Varney, M.; Hess, S.D.; et al. (R,S)-4-phosphonophenylglycine, a potent and selective group III metabotropic glutamate receptor agonist, is anticonvulsive and neuroprotective in vivo. J. Pharmacol. Exp. Ther. 1999, 289, 1678–1687. (In English) [Google Scholar] [PubMed]
- Erdmann, E.; Rupprecht, V.; Matthews, E.; Kukley, M.; Schoch, S.; Dietrich, D. Depression of release by mGluR8 alters Ca2+ dependence of release machinery. Cereb. Cortex 2012, 22, 1498–1509. (In English) [Google Scholar] [CrossRef] [PubMed][Green Version]
- Kammermeier, P.J. Constitutive activity of metabotropic glutamate receptor 7. BMC Neurosci. 2015, 16, 17. (In English) [Google Scholar] [CrossRef] [PubMed]


{kind=link}
| Group | Receptors/Splice Variants | Gene | Synapse Site | Expression in the CNS | Mechanism of Action |
|---|---|---|---|---|---|
| Group I | mGluR1 a,b,c,d,e,f | GRM1 | Mainly postsynaptic | Abundant in neurons and taste buds | PLC-dependent Ca2+ mobilization, stimulation of adenylyl cyclase (certain systems), activation of MAP kinases, PLD, mTOR/p70 S6 kinase |
| mGluR5 a,b | GRM5 | Abundant in neurons and astrocytes | |||
| Group II | mGluR2 | GRM2 | Presynaptic and postsynaptic | Abundant in neurons | Inhibition of adenylyl cyclase and voltage-dependent Ca2+ channels, activation of voltage-dependent K+ channels [16], MAPK and PI3 kinase pathway [17] |
| mGluR3 GRM3A2 GRM3A4 GRM3A2A3 | GRM3 | Abundant in neurons and astrocytes | |||
| Group III | mGluR4 | GRM4 | Mainly presynaptic (mGluR6 is postsynaptic in ON bipolar retinal cells) | Abundant in neurons (high in cerebellum) and taste buds | Inhibition of adenylyl cyclase, inhibition of voltage-dependent Ca2+ channels, activation of K+ channels [16], activation of cGMP phosphodiesterase [18] (mGluR6), MAPK and PI3 kinase pathway [17] |
| mGluR6 a,b,c | GRM6 | Retina | |||
| mGluR7 a,b,c,d,e | GRM7 | Abundant in neurons | |||
| mGluR8 a,b,c | GRM8 | Abundant in neurons (expression lower than mGluR4/7) |
| Target | Tested Drug | Animal Model | Results | References |
|---|---|---|---|---|
| mGluR1 | CFMTI | ketamine- and metAMPH-induced model | CFMTI reduced ketamine- and metAMPH-induced hyperlocomotion and drug-induced deficits in prepulse inhibition | [45] |
| CFMTI | MK-801-induced model | Ameliorated MK-801-disrupted social interactions | [45] | |
| FTIDC | metAMPH-induced model | Reduced metAMPH-evoked hyperlocomotion and ameliorated deficits in prepulse inhibition | [43] | |
| mGluR5 | MPEP | PCP-induced model | Potentiated the psychotomimetic, cognition impairing and prepulse inhibition-disruptive effects of PCP. | [44,46] |
| CHPG | MK-801-induced model Ketamine-induced model | CHPG reversed the cognitive-impairing effects of NMDA receptor antagonists and attenuated ketamine-induced locomotor activity, motor coordination and deficits in prepulse inhibition | [47,48] | |
| CDPPB | AMPH-induced model | Reduced AMPH-induced hyperlocomotion and ameliorated AMPH-disrupted prepulse inhibition | [49] | |
| CDPPB | MK-801-induced model | Attenuated MK-801-induced decrease in sucrose preference and deficits in cognitive flexibility | [50,51] | |
| ADX47273 | PCP- and AMPH-induced model | ADX47273 blocked PCP- and AMPH-induced locomotor activity and decreased extracellular dopamine level in the nucleus accumbens | [52] | |
| VU0409551 | AMPH-induced model MK-801-induced model | Reduced AMPH- and MK-801-induced hyperlocomotion | [53,54] |
| Tested Drug | Animal Model | Results | References |
|---|---|---|---|
| LY379268 | PCP- and AMPH-induced model | LY379268 reversed PCP and AMPH-induced hyperactivity and PCP-evoked behavioral deficits | [71] |
| LY354740 LY379268 | PCP- and AMPH-induced model | LY354740 and LY379268 attenuated PCP- but not AMPH-induced motor behaviors | [72] |
| LY379268 | AMPH-induced model | LY379268 attenuated AMPH-induced ambulations and rearing but not fine motor movements | [73] |
| LY379268 | PCP-induced model | LY379268 reduced PCP-induced falling, turning, and back pedaling in a dose-dependent manner but did not affect PCP-evoked forepaw treading | [74] |
| LY487379 LY379268 | PCP- and AMPH-induced model | LY487379 and LY379268 induced dose-dependent reductions in PCP- and AMPH-induced hyperlocomotor activity. LY487379 reversed AMPH-induced disruption of prepulse inhibition of the acoustic startle reflex. LY379268 when administered chronically failed to block AMPH- and PCP-induced hyperlocomotor activity. | [75] |
| LY354740 | PCP-induced psychosis | LY354740 induced anxiolytic-like effects and attenuated PCP-induced hyperlocomotion. It did not modify PCP-induced working memory deficits and had no effect on PCP-evoked amnesia. | [76] |
| LY354740 | PCP- and AMPH-induced model | LY354740 moderated effects of PCP on prepulse inhibition of acoustic startle | [77] |
| LY354740 LY487379 | PCP-induced psychosis | Acute pretreatment with LY354740 or LY487379 facilitated social discrimination in rats with PCP administration history without affecting total time spent in social interaction | [78] |
| LY379268 | PCP- and MK-801-induced model | LY379268 blocked PCP- and MK-801-induced hyperlocomotion in dopamine-deficient and control mice | [79] |
| LY354750 LY544344 | PCP-induced model | LY544344 but not LY354740 inhibited PCP-induced hyperlocomotion. | [80] |
| LY379268 | Ketamine-induced model | LY379268 reduced ketamine-evoked hyperlocomotion but it failed to restore prepulse inhibition deficits. Low dose (1 mg/kg) produced anxiolytic effects whereas a higher dose (3 mg/kg) appeared to be anxiogenic. | [81] |
| LY379268 | MK-801-induced model | LY379268 recovered the disrupted NMDA receptor expression induced by MK-801, enhanced NMDA-induced current in prefrontal neurons and reversed MK-801-induced NMDA receptor dysfunction via Akt/GSK-3β signaling. | [82] |
| LY379268 | Ketamine-induced model | Pretreatment with either systemic or local LY379268 blocked ketamine-induced glutamate, but no dopamine release in the mPFC. Systemic but not local administration blocked the effect of NMDA on evoking glutamate release. | [65] |
| LY379268 LY341495 | PCP- and LSD-induced model | LY341495 potentiated LSD-induced stimulus control, which was diminished in the presence of LY379268. In PCP-trained rats, LY341495 has no effect on stimulus control by an intermediate dose of PCP. In contrast, the training dose of PCP was significantly but incompletely antagonized by LY379268. | [83] |
| LY379268 | Ketamine-induced model | LY379268 reversed ketamine-induced hyperactivity and inhibited ketamine-evoked norepinephrine release in the ventral hippocampus. | [84] |
| LY379268 | PCP-induced model | LY379268 blocked PCP-evoked ambulatory activity and fine movements. | [85] |
| LY379268 | two-hit model (PCP-induced on the background of neuregulin 1 mutation) | PCP significantly reduced NMDA receptor and GABAA receptor binding density in the prefrontal cortex, hippocampus, and nucleus accumbens while LY379268 restored NMDA and GABAA receptors level | [86] |
| LY379268 | MK-801-induced model | LY379268 failed to improve MK-801-induced impairments in working memory. LY379268 augmented MK-801 potentiated gamma and high gamma oscillations but did not affect auditory-evoked gamma oscillation deficits caused by MK-801. | [87] |
| Target | Tested Drug | Animal Model | Results | References |
|---|---|---|---|---|
| mGluR4 | LSP1-2111 LSP4-2022 | MK-801-induced model AMPH-induced model | Reversed AMPH- and MK-801-induced hyperlocomotion | [131,134] |
| LSP1-2111 LSP4-2022 | DOI-induced model of hallucinations |
Antagonized
DOI-induced head twitches | [131,134] | |
|
LuAF21934
LuAF32615 | MK-801-induced model AMPH-induced model | Both compounds antagonized AMPH- and MK-801-induced hyperlocomotion | [136] | |
|
LuAF21934
LuAF32615 | DOI-induced model of hallucinations | Inhibited DOI-induced head twitches. Lu AF21934 was ineffective in mGluR4-/- mice | [136] | |
| ADX88178 | MK-801-induced model DOI-induced model of hallucinations | ADX88178 reduced MK-801-induced hyperlocomotion and DOI-evoked head twitches but had no effect in conditioned avoidance response test | [137] | |
| Cinnabarinic acid | MK-801-induced model | Reduced MK-801-induced glutamate release and hyperlocomotion | [139] | |
| mGluR7 | AMN082 | MK-801-induced model AMPH-induced model | AMN082 did not influence AMPH-induced hyperlocomotion but enhanced MK-801-induced hyperactivity | [131] |
| AMN082 | DOI-induced model of hallucinations | Increased number of the DOI-induced head twitches | [131] | |
| MMPIP ADX71743 | MK-801-induced model | Both drugs inhibited MK-801-induced hyperactivity, reversed MK-801-induced disturbances in novel object recognition, prepulse inhibition and spatial delayed alternation | [140] | |
| MMPIP ADX71743 | DOI-induced model of hallucinations | Inhibited DOI-induced head twitches | [140] | |
| mGluR8 | DCPG | PCP-induced model AMPH-induced model | Decreased amphetamine but not PCP-induced hyperactivity | [141] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kryszkowski, W.; Boczek, T. The G Protein-Coupled Glutamate Receptors as Novel Molecular Targets in Schizophrenia Treatment—A Narrative Review. J. Clin. Med. 2021, 10, 1475. https://doi.org/10.3390/jcm10071475
Kryszkowski W, Boczek T. The G Protein-Coupled Glutamate Receptors as Novel Molecular Targets in Schizophrenia Treatment—A Narrative Review. Journal of Clinical Medicine. 2021; 10(7):1475. https://doi.org/10.3390/jcm10071475
Chicago/Turabian StyleKryszkowski, Waldemar, and Tomasz Boczek. 2021. "The G Protein-Coupled Glutamate Receptors as Novel Molecular Targets in Schizophrenia Treatment—A Narrative Review" Journal of Clinical Medicine 10, no. 7: 1475. https://doi.org/10.3390/jcm10071475
APA StyleKryszkowski, W., & Boczek, T. (2021). The G Protein-Coupled Glutamate Receptors as Novel Molecular Targets in Schizophrenia Treatment—A Narrative Review. Journal of Clinical Medicine, 10(7), 1475. https://doi.org/10.3390/jcm10071475