
Characteristics of the Urinary Proteome in Women with Overactive Bladder Syndrome: A Case-Control Study

 , ,
, ,  ,
,
Abstract
1. Introduction
2. Materials and Methods
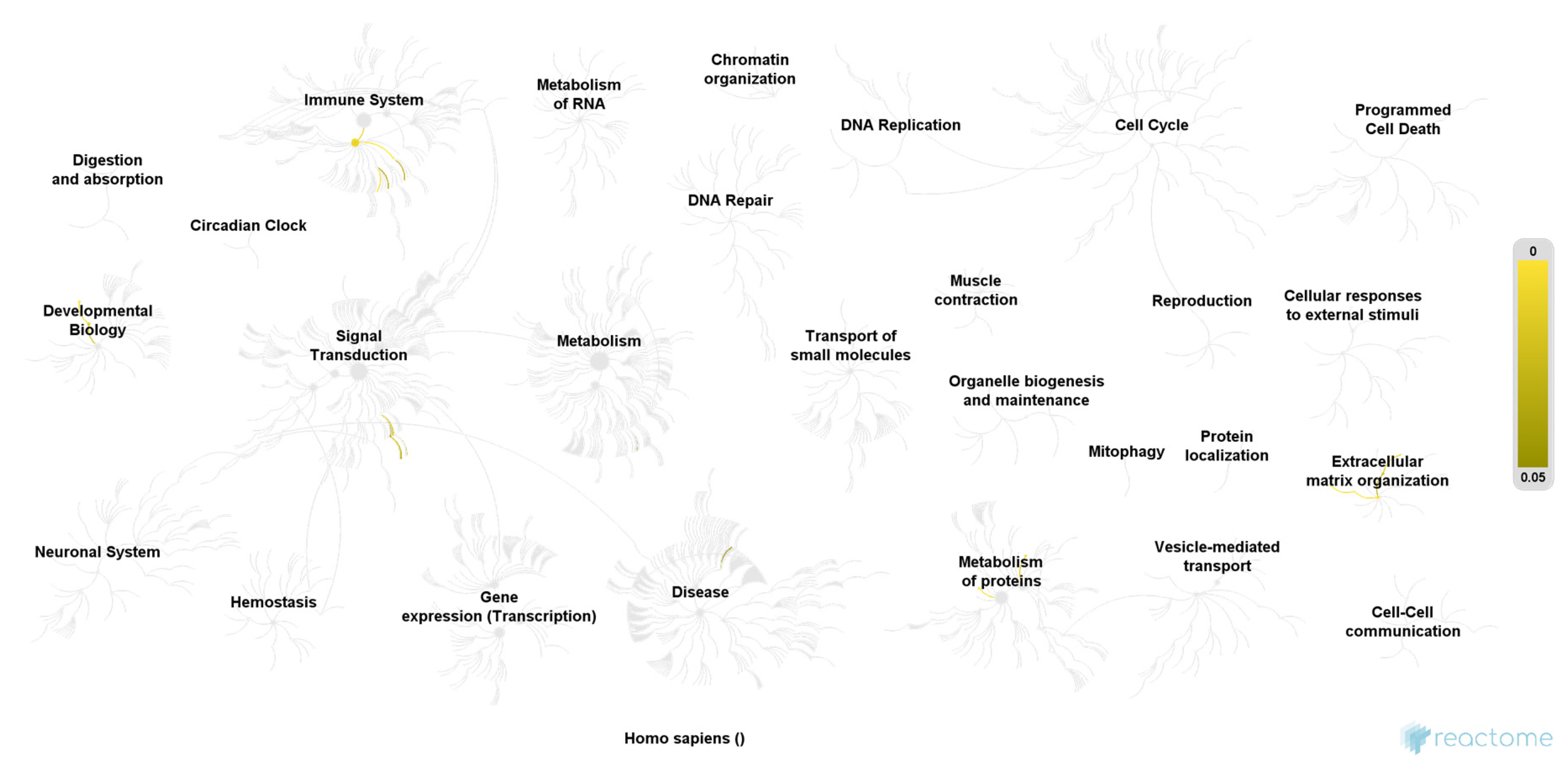
3. Results
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Haylen, B.T.; de Ridder, D.; Freeman, R.M.; Swift, S.E.; Berghmans, B.; Lee, J.; Monga, A.; Petri, E.; Rizk, D.E.; Sand, P.K.; et al. An International Urogynecological Association (IUGA)/International Continence Society (ICS) joint report on the terminology for female pelvic floor dysfunction. Int. Urogynecol. J. 2010, 21, 5–26. [Google Scholar] [CrossRef] [PubMed]
- Cheung, W.W.; Khan, N.H.; Choi, K.K.; Bluth, M.H.; Vincent, M.T. Prevalence, evaluation and management of overactive bladder in primary care. BMC Fam. Pract. 2009, 10, 8. [Google Scholar] [CrossRef]
- Melville, J.L.; Katon, W.; Delaney, K.; Newton, K. Urinary incontinence in US women: A population-based study. Arch. Intern. Med. 2005, 165, 537–542. [Google Scholar] [CrossRef]
- Irwin, D.E.; Mungapen, L.; Milsom, I.; Kopp, Z.; Reeves, P.; Kelleher, C. The economic impact of overactive bladder syndrome in six Western countries. BJU Int. 2009, 103, 202–209. [Google Scholar] [CrossRef]
- Corcos, J.; Przydacz, M.; Campeau, L.; Gray, G.; Hickling, D.; Honeine, C.; Radomski, S.B.; Stothers, L.; Wagg, A.; Lond, F. CUA guideline on adult overactive bladder. Can. Urol. Assoc. J. J. L’association Urol. Can. 2017, 11, E142–E173. [Google Scholar] [CrossRef]
- Cardozo, L.; Chapple, C.R.; Dmochowski, R.; Fitzgerald, M.P.; Hanno, P.; Michel, M.C.; Staskin, D.; Van Kerrebroeck, P.; Wyndaele, J.J.; Yamaguchi, O.; et al. Urinary urgency—Translating the evidence base into daily clinical practice. Int. J. Clin. Pract. 2009, 63, 1675–1682. [Google Scholar] [CrossRef]
- Bhide, A.A.; Cartwright, R.; Khullar, V.; Digesu, G.A. Biomarkers in overactive bladder. Int. Urogynecol. J. 2013, 24, 1065–1072. [Google Scholar] [CrossRef]
- Tyagi, P.; Tyagi, V.; Qu, X.; Lin, H.T.; Kuo, H.C.; Chuang, Y.C.; Chancellor, M. Association of inflammaging (inflammation + aging) with higher prevalence of OAB in elderly population. Int. Urol. Nephrol. 2013. [Google Scholar] [CrossRef]
- Liu, H.T.; Jiang, Y.H.; Kuo, H.C. Increased serum adipokines implicate chronic inflammation in the pathogenesis of overactive bladder syndrome refractory to antimuscarinic therapy. PLoS ONE 2013, 8, e76706. [Google Scholar] [CrossRef]
- Rachaneni, S.; Arya, P.; Latthe, P. Urinary nerve growth factor: A biomarker of detrusor overactivity? A systematic review. Int. Urogynecol. J. 2013, 24, 1603–1609. [Google Scholar] [CrossRef] [PubMed]
- Kashyap, M.; Kawamorita, N.; Tyagi, V.; Sugino, Y.; Chancellor, M.; Yoshimura, N.; Tyagi, P. Down-regulation of nerve growth factor expression in the bladder by antisense oligonucleotides as new treatment for overactive bladder. J. Urol. 2013, 190, 757–764. [Google Scholar] [CrossRef]
- Seth, J.H.; Sahai, A.; Khan, M.S.; van der Aa, F.; de Ridder, D.; Panicker, J.N.; Dasgupta, P.; Fowler, C.J. Nerve growth factor (NGF): A potential urinary biomarker for overactive bladder syndrome (OAB)? BJU Int. 2013, 111, 372–380. [Google Scholar] [CrossRef]
- Antunes-Lopes, T.; Pinto, R.; Barros, S.C.; Botelho, F.; Silva, C.M.; Cruz, C.D.; Cruz, F. Urinary neurotrophic factors in healthy individuals and patients with overactive bladder. J. Urol. 2013, 189, 359–365. [Google Scholar] [CrossRef]
- Cho, K.J.; Kim, H.S.; Koh, J.S.; Kim, J.C. Changes in urinary nerve growth factor and prostaglandin E2 in women with overactive bladder after anticholinergics. Int. Urogynecol. J. 2013, 24, 325–330. [Google Scholar] [CrossRef]
- Wang, L.W.; Han, X.M.; Chen, C.H.; Ma, Y.; Hai, B. Urinary brain-derived neurotrophic factor: A potential biomarker for objective diagnosis of overactive bladder. Int. Urol. Nephrol. 2013. [Google Scholar] [CrossRef]
- Koch, M.; Mitulovic, G.; Hanzal, E.; Umek, W.; Seyfert, S.; Mohr, T.; Koelbl, H.; Laterza, R.M. Urinary proteomic pattern in female stress urinary incontinence: A pilot study. Int. Urogynecol. J. 2016, 27, 1729–1734. [Google Scholar] [CrossRef][Green Version]
- Koch, M.; Umek, W.; Hanzal, E.; Mohr, T.; Seyfert, S.; Koelbl, H.; Mitulović, G. Serum proteomic pattern in female stress urinary incontinence. Electrophoresis 2018, 39, 1071–1078. [Google Scholar] [CrossRef]
- Futyma, K.; Nowakowski, Ł.; Ziętek-Strobl, A.; Kamińska, A.; Taoussi, N.; Rechberger, T. Urine Proteomic Study in OAB Patients-Preliminary Report. J. Clin. Med. 2020, 9, 1389. [Google Scholar] [CrossRef]
- Wessel, D.; Flügge, U.I. A method for the quantitative recovery of protein in dilute solution in the presence of detergents and lipids. Anal. Biochem. 1984, 138, 141–143. [Google Scholar] [CrossRef]
- Mitulović, G.; Stingl, C.; Steinmacher, I.; Hudecz, O.; Hutchins, J.R.; Peters, J.M.; Mechtler, K. Preventing carryover of peptides and proteins in nano LC-MS separations. Anal. Chem. 2009, 81, 5955–5960. [Google Scholar] [CrossRef]
- Jung, S.H. Sample size for FDR-control in microarray data analysis. Bioinformatics 2005, 21, 3097–3104. [Google Scholar] [CrossRef] [PubMed]
- Jassal, B.; Matthews, L.; Viteri, G.; Gong, C.; Lorente, P.; Fabregat, A.; Sidiropoulos, K.; Cook, J.; Gillespie, M.; Haw, R.; et al. The reactome pathway knowledgebase. Nucleic Acids Res. 2020, 48, D498–D503. [Google Scholar] [CrossRef]
- Fabregat, A.; Sidiropoulos, K.; Viteri, G.; Forner, O.; Marin-Garcia, P.; Arnau, V.; D’Eustachio, P.; Stein, L.; Hermjakob, H. Reactome pathway analysis: A high-performance in-memory approach. BMC Bioinform. 2017, 18, 142. [Google Scholar] [CrossRef]
- Kültz, D. Molecular and evolutionary basis of the cellular stress response. Annu. Rev. Physiol. 2005, 67, 225–257. [Google Scholar] [CrossRef]
- Yang, Y.; Yu, X. Regulation of apoptosis: The ubiquitous way. FASEB J. Off. Publ. Fed. Am. Soc. Exp. Biol. 2003, 17, 790–799. [Google Scholar] [CrossRef]
- Stanley, P.; Schachter, H.; Taniguchi, N. N-Glycans. In Essentials of Glycobiology; Varki, A., Cummings, R.D., Esko, J.D., Freeze, H.H., Stanley, P., Bertozzi, C.R., Hart, G.W., Etzler, M.E., Eds.; Cold Spring Harbor Laboratory Press: Cold Spring Harbor, NY, USA, 2009. [Google Scholar]
- Sparks, S.E.; Krasnewich, D.M. Congenital Disorders of N-Linked Glycosylation and Multiple Pathway Overview. In GeneReviews(®); Adam, M.P., Ardinger, H.H., Pagon, R.A., Wallace, S.E., Bean, L.J.H., Mirzaa, G., Amemiya, A., Eds.; University of Washington: Seattle, WA, USA, 1993. [Google Scholar]
- Depner, C.M.; Melanson, E.L.; McHill, A.W.; Wright, K.P., Jr. Mistimed food intake and sleep alters 24-hour time-of-day patterns of the human plasma proteome. Proc. Natl. Acad. Sci. USA 2018, 115, E5390–E5399. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
| OAB (N = 20) | Control (N = 20) | p-Value | |
|---|---|---|---|
| (Mean ± SD) | |||
| Age (years) | 57.5 (±16) | 55.3 (±17) | 0.904 |
| BMI (kg/m2) | 27.7 (±5) | 25.5 (±5) | 0.740 |
| Vaginal deliveries (n) | 1.15 | 0.99 | 0.484 |
| Chronic diseases * (n) | 12/20 (60%) | 8/20 (40%) | 0.206 |
| Menopausal status (n) | 0.744 | ||
| premenopausal | 7/20 (35%) | 8/20 (40%) | |
| postmenopausal | 13/20 (65%) | 12/20 (60%) | |
| Smoking (yes) | 2/20 (10%) | 3/20 (15%) | 0.633 |
| Caffeine consumption | 17/20 (85%) | 16/20 (80%) | 0.677 |
| ICIQ sum score | 31 | 0 | - |
| Protein | Accession | Gene Symbol |
|---|---|---|
| 26S proteasome non-ATPase regulatory subunit 11 | O00231 | PSD11 |
| 26S proteasome regulatory subunit 10B | A0A087 × 2I1 | A0A087 × 2I1 |
| 3-hydroxyacyl-CoA dehydrogenase type-2 | Q99714 | HCD2 |
| 60S ribosomal protein L15 | P61313 | RL15 |
| Alpha-centractin | R4GMT0 | R4GMT0 |
| Alpha-mannosidase 2 | Q16706 | MA2A1 |
| Asparagine--tRNA ligase, cytoplasmic | O43776 | SYNC |
| Beta/gamma crystallin domain-containing protein 1 | A0A0J9YWL0 | A0A0J9YWL0 |
| Bleomycin hydrolase | Q13867 | BLMH |
| Coatomer subunit beta | P53618 | COPB |
| Cytoplasmic dynein 1 heavy chain 1 | Q14204 | DYHC1 |
| Cytosolic phospholipase A2 delta | Q86XP0 | PA24D |
| Electron transfer flavoprotein subunit alpha, mitochondrial | P13804 | ETFA |
| Eukaryotic translation initiation factor 2 subunit 1 | P05198 | IF2A |
| Eukaryotic translation initiation factor 3 subunit A | Q14152 | EIF3A |
| Heat shock 70 kDa protein 4L | O95757 | HS74L |
| Isoform 2 of 26S proteasome regulatory subunit 6B | P43686-2 | PRS6B |
| Isoform 2 of Alpha-S1-casein | P47710-2 | CASA1 |
| Isoform 2 of Glutamine--tRNA ligase | P47897-2 | SYQ |
| Isoform 2 of Nck-associated protein 1 | Q9Y2A7-2 | NCKP1 |
| Isoform 2 of Serine/threonine-protein phosphatase 2A 55 kDa regulatory subunit B alpha isoform | P63151-2 | 2ABA |
| Isoform 2 of Threonine--tRNA ligase, cytoplasmic | P26639-2 | SYTC |
| Kappa-casein | P07498 | CASK |
| Mitogen-activated protein kinase 1 | P28482 | MK01 |
| Mucin-4 | Q99102 | MUC4 |
| Peroxisomal acyl-coenzyme A oxidase 1 | Q15067 | ACOX1 |
| Polypyrimidine tract-binding protein 1 | A0A0U1RRM4 | A0A0U1RRM4 |
| Prenylcysteine oxidase 1 | Q9UHG3 | PCYOX |
| Proteasome subunit alpha type-2 | P25787 | PSA2 |
| Protein flightless-1 homolog | Q13045 | FLII |
| Spectrin beta chain, non-erythrocytic 2 | O15020 | SPTN2 |
| Spliceosome RNA helicase DDX39B | Q13838 | DX39B |
| Tripartite motif-containing protein 16 | O95361 | TRI16 |
| Tropomodulin-3 | Q9NYL9 | TMOD3 |
| UDP-glucose 4-epimerase | Q14376 | GALE |
| Vesicle-fusing ATPase | P46459 | NSF |
| X-ray repair cross-complementing protein 5 | P13010 | XRCC5 |
| Protein | Accession | Gene Symbol |
|---|---|---|
| Trefoil factor 3 | X6R3S7 | X6R3S7 |
| Latent-transforming growth factor beta-binding protein 2 | G3V3 × 5 | G3V3 × 5 |
| Meprin A subunit | B7ZL91 | B7ZL91 |
| Testican-1 | A0A0A0MQX7 | A0A0A0MQX7 |
| IgGFc-binding protein | Q9Y6R7 | FCGBP |
| Frizzled-4 | Q9ULV1 | FZD4 |
| Mucin-5B | Q9HC84 | MUC5B |
| Growth/differentiation factor 15 | Q99988 | GDF15 |
| Isoform 2 of Phosphoinositide-3-kinase-interacting protein 1 | Q96FE7-2 | P3IP1 |
| BPI fold-containing family B member 1 | Q8TDL5 | BPIB1 |
| CD99 antigen-like protein 2 | Q8TCZ2 | C99L2 |
| Adhesion G protein-coupled receptor F5 | Q8IZF2 | AGRF5 |
| Protein S100-A7A | Q86SG5 | S1A7A |
| Elongation factor Tu | Q83ES6 | EFTU |
| Protocadherin Fat 4 | Q6V0I7 | FAT4 |
| Na(+)/H(+) exchange regulatory cofactor NHE-RF3 | Q5T2W1 | NHRF3 |
| FRAS1-related extracellular matrix protein 2 | Q5SZK8 | FREM2 |
| Trefoil factor 2 | Q03403 | TFF2 |
| Fibulin-2 | P98095 | FBLN2 |
| Hepcidin | P81172 | HEPC |
| Brain acid soluble protein 1 | P80723 | BASP1 |
| Hemoglobin subunit gamma-2 | P69892 | HBG2 |
| NPC intracellular cholesterol transporter 2 | P61916 | NPC2 |
| Beta-defensin 1 | P60022 | DEFB1 |
| Secreted Ly-6/uPAR-related protein 1 | P55000 | SLUR1 |
| Isoform 2 of Cysteine-rich secretory protein 3 | P54108-2 | CRIS3 |
| Afamin | P43652 | AFAM |
| Fibrillin-1 | P35555 | FBN1 |
| Isoform 2 of Syndecan-4 | P31431-2 | SDC4 |
| Granulins | P28799 | GRN |
| Serum paraoxonase/arylesterase 1 | P27169 | PON1 |
| CD27 antigen | P26842 | CD27 |
| Fibulin-1 | P23142 | FBLN1 |
| Small proline-rich protein 2D | P22532 | SPR2D |
| Cornifin-B | P22528 | SPR1B |
| Azurocidin | P20160 | CAP7 |
| Elafin | P19957 | ELAF |
| Inter-alpha-trypsin inhibitor heavy chain H2 | P19823 | ITIH2 |
| Complement component C7 | P10643 | CO7 |
| Matrilysin | P09237 | MMP7 |
| Tumor necrosis factor receptor superfamily member 16 | P08138 | TNR16 |
| Calbindin | P05937 | CALB1 |
| Heparin cofactor 2 | P05546 | HEP2 |
| Major prion protein | P04156 | PRIO |
| Trefoil factor 1 | P04155 | TFF1 |
| Antileukoproteinase | P03973 | SLPI |
| Cystatin-A | P01040 | CYTA |
| Complement C5 | P01031 | CO5 |
| Plasminogen | P00747 | PLMN |
| Superoxide dismutase [Cu-Zn] | P00441 | SODC |
| CD5 antigen-like | O43866 | CD5L |
| Ribonuclease T2 | O00584 | RNT2 |
| Isoform 2 of Deoxyribonuclease-2-alpha | O00115-2 | DNS2A |
| Pathway | Ratio of Identified Proteins | p-Value | False Discovery Rate |
|---|---|---|---|
| COPI-mediated anterograde transport | 5/102 | 0.00003 | 0.006 |
| Transport to the Golgi and subsequent modification | 6/186 | 0.00004 | 0.007 |
| ER to Golgi Anterograde Transport | 5/155 | 0.0002 | 0.012 |
| ABC-family proteins mediated transport | 4/103 | 0.0005 | 0.012 |
| Cellular responses to stress | 7/408 | 0.0005 | 0.012 |
| G2/M Transition | 5/198 | 0.0006 | 0.012 |
| Asparagine N-linked glycosylation | 6/305 | 0.0006 | 0.012 |
| Mitotic G2-G2/M phases | 5/200 | 0.0007 | 0.012 |
| Regulation of activated PAK-2p34 by proteasome mediated degradation | 3/50 | 0.0007 | 0.012 |
| Cross-presentation of soluble exogenous antigens (endosomes) | 3/50 | 0.0007 | 0.012 |
| Intra-Golgi and retrograde Golgi-to-ER traffic | 5/206 | 0.0007 | 0.012 |
| Regulation of ornithine decarboxylase (ODC) | 3/51 | 0.0008 | 0.012 |
| Autodegradation of the E3 ubiquitin ligase COP1 | 3/52 | 0.0008 | 0.012 |
| Ubiquitin-dependent degradation of Cyclin D1 | 3/52 | 0.0008 | 0.012 |
| Ubiquitin-dependent degradation of Cyclin D | 3/52 | 0.0008 | 0.012 |
| Transcriptional regulation by RUNX2 | 4/121 | 0.0009 | 0.012 |
| Vpu mediated degradation of CD4 | 3/53 | 0.0009 | 0.012 |
| Regulation of Apoptosis | 3/53 | 0.0009 | 0.012 |
| p53-Independent DNA Damage Response | 3/53 | 0.0009 | 0.012 |
| p53-Independent G1/S DNA damage checkpoint | 3/53 | 0.0009 | 0.012 |
| Ubiquitin Mediated Degradation of Phosphorylated Cdc25A | 3/53 | 0.0009 | 0.012 |
| FBXL7 down-regulates AURKA during mitotic entry and in early mitosis | 3/55 | 0.0010 | 0.012 |
| SCF-beta-TrCP mediated degradation of Emi1 | 3/55 | 0.0010 | 0.012 |
| Degradation of AXIN | 3/55 | 0.0010 | 0.012 |
| Negative regulation of NOTCH4 signaling | 3/55 | 0.0010 | 0.012 |
| Pathway | Ratio of Identified Proteins | p-Value | False Discovery Rate |
|---|---|---|---|
| Molecules associated with elastic fibers | 4/38 | 0.00004 | 0.004 |
| Elastic fiber formation | 4/45 | 0.00007 | 0.004 |
| Terminal pathway of complement | 2/8 | 0.0007 | 0.026 |
| Antimicrobial peptides | 4/95 | 0.001 | 0.033 |
| Regulation of Insulin-like Growth Factor (IGF) transport and uptake by Insulin-like Growth Factor Binding Proteins (IGFBPs) | 4/124 | 0.003 | 0.054 |
| Extracellular matrix organization | 6/301 | 0.003 | 0.054 |
| Formation of the cornified envelope | 4/129 | 0.003 | 0.054 |
| Innate Immune System | 12/1183 | 0.009 | 0.122 |
| Activation of Matrix Metalloproteinases | 2/33 | 0.011 | 0.132 |
| NFG and proNGF binds to p75NTR | 1/3 | 0.014 | 0.132 |
| Ceramide signaling | 1/3 | 0.014 | 0.132 |
| Post-translational protein phosphorylation | 3/107 | 0.015 | 0.132 |
| Axonal growth stimulation | 1/4 | 0.019 | 0.151 |
| Keratinization | 4/217 | 0.020 | 0.160 |
| NADE modulates death signaling | 1/6 | 0.028 | 0.169 |
| p75NTR negatively regulates cell cycle via SC1 | 1/6 | 0.028 | 0.169 |
| Metal sequestration by antimicrobial proteins | 1/6 | 0.028 | 0.169 |
| Degradation of the extracellular matrix | 3/140 | 0.029 | 0.176 |
| Activation of C3 and C5 | 1/7 | 0.033 | 0.187 |
| RNF mutants show enhanced WNT signaling and proliferation | 1/8 | 0.037 | 0.187 |
| Axonal growth inhibition (RHOA activation) | 1/9 | 0.042 | 0.205 |
| Synthesis of 5-eicosatetraenoic acids | 1/9 | 0.042 | 0.205 |
| p75NTR regulates axonogenesis | 1/10 | 0.047 | 0.205 |
| Regulated proteolysis of p75NTR | 1/11 | 0.051 | 0.205 |
| Dissolution of Fibrin Clot | 1/13 | 0.06 | 0.224 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Koch, M.; Lyatoshinsky, P.; Mitulovic, G.; Bodner-Adler, B.; Lange, S.; Hanzal, E.; Umek, W. Characteristics of the Urinary Proteome in Women with Overactive Bladder Syndrome: A Case-Control Study. J. Clin. Med. 2021, 10, 2446. https://doi.org/10.3390/jcm10112446
Koch M, Lyatoshinsky P, Mitulovic G, Bodner-Adler B, Lange S, Hanzal E, Umek W. Characteristics of the Urinary Proteome in Women with Overactive Bladder Syndrome: A Case-Control Study. Journal of Clinical Medicine. 2021; 10(11):2446. https://doi.org/10.3390/jcm10112446
Chicago/Turabian StyleKoch, Marianne, Pavel Lyatoshinsky, Goran Mitulovic, Barbara Bodner-Adler, Sören Lange, Engelbert Hanzal, and Wolfgang Umek. 2021. "Characteristics of the Urinary Proteome in Women with Overactive Bladder Syndrome: A Case-Control Study" Journal of Clinical Medicine 10, no. 11: 2446. https://doi.org/10.3390/jcm10112446
APA StyleKoch, M., Lyatoshinsky, P., Mitulovic, G., Bodner-Adler, B., Lange, S., Hanzal, E., & Umek, W. (2021). Characteristics of the Urinary Proteome in Women with Overactive Bladder Syndrome: A Case-Control Study. Journal of Clinical Medicine, 10(11), 2446. https://doi.org/10.3390/jcm10112446