
Respiratory Nasal Mucosa in Chronic Rhinosinusitis with Nasal Polyps versus COVID-19: Histopathology, Electron Microscopy Analysis and Assessing of Tissue Interleukin-33

 ,
, 
 , ,
, ,  , ,
, ,  ,
,  ,
,  ,
,
Abstract
1. Introduction
2. Materials and Methods
2.1. Study Design and Population
2.2. Sampling
2.3. Real-Time PCR (RT-PCR) SARS-CoV-2 (for the COVID-19 Study)
2.4. Histopathology
2.5. Scanning Electron Microscopy (SEM)
2.6. Transmission Electron Microscopy (TEM)
2.7. Assessing Tissue IL-33
2.7.1. Tissue Lysate Preparation and Protein Extraction
2.7.2. Quantitative Analysis of IL-33 by ELISA
3. Results

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Age | COVID-19 Study | ||
|---|---|---|---|
| CRSwNP Study | COVID-19 Patients | Non-COVID-19 Patients (Control) | |
| 18–29 | 2 | - | - |
| 30–49 | 12 | - | - |
| 50–69 | 9 | 3 | 2 |
| >70 | 2 | 9 | 3 |
| Total | 25 | 12 | 5 |
| Histopathology | |||
| Stromal edema (p < 0.0001) | 92% (n = 23/25) (Figure 1B) | 25% (n = 3/12) | - |
| 8% (n = 2/25) | - | - |
| 12% (n = 3/25) | 25% (n = 3/12) (Figure 2A,B) | - |
| 24% (n = 6/25) (Figure 1C) | - | - |
| 56% (n = 14/25) (Figure 1D) | - | - |
| - | - | - |
| Squamous metaplasia | 28% (n = 7/25) (Figure 1E,F) | 41.6% (n = 5/12) (Figure 2B–D) | - |
| Basal membrane thickening | 68% (n = 17/25) (Figure 1G,H) | 33.3% (n = 4/12) | - |
| Stromal fibrosis | 36% (n = 9/25) (Figure 1I,J) | - | - |
| Viral cytopathic effect | - | 8.3% (n = 1/12) (Figure 2C–E) | - |
| Microbial biofilms | 40% (n = 10/25) (Figure 1I,J) | - | - |
| Scanning electron microscopy analysis | |||
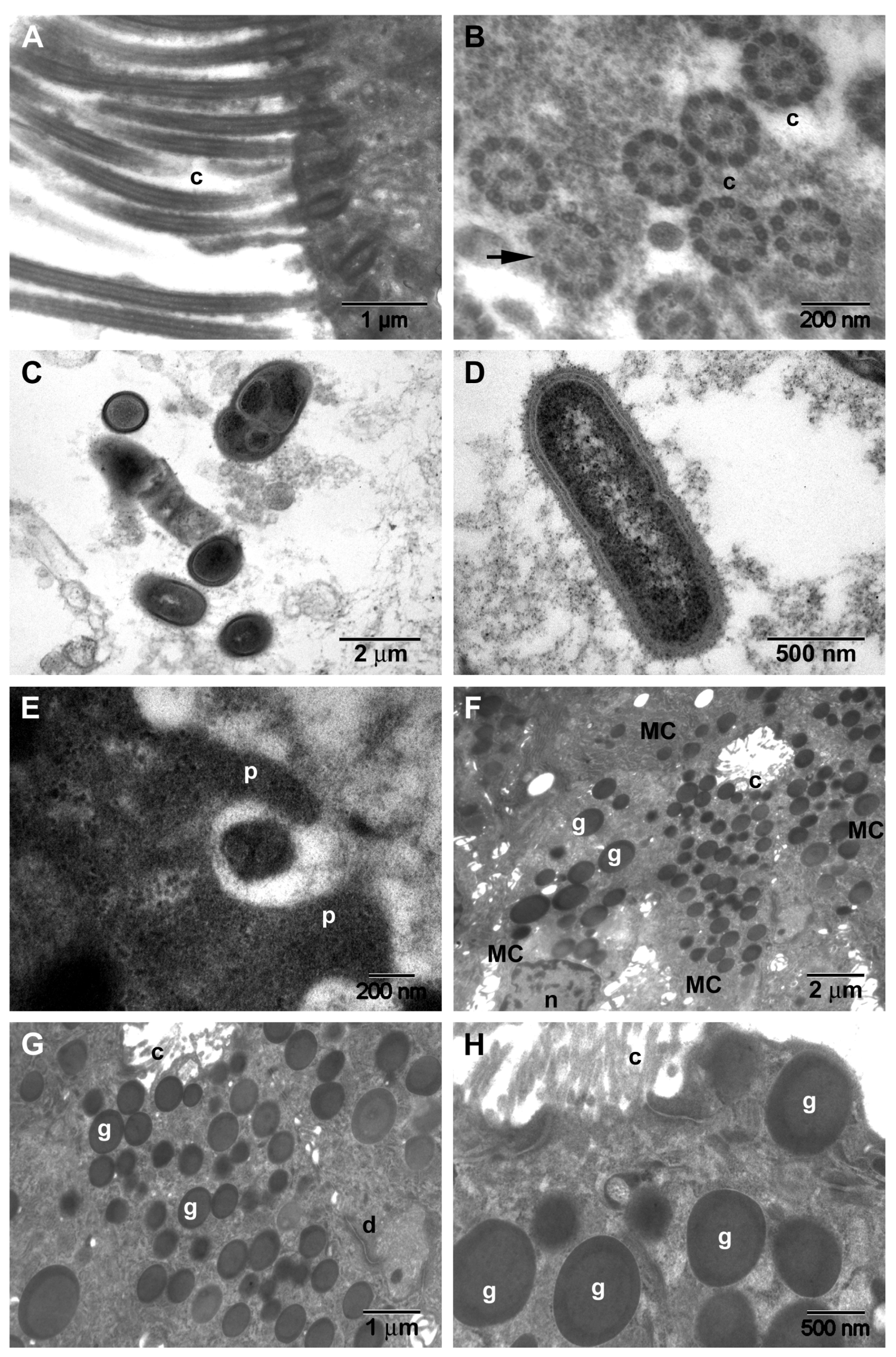
| Microbial biofilms | 56% (n = 14/25) | - | - |
| 36% (n = 9/25) | ||
| 20% (n = 5/25) (Figure 3A–C) | ||
| Microbial aggregates (without biofilm) (p = 0.0148, Chi-square 8.422) | 28% (n = 7/25) | 75% (n = 9/12) | 20% (n = 1/5) |
| 4% (n = 1/25) | 25% (n = 3/12) (Figure 6A–C) | 20% (n = 1/5) |
| 12% (n = 3/25) (Figure 3D–G) | 16.6% (n = 2/12) (Figure 6D–F) | - |
| 4% (n = 1/25) | 25% (n = 3/12) (Figure 6G–I) | - |
| 8% (n = 2/25) (Figure 3H) | 8.4% (n = 1/12) (Figure 6J,K) | - |
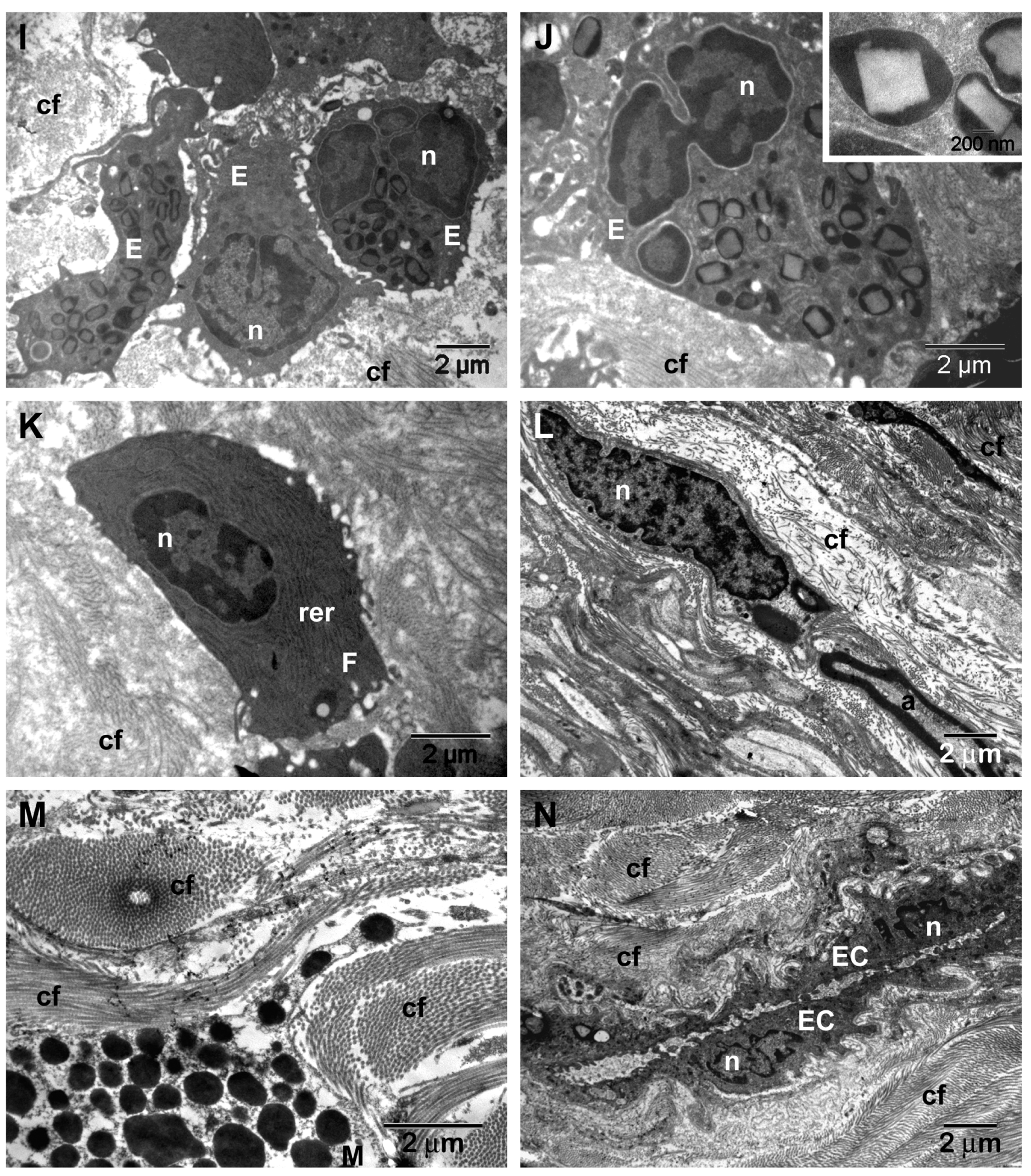
| Surface immune cells | 28% (n = 7/25) (Figure 4F) | 58.3% (n = 7/12) (Figure 6M–R) | 20% (n = 1/5) (Figure 5D) |
| Loss and dysfunction of cilia (p = 0.0274) | 76% (n = 19/25) (Figure 4F) | 33.3% (n = 4/12) | - |
| Transmission electron microscopy analysis | |||
| Ciliary abnormalities | 44% (n = 11/25) (Figure 7B) | 8.3% (n = 1/12) | - |
| Microbial biofilms | 36% (n = 9/25) (Figure 7C,D) | - | - |
| Hyperplasia of goblet cells (p = 0.0011) | 76% (n = 19/25) (Figure 7F–H) | 16.6% (n = 2/12) | - |
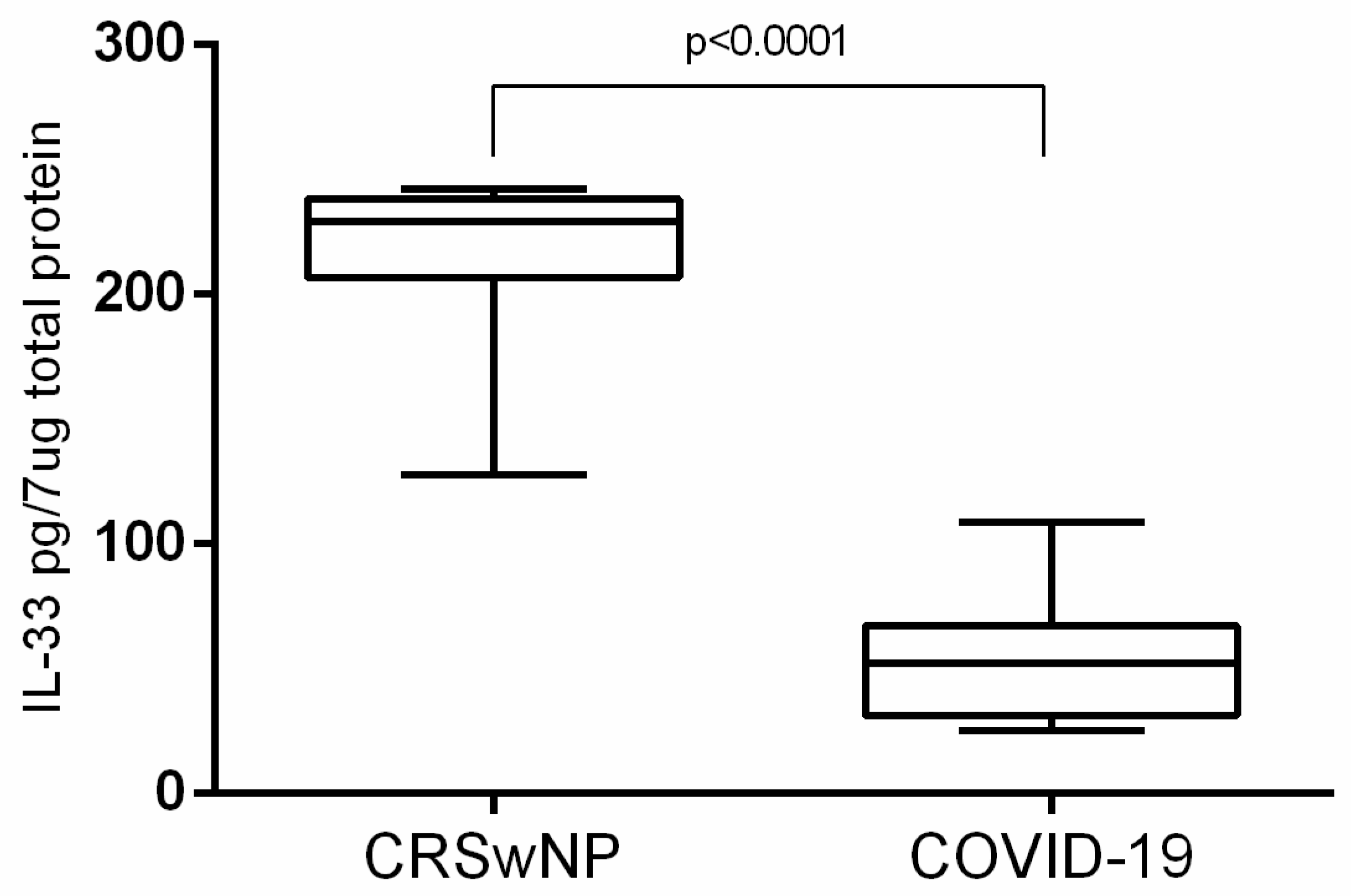
| Assessing of tissue interleukin-33 | |||
| 210.0 pg/7 μg total protein ± 8.327 (n = 25) | 52.77 pg/7 μg total protein ± 6.869 (n = 12) | ||
4. Discussion
4.1. Microscopic Inflammatory Findings in CRSwNP
4.2. Microscopic Inflammatory Findings of the Nasal Mucosa in COVID-19 Patients
4.3. Microbial Surface Communities
4.4. Surface Immune Cell Communities
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Gallo, O.; Locatello, L.G.; Mazzoni, A.; Novelli, L.; Annunziato, F. The central role of the nasal microenvironment in the transmission, modulation, and clinical progression of SARS-CoV-2 infection. Mucosal Immunol. 2021, 14, 305–316. [Google Scholar] [CrossRef] [PubMed]
- Keir, J.; Pedelty, L.; Swift, A.C. Biofilms in chronic rhinosinusitis: Systematic review and suggestions for future research. J. Laryngol. Otol. 2011, 125, 331–337. [Google Scholar] [CrossRef] [PubMed]
- Hong, H.; Liao, S.; Chen, F.; Yang, Q.; Wang, D.Y. Role of IL-25, IL-33, and TSLP in triggering united airway diseases toward type 2 inflammation. Allergy 2020, 75, 2794–2804. [Google Scholar] [CrossRef]
- Fokkens, W.J.; Lund, V.J.; Hopkins, C.; Hellings, P.W.; Kern, R.; Reitsma, S.; Toppila-Salmi, S.; Bernal-Sprekelsen, M.; Mullol, J.; Alobid, I.; et al. European position paper on rhinosinusitis and nasal polyps 2020. Rhinology 2020, 58, 1–464. [Google Scholar] [CrossRef] [PubMed]
- Laidlaw, T.M.; Buchheit, K.M. Biologics in chronic rhinosinusitis with nasal polyposis. Ann. Allergy Asthma Immunol. 2020, 124, 326–332. [Google Scholar] [CrossRef]
- Coronaviridae Study Group of the International Committee on Taxonomy of Viruses. The species Severe acute respiratory syndrome-related coronavirus: Classifying 2019-nCoV and naming it SARS-CoV-2. Nat. Microbiol. 2020, 5, 536–544. [Google Scholar] [CrossRef]
- Hou, Y.J.; Okuda, K.; Edwards, C.E.; Martinez, D.R.; Asakura, T.; Dinnon, K.H., III; Kato, T.; Lee, R.E.; Yount, B.L.; Macenik, T.M.; et al. SARS-CoV-2 Reverse Genetics Reveals a Variable Infection Gradient in the Respiratory Tract. Cell 2020, 182, 429–446.e14. [Google Scholar] [CrossRef]
- Ho, J.; Earls, P.; Harvey, R.J. Systemic biomarkers of eosinophilic chronic rhinosinusitis. Curr. Opin. Allergy Clin. Immunol. 2020, 20, 23–29. [Google Scholar] [CrossRef] [PubMed]
- Cayrol, C.; Girard, J.P. Interleukin-33 (IL-33): A nuclear cytokine from the IL-1 family. Immunol. Rev. 2018, 281, 154–168. [Google Scholar] [CrossRef] [PubMed]
- Werder, R.B.; Zhang, V.; Lynch, J.P.; Snape, N.; Upham, J.W.; Spann, K.; Phipps, S. Chronic IL-33 expression predisposes to virus-induced asthma exacerbations by increasing type 2 inflammation and dampening antiviral immunity. J. Allergy Clin. Immunol. 2018, 141, 1607–1619.e9. [Google Scholar] [CrossRef]
- Kim, D.K.; Jin, H.R.; Eun, K.M.; Mo, J.H.; Cho, S.H.; Oh, S.; Cho, D.; Kim, D.W. The role of interleukin-33 in chronic rhinosinusitis. Thorax 2017, 72, 635–645. [Google Scholar] [CrossRef]
- Zizzo, G.; Cohen, P.L. Imperfect storm: Is interleukin-33 the Achilles heel of COVID-19? Lancet Rheumatol. 2020, 2, e779–e790. [Google Scholar] [CrossRef]
- Liang, Y.; Ge, Y.; Sun, J. IL-33 in COVID-19: Friend or foe? Cell. Mol. Immunol. 2021, 18, 1602–1604. [Google Scholar] [CrossRef] [PubMed]
- Yalcin, A.D.; Yalcin, A.N. Future perspective: Biologic agents in patients with severe COVID-19. Immunopharmacol. Immunotoxicol. 2021, 43, 1–7. [Google Scholar] [CrossRef] [PubMed]
- WHO. Infection Prevention and Control for the Safe Management of a Dead Body in the Context of COVID-19. 2020. Available online: https://apps.who.int/iris/bitstream/handle/10665/331538/WHO-COVID-19-lPC_DBMgmt-2020.1-eng.pdf (accessed on 30 March 2021).
- Centers for Disease Control and Prevention, Collection and Submission of Postmortem Specimens from Deceased Persons with Known or Suspected COVID-19, (Interim Guidance). 2020. Available online: https://www.cdc.gov/coronavirus/2019-ncov/hcp/guidance-postmortem-specimens.html (accessed on 30 March 2021).
- European Centre for Disease Prevention and Control. Considerations Related to the Safe Handling of Bodies of Deceased Persons with Suspected or Confirmed COVID-19. Available online: https://www.ecdc.europa.eu/en/publications-data/considerations-related-safe-handling-bodies-deceased-persons-suspected-or (accessed on 30 March 2021).
- National Institute for Public Health from Romania. Available online: https://www.cnscbt.ro/index.php/info-medical/2047-metodologia-de-supraveghere-a-covid-19-actualizare-23-10-2020-1/file (accessed on 30 March 2021).
- Mladina, R.; Poje, G.; Vuković, K.; Ristić, M.; Musić, S. Biofilm in nasal polyps. Rhinology 2008, 46, 302–307. [Google Scholar] [PubMed]
- Cocuzza, S.; Maniaci, A.; Di Luca, M.; La Mantia, I.; Grillo, C.; Spinato, G.; Motta, G.; Testa, D.; Ferlito, S. Long-term results of nasal surgery: Comparison of mini-invasive turbinoplasty. J. Biol. Regul. Homeost. Agents 2020, 34, 1203–1208. [Google Scholar] [CrossRef] [PubMed]
- Wang, M.; Bu, X.; Luan, G.; Lin, L.; Wang, Y.; Jin, J.; Zhang, L.; Wang, C. Distinct type 2-high inflammation associated molecular signatures of chronic rhinosinusitis with nasal polyps with comorbid asthma. Clin. Transl. Allergy 2020, 10, 26. [Google Scholar] [CrossRef] [PubMed]
- Lou, H.; Meng, Y.; Piao, Y.; Zhang, N.; Bachert, C.; Wang, C.; Zhang, L. Cellular phenotyping of chronic rhinosinusitis with nasal polyps. Rhinology 2016, 54, 150–159. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Lou, H.; Zhang, N.; Bachert, C.; Zhang, L. Highlights of eosinophilic chronic rhinosinusitis with nasal polyps in definition, prognosis, and advancement. Int. Forum Allergy Rhinol. 2018, 8, 1218–1225. [Google Scholar] [CrossRef]
- Ishitoya, J.; Sakuma, Y.; Tsukuda, M. Eosinophilic chronic rhinosinusitis in Japan. Allergol. Int. 2010, 59, 239–245. [Google Scholar] [CrossRef]
- McHugh, T.; Snidvongs, K.; Xie, M.; Banglawala, S.; Sommer, D. High tissue eosinophilia as a marker to predict recurrence for eosinophilic chronic rhinosinusitis: A systematic review and meta-analysis. Int. Forum Allergy Rhinol. 2018, 8, 1421–1429. [Google Scholar] [CrossRef] [PubMed]
- Pyo, J.S.; Kim, S.J. Relationship between histologic changes and inflammatory markers in chronic rhinosinusitis. Int. J. Clin. Exp. Pathol. 2021, 14, 501–507. [Google Scholar] [PubMed]
- Tu, Y.; Liu, J.; Li, T.; Zhou, X.; Tan, K.S.; Ong, H.H.; Sun, K.; Ouyang, Y.; Liang, X.; Ong, Y.K.; et al. Mucus composition abnormalities in sinonasal mucosa of chronic rhinosinusitis with and without nasal polyps. Inflammation 2021, 1–12. [Google Scholar] [CrossRef]
- Whitsett, J.A. Airway Epithelial Differentiation and Mucociliary Clearance. Ann. Am. Thorac. Soc. 2018, 15, S143–S148. [Google Scholar] [CrossRef]
- Perić, A.; Stoiljkov, M.; Đokić, D.; Đurđević, B.V. Epithelial Squamous Metaplasia and Dysplasia in Inflammatory Nasal Polyps: An Observational Study. Ear Nose Throat J. 2021, 100, NP120–NP124. [Google Scholar] [CrossRef] [PubMed]
- Brescia, G.; Contro, G.; Giacomelli, L.; Barion, U.; Frigo, A.C.; Marioni, G. Blood Eosinophilic and Basophilic Trends in Recurring and Non-Recurring Eosinophilic Rhinosinusitis with Nasal Polyps. Am. J. Rhinol. Allergy 2021, 35, 296–301. [Google Scholar] [CrossRef]
- Ren, H.L.; Li, J.D.; Yue, F.S.; Sun, J.L.; Rebeiz, E.E.; Theoharides, T.C. Nasal cytology with emphasis on mast cells can improve the diagnosis and treatment of chronic rhinosinusitis. Chin. Med. J. 2019, 132, 2237–2241. [Google Scholar] [CrossRef]
- Dwyer, D.F.; Ordovas-Montanes, J.M.; Buchheit, K.M.; Lai, J.; Katz, H.R.; Bhattacharyya, N.; Shalek, A.K.; Laidlaw, T.M.; Boyce, J.A.; Barrett, N.A. Unlocking mast cell diversity in human nasal polyps. J. Immunol. 2017, 198, 145–149. [Google Scholar]
- Liu, T.; Kanaoka, Y.; Barrett, N.A.; Feng, C.; Garofalo, D.; Lai, J.; Buchheit, K.; Bhattacharya, N.; Laidlaw, T.M.; Katz, H.R.; et al. Aspirin-Exacerbated Respiratory Disease Involves a Cysteinyl Leukotriene-Driven IL-33-Mediated Mast Cell Activation Pathway. J. Immunol. 2015, 195, 3537–3545. [Google Scholar] [CrossRef]
- Shaw, J.L.; Ashoori, F.; Fakhri, S.; Citardi, M.J.; Luong, A. Increased percentage of mast cells within sinonasal mucosa of chronic rhinosinusitis with nasal polyp patients independent of atopy. Int. Forum Allergy Rhinol. 2012, 2, 233–240. [Google Scholar] [CrossRef]
- Galli, J.; Calo, L.; Ardito, F.; Imperiali, M.; Bassotti, E.; Passali, G.C.; La Torre, G.; Paludetti, G.; Fadda, G. Damage to ciliated epithelium in chronic rhinosinusitis: What is the role of bacterial biofilms? Ann. Otol. Rhinol. Laryngol. 2008, 117, 902–909. [Google Scholar] [CrossRef]
- Ma, Y.; Tian, P.; Zhong, H.; Wu, F.; Zhang, Q.; Liu, X.; Dang, H.; Chen, Q.; Zou, H.; Zheng, Y. WDPCP Modulates Cilia Beating Through the MAPK/ERK Pathway in Chronic Rhinosinusitis with Nasal Polyps. Front. Cell Dev. Biol. 2021, 8, 1930. [Google Scholar] [CrossRef] [PubMed]
- Soares, C.D.; Carvalho, R.A.; Carvalho, K.A.; Carvalho, M.G.; Almeida, O.P. Letter to Editor: Oral lesions in a patient with Covid-19. Med. Oral Patol. Oral Cir. Bucal 2020, 25, e563–e564. [Google Scholar] [CrossRef]
- Mohanty, S.K.; Satapathy, A.; Naidu, M.M.; Mukhopadhyay, S.; Sharma, S.; Barton, L.M.; Stroberg, E.; Duval, E.J.; Pradhan, D.; Tzankov, A.; et al. Severe acute respiratory syndrome coronavirus-2 (SARS-CoV-2) and coronavirus disease 19 (COVID-19)—Anatomic pathology perspective on current knowledge. Diagn. Pathol. 2020, 15, 103. [Google Scholar] [CrossRef] [PubMed]
- Pritt, B.S.; Aubry, M.C. Histopathology of viral infections of the lung. Semin. Diagn. Pathol. 2017, 34, 510–517. [Google Scholar] [CrossRef]
- Bradley, B.T.; Maioli, H.; Johnston, R.; Chaudhry, I.; Fink, S.L.; Xu, H.; Najafian, B.; Deutsch, G.; Lacy, J.M.; Williams, T.; et al. Histopathology and ultrastructural findings of fatal COVID-19 infections in Washington State: A case series. Lancet 2020, 396, 320–332. [Google Scholar] [CrossRef]
- Zhu, N.; Wang, W.; Liu, Z.; Liang, C.; Wang, W.; Ye, F.; Huang, B.; Zhao, L.; Wang, H.; Zhou, W.; et al. Morphogenesis and cytopathic effect of SARS-CoV-2 infection in human airway epithelial cells. Nat. Commun. 2020, 11, 3910. [Google Scholar] [CrossRef]
- Chilvers, M.A.; McKean, M.; Rutman, A.; Myint, B.S.; Silverman, M.; O’Callaghan, C. The effects of coronavirus on human nasal ciliated respiratory epithelium. Eur. Respir. J. 2001, 18, 965–970. [Google Scholar] [CrossRef]
- Fajnzylber, J.; Regan, J.; Coxen, K.; Corry, H.; Wong, C.; Rosenthal, A.; Worrall, D.; Giguel, F.; Piechocha-Trocha, A.; Atyeo, C.; et al. SARS-CoV-2 viral load is associated with increased disease severity and mortality. Nat. Commun. 2020, 11, 5493. [Google Scholar] [CrossRef]
- Zou, L.; Ruan, F.; Huang, M.; Liang, L.; Huang, H.; Hong, Z.; Yu, J.; Kang, M.; Song, Y.; Xia, J.; et al. SARS-CoV-2 viral load in upper respiratory specimens of infected patients. N. Engl. J. Med. 2020, 382, 1177–1179. [Google Scholar] [CrossRef] [PubMed]
- Jeican, I.I.; Aluaș, M.; Lazăr, M.; Barbu-Tudoran, L.; Gheban, D.; Inișca, P.; Albu, C.; Tripon, S.; Albu, S.; Siserman, C.; et al. Evidence of SARS-CoV-2 Virus in the Middle Ear of Deceased COVID-19 Patients. Diagnostics 2021, 11, 1535. [Google Scholar] [CrossRef]
- Dimitri-Pinheiro, S.; Soares, R.; Barata, P. The Microbiome of the Nose-Friend or Foe? Allergy Rhinol. 2020, 11, 2152656720911605. [Google Scholar] [CrossRef] [PubMed]
- Jeican, I.I.; Barbu-Tudoran, L.; Florea, A.; Flonta, M.; Trombitas, V.; Apostol, A.; Dumitru, M.; Aluaș, M.; Junie, L.M.; Albu, S. Chronic Rhinosinusitis: MALDI-TOF Mass Spectrometry Microbiological Diagnosis and Electron Microscopy Analysis; Experience of the 2nd Otorhinolaryngology Clinic of Cluj-Napoca, Romania. J. Clin. Med. 2020, 9, 3973. [Google Scholar] [CrossRef] [PubMed]
- Zhong, H.; Wang, Y.; Shi, Z.; Zhang, L.; Ren, H.; He, W.; Zhang, Z.; Zhu, A.; Zhao, J.; Xiao, F.; et al. Characterization of respiratory microbial dysbiosis in hospitalized COVID-19 patients. Cell Discov. 2021, 7, 23. [Google Scholar] [CrossRef]
- Wolter, N.; Tempia, S.; Cohen, C.; Madhi, S.A.; Venter, M.; Moyes, J.; Walaza, S.; Malope-Kgokong, B.; Groome, M.; du Plessis, M.; et al. High nasopharyngeal pneumococcal density, increased by viral coinfection, is associated with invasive pneumococcal pneumonia. J. Infect. Dis. 2014, 210, 1649–1657. [Google Scholar] [CrossRef]
- Westblade, L.F.; Simon, M.S.; Satlin, M.J. Bacterial coinfections in coronavirus disease 2019. Trends Microbiol. 2021. [Google Scholar] [CrossRef]
- Rawson, T.M.; Wilson, R.C.; Holmes, A. Understanding the role of bacterial and fungal infection in COVID-19. Clin. Microbiol. Infect. 2021, 27, 9–11. [Google Scholar] [CrossRef]
- Danielsen, A.K.; Eskeland, Ø.; Fridrich-Aas, K.; CecilieOrszagh, V.; Bachmann-Harildstad, G.; Burum-Auensen, E. Bacterial biofilms in chronic rhinosinusitis; distribution and prevalence. Acta Otolaryngol. 2016, 136, 109–112. [Google Scholar] [CrossRef]
- Healy, D.Y.; Leid, J.G.; Sanderson, A.R.; Hunsaker, D.H. Biofilms with fungi in chronic rhinosinusitis. Otolaryngol. Head Neck Surg. 2008, 138, 641–647. [Google Scholar] [CrossRef]
- Bezerra, T.F.; Padua, F.G.; Gebrim, E.M.; Saldiva, P.H.; Voegels, R.L. Biofilms in chronic rhinosinusitis with nasal polyps. Otolaryngol. Head Neck Surg. 2011, 144, 612–616. [Google Scholar] [CrossRef]
- Cutcheon, J.M.; Southam, G. Advanced biofilm staining techniques for TEM and SEM in geomicrobiology: Implications for visualizing EPS architecture, mineral nucleation, and microfossil generation. Chem. Geol. 2018, 498, 115–127. [Google Scholar] [CrossRef]
- Relucenti, M.; Familiari, G.; Donfrancesco, O.; Taurino, M.; Li, X.; Chen, R.; Artini, M.; Papa, R.; Selan, L. Microscopy Methods for Biofilm Imaging: Focus on SEM and VP-SEM Pros and Cons. Biology 2021, 10, 51. [Google Scholar] [CrossRef]
- Achinas, S.; Yska, S.K.; Charalampogiannis, N.; Krooneman, J.; Euverink, G.J.W. A Technological Understanding of Biofilm Detection Techniques: A Review. Materials 2020, 13, 3147. [Google Scholar] [CrossRef]
- Braun, H.; Buzina, W.; Freudenschuss, K.; Beham, A.; Stammberger, H. ‘Eosinophilic fungal rhinosinusitis’: A common disorder in Europe? Laryngoscope 2003, 113, 264–269. [Google Scholar] [CrossRef]
- Sasama, J.; Sherris, D.A.; Shin, S.H.; Kephart, G.M.; Kern, E.B.; Ponikau, J.U. New paradigm for the roles of fungi and eosinophils in chronic rhinosinusitis. Curr. Opin. Otolaryngol. Head Neck Surg. 2005, 13, 2–8. [Google Scholar] [CrossRef] [PubMed]
- Ali Alharbi, S. Isolation of ultrasmall (filterable) bacteria from patients suffering from ME, and patients and staff of a paediatric hospital. Saudi J. Biol. Sci. 2020, 27, 1566–1568. [Google Scholar] [CrossRef]
- Thoulouze, M.I.; Alcover, A. Can viruses form biofilms? Trends Microbiol. 2011, 19, 257–262. [Google Scholar] [CrossRef] [PubMed]
- Maali, Y.; Journo, C.; Mahieux, R.; Dutartre, H. Microbial Biofilms: Human T-cell Leukemia Virus Type 1 First in Line for Viral Biofilm but Far Behind Bacterial Biofilms. Front. Microbiol. 2020, 11, 2041. [Google Scholar] [CrossRef]
- Bullock, H.A.; Goldsmith, C.S.; Miller, S.E. Best practices for correctly identifying coronavirus by transmission electron microscopy. Kidney Int. 2021, 99, 824–827. [Google Scholar] [CrossRef]
- Dittmayer, C.; Meinhardt, J.; Radbruch, H.; Radke, J.; Heppner, B.I.; Heppner, F.L.; Stenzel, W.; Holland, G.; Laue, M. Why misinterpretation of electron micrographs in SARS-CoV-2-infected tissue goes viral. Lancet 2020, 396, e64–e65. [Google Scholar] [CrossRef]
- Pérez, A.; Torregrosa, I.; D’Marco, L.; Juan, I.; Terradez, L.; Solís, M.Á.; Moncho, F.; Carda-Batalla, C.; Forner, M.J.; Gorriz, J.L. IgA-Dominant Infection-Associated Glomerulonephritis Following SARS-CoV-2 Infection. Viruses 2021, 13, 587. [Google Scholar] [CrossRef] [PubMed]
- Meinhardt, J.; Radke, J.; Dittmayer, C.; Franz, J.; Thomas, C.; Mothes, R.; Laue, M.; Schneider, J.; Brünink, S.; Greuel, S.; et al. Olfactory transmucosal SARS-CoV-2 invasion as a port of central nervous system entry in individuals with COVID-19. Nat. Neurosci. 2021, 24, 168–175. [Google Scholar] [CrossRef]
- Saraste, J.; Prydz, K. Assembly and Cellular Exit of Coronaviruses: Hijacking an Unconventional Secretory Pathway from the Pre-Golgi Intermediate Compartment via the Golgi Ribbon to the Extracellular Space. Cells 2021, 10, 503. [Google Scholar] [CrossRef] [PubMed]
- Wu, R.Q.; Zhang, D.F.; Tu, E.; Chen, Q.M.; Chen, W. The mucosal immune system in the oral cavity-an orchestra of T cell diversity. Int. J. Oral Sci. 2014, 6, 125–132. [Google Scholar] [CrossRef] [PubMed]
- Chang, S.Y.; Ko, H.J.; Kweon, M.N. Mucosal dendritic cells shape mucosal immunity. Exp. Mol. Med. 2014, 46, e84. [Google Scholar] [CrossRef]
- Jahnsen, F.L.; Gran, E.; Haye, R.; Brandtzaeg, P. Human nasal mucosa contains antigen-presenting cells of strikingly different functional phenotypes. Am. J. Respir. Cell Mol. Biol. 2004, 30, 31–37. [Google Scholar] [CrossRef]
- Banchereau, J.; Steinman, R.M. Dendritic cells and the control of immunity. Nature 1998, 392, 245–252. [Google Scholar] [CrossRef]
- Campana, P.; Parisi, V.; Leosco, D.; Bencivenga, D.; Della Ragione, F.; Borriello, A. Dendritic Cells and SARS-CoV-2 Infection: Still an Unclarified Connection. Cells 2020, 9, 2046. [Google Scholar] [CrossRef]
- Xiong, Y.; Liu, Y.; Cao, L.; Wang, D.; Guo, M.; Jiang, A.; Guo, D.; Hu, W.; Yang, J.; Tang, Z.; et al. Transcriptomic characteristics of bronchoalveolar lavage fluid and peripheral blood mononuclear cells in COVID-19 patients. Emerg. Microbes Infect. 2020, 9, 761–770. [Google Scholar] [CrossRef]
- Xing, F.; Wang, J.; Hu, M.; Yu, Y.; Chen, G.; Liu, J. Comparison of immature and mature bone marrow-derived dendritic cells by atomic force microscopy. Nanoscale Res. Lett. 2011, 6, 455. [Google Scholar] [CrossRef]
- Kim, M.K.; Kim, J. Properties of immature and mature dendritic cells: Phenotype, morphology, phagocytosis, and migration. RSC Adv. 2019, 9, 11230–11238. [Google Scholar] [CrossRef]
- Dumortier, H.; van Mierlo, G.J.; Egan, D.; van Ewijk, W.; Toes, R.E.; Offringa, R.; Melief, C.J. Antigen presentation by an immature myeloid dendritic cell line does not cause CTL deletion in vivo, but generates CD8+ central memory-like T cells that can be rescued for full effector function. J. Immunol. 2005, 175, 855–863. [Google Scholar] [CrossRef] [PubMed]
- Newberry, R. Lymphocytes. In Encyclopedia of Gastroenterology; Johnson, L.R., Ed.; Elsevier: Amsterdam, The Netherlands, 2004; pp. 570–575. [Google Scholar]
- Cano, R.L.E.; Lopera, H.D.E. Introduction to T and B lymphocytes. In Autoimmunity: From Bench to Bedside; Anaya, J.M., Shoenfeld, Y., Rojas-Villarraga, A., et al., Eds.; El Rosario University Press: Bogota, Colombia, 2013; Chapter 5. Available online: https://www.ncbi.nlm.nih.gov/books/NBK459471/ (accessed on 30 March 2021).
- Lin, W.; Suo, Y.; Deng, Y.; Fan, Z.; Zheng, Y.; Wei, X.; Chu, Y. Morphological change of CD4(+) T cell during contact with DC modulates T-cell activation by accumulation of F-actin in the immunology synapse. BMC Immunol. 2015, 16, 49. [Google Scholar] [CrossRef]
- Liew, F.Y.; Girard, J.P.; Turnquist, H.R. Interleukin-33 in health and disease. Nat. Rev. Immunol. 2016, 16, 676–689. [Google Scholar] [CrossRef] [PubMed]
- Ishinaga, H.; Kitano, M.; Toda, M.; D’Alessandro-Gabazza, C.N.; Gabazza, E.C.; Shah, S.A.; Takeuchi, K. Interleukin-33 induces mucin gene expression and goblet cell hyperplasia in human nasal epithelial cells. Cytokine 2017, 90, 60–65. [Google Scholar] [CrossRef] [PubMed]
- Dogan, M.; Sahin, M.; Yenisey, C. Increased TSLP, IL-33, IL-25, IL-19, IL 21 and amphiregulin (AREG) levels in chronic rhinosinusitis with nasal polyp. Eur. Arch. Otorhinolaryngol. 2019, 276, 1685–1691. [Google Scholar] [CrossRef] [PubMed]
- Song, W.; Wang, C.; Zhou, J.; Pan, S.; Lin, S. IL-33 Expression in Chronic Rhinosinusitis with Nasal Polyps and Its Relationship with Clinical Severity. ORL 2017, 79, 323–330. [Google Scholar] [CrossRef]
- Soyka, M.B.; Holzmann, D.; Basinski, T.M.; Wawrzyniak, M.; Bannert, C.; Bürgler, S.; Akkoc, T.; Treis, A.; Rückert, B.; Akdis, M.; et al. The Induction of IL-33 in the Sinus Epithelium and Its Influence on T-Helper Cell Responses. PLoS ONE 2015, 10, e0123163. [Google Scholar] [CrossRef]
- Huang, C.; Wang, Y.; Li, X.; Ren, L.; Zhao, J.; Hu, Y.; Zhang, L.; Fan, G.; Xu, J.; Gu, X.; et al. Clinical features of patients infected with 2019 novel coronavirus in Wuhan, China. Lancet 2020, 395, 497–506. [Google Scholar] [CrossRef]
- Del Valle, D.M.; Kim-Schulze, S.; Huang, H.-H.; Beckmann, N.D.; Nirenberg, S.; Wang, B.; Lavin, Y.; Swartz, T.H.; Madduri, D.; Stock, A.; et al. An inflammatory cytokine signature predicts COVID-19 severity and survival. Nat. Med. 2020, 26, 1636–1643. [Google Scholar] [CrossRef]
- Stanczak, M.A.; Sanin, D.E.; Apostolova, P.; Nerz, G.; Lampaki, D.; Hofmann, M.; Steinmann, D.; Krohn-Grimberghe, M.; Thimme, R.; Mittler, G.; et al. IL-33 expression in response to SARS-CoV-2 correlates with seropositivity in COVID-19 convalescent individuals. Nat. Commun. 2021, 12, 2133. [Google Scholar] [CrossRef] [PubMed]
- Zeng, Z.; Hong, X.Y.; Li, Y.; Chen, W.; Ye, G.; Li, Y.; Luo, Y. Serum-soluble ST2 as a novel biomarker reflecting inflammatory status and illness severity in patients with COVID-19. Biomark. Med. 2020, 14, 1619–1629. [Google Scholar] [CrossRef] [PubMed]













| CRSwNP Study | COVID-19 Study | ||
|---|---|---|---|
| Case Group | Control Group | ||
| Number of patients | 25 | 12 | 5 |
| Medical institution of the patients | 2nd Otorhinolaryngology Clinic, University Clinical Hospital of Railway Company, Cluj-Napoca, Romania | Department of Pathology, County Emergency Hospital Deva/Institute of Legal Medicine Cluj-Napoca | Department of Pathology, County Emergency Hospital Deva/Institute of Legal Medicine Cluj-Napoca |
| Inclusion criteria | Patients undergoing functional endoscopic sinus surgery for CRSwNP, diagnosed according to the EPOS 2020 criteria [4] | Deceased patients with antemortem COVID-19 diagnosis confirmed through combined throat/nasal sampling RT-PCR SARS-CoV-2 test | Deceased patients, with antemortem negative RT-PCR SARS-CoV-2 test performed through combined throat/nasal sampling |
| Exclusion criteria |
|
|
|
| Research analysis | Histopathology, electron microscopy analysis, and assessing of tissue interleukin-33 | Tissue RT-PCR SARS-CoV-2, histopathology, electron microscopy analysis, and assessing of tissue interleukin-33 | Tissue RT-PCR SARS-CoV-2, histopathology, and electron microscopy analysis |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Jeican, I.I.; Gheban, D.; Barbu-Tudoran, L.; Inișca, P.; Albu, C.; Ilieș, M.; Albu, S.; Vică, M.L.; Matei, H.V.; Tripon, S.; et al. Respiratory Nasal Mucosa in Chronic Rhinosinusitis with Nasal Polyps versus COVID-19: Histopathology, Electron Microscopy Analysis and Assessing of Tissue Interleukin-33. J. Clin. Med. 2021, 10, 4110. https://doi.org/10.3390/jcm10184110
Jeican II, Gheban D, Barbu-Tudoran L, Inișca P, Albu C, Ilieș M, Albu S, Vică ML, Matei HV, Tripon S, et al. Respiratory Nasal Mucosa in Chronic Rhinosinusitis with Nasal Polyps versus COVID-19: Histopathology, Electron Microscopy Analysis and Assessing of Tissue Interleukin-33. Journal of Clinical Medicine. 2021; 10(18):4110. https://doi.org/10.3390/jcm10184110
Chicago/Turabian StyleJeican, Ionuț Isaia, Dan Gheban, Lucian Barbu-Tudoran, Patricia Inișca, Camelia Albu, Maria Ilieș, Silviu Albu, Mihaela Laura Vică, Horea Vladi Matei, Septimiu Tripon, and et al. 2021. "Respiratory Nasal Mucosa in Chronic Rhinosinusitis with Nasal Polyps versus COVID-19: Histopathology, Electron Microscopy Analysis and Assessing of Tissue Interleukin-33" Journal of Clinical Medicine 10, no. 18: 4110. https://doi.org/10.3390/jcm10184110
APA StyleJeican, I. I., Gheban, D., Barbu-Tudoran, L., Inișca, P., Albu, C., Ilieș, M., Albu, S., Vică, M. L., Matei, H. V., Tripon, S., Lazăr, M., Aluaș, M., Siserman, C. V., Muntean, M., Trombitas, V., Iuga, C. A., Opincariu, I., & Junie, L. M. (2021). Respiratory Nasal Mucosa in Chronic Rhinosinusitis with Nasal Polyps versus COVID-19: Histopathology, Electron Microscopy Analysis and Assessing of Tissue Interleukin-33. Journal of Clinical Medicine, 10(18), 4110. https://doi.org/10.3390/jcm10184110