
Recombinant Toxoplasma gondii Ribosomal Protein P2 Modulates the Functions of Murine Macrophages In Vitro and Provides Immunity against Acute Toxoplasmosis In Vivo





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Materials and Methods
2.1. Animals, Parasites, and Cultured Cells
2.2. Cloning and Molecular Characterization of TgRPP2
2.3. Expression and Purification of Recombinant TgRPP2 and the pET32a Vector Protein
2.4. Total Soluble Protein of T. gondii Tachyzoites
2.5. Western Blot Analysis of the pET32a Vector Protein, Recombinant, and Native TgRPP2
2.6. Confirmation of rTgRPP2 Binding with Murine Macrophages
2.7. Cell Proliferation Detection Assay
2.8. Detection of Cell Apoptosis
2.9. FITC-dextran Internalization Assay
2.10. Nitric oxide (NO) Production Assay
2.11. Identification of the Cytokine Level
2.12. Animal Vaccination and Challenge
2.13. Determination of the Levels of Antibodies and Cytokines
2.14. Major Histocompatibility Complex (MHC) Molecule Analysis
2.15. Statistical Analysis
3. Results
3.1. Preparation of the Eukaryotic Expression Plasmid
3.2. Expression, Purification, and Western Blot Analysis of Recombinant TgRPP2 and the pET32a Vector Protein
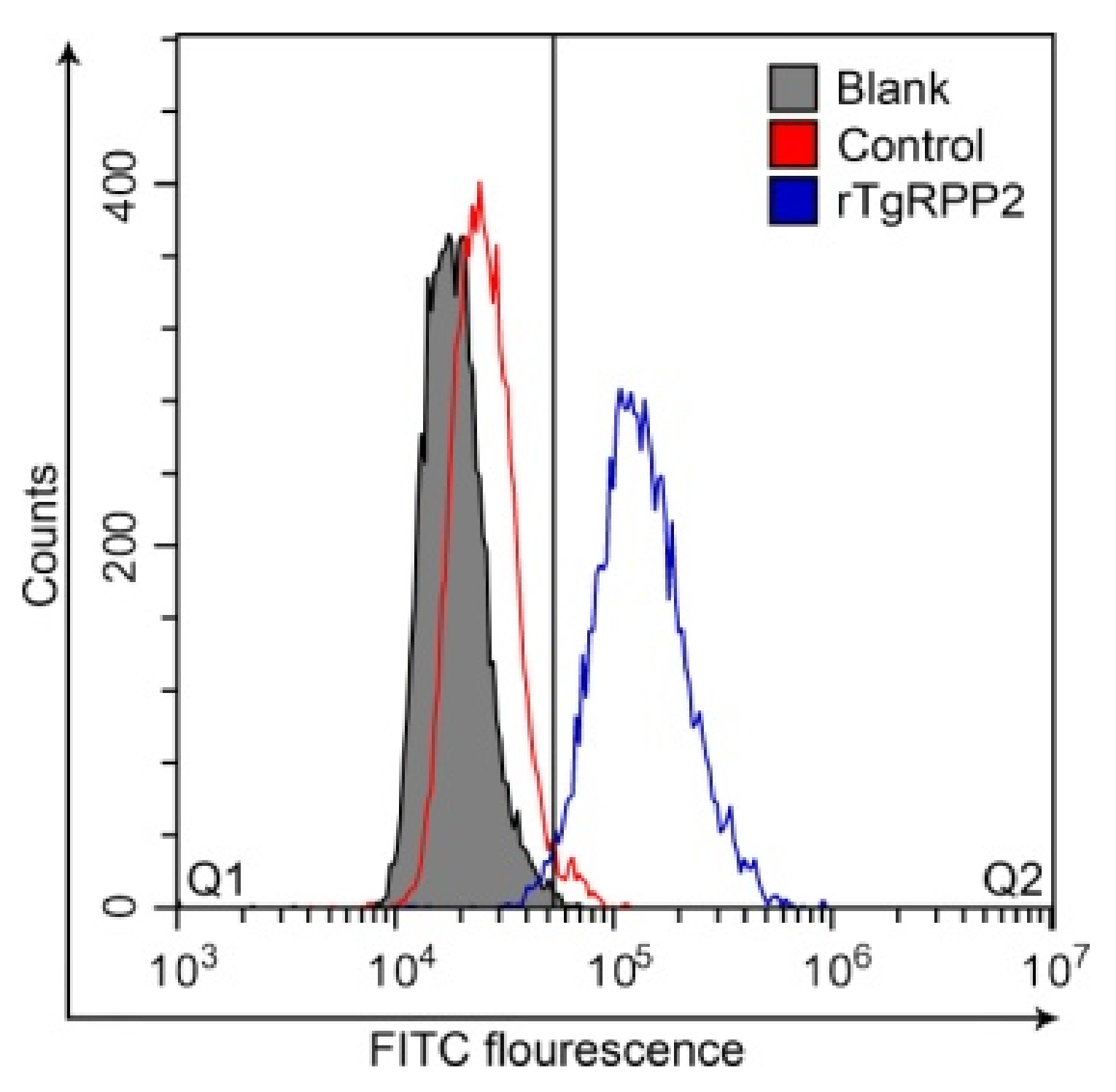
3.3. Validation of the Binding Capability of Recombinant TgRPP2 with Murine Macrophages
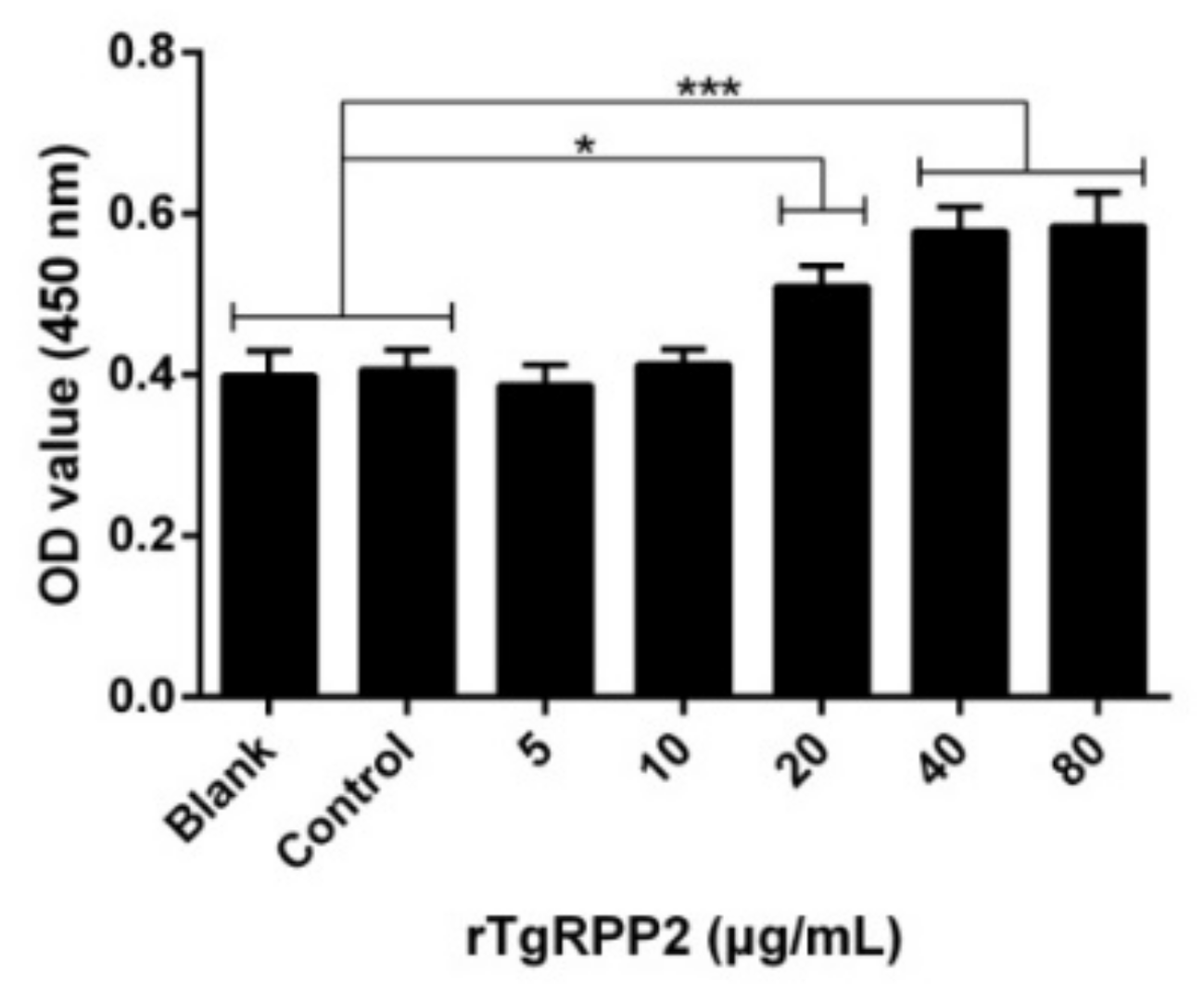
3.4. Enhanced Proliferation of Murine Macrophages Triggered by Recombinant TgRPP2
3.5. Promoted Apoptosis of Ana-1 Cells Induced by Recombinant TgRPP2
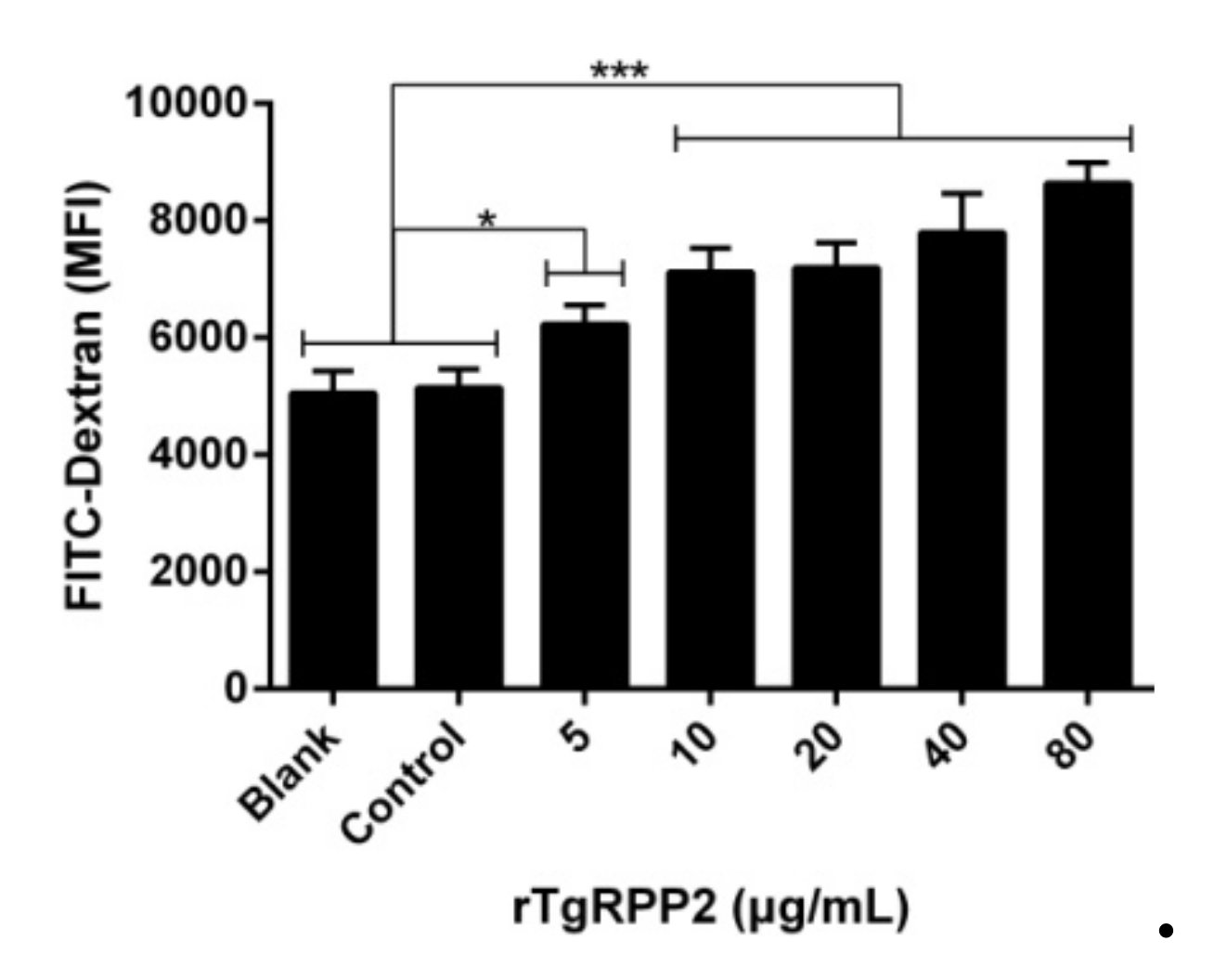
3.6. Promoted Phagocytosis in Ana-1 Cells Induced by Recombinant TgRPP2
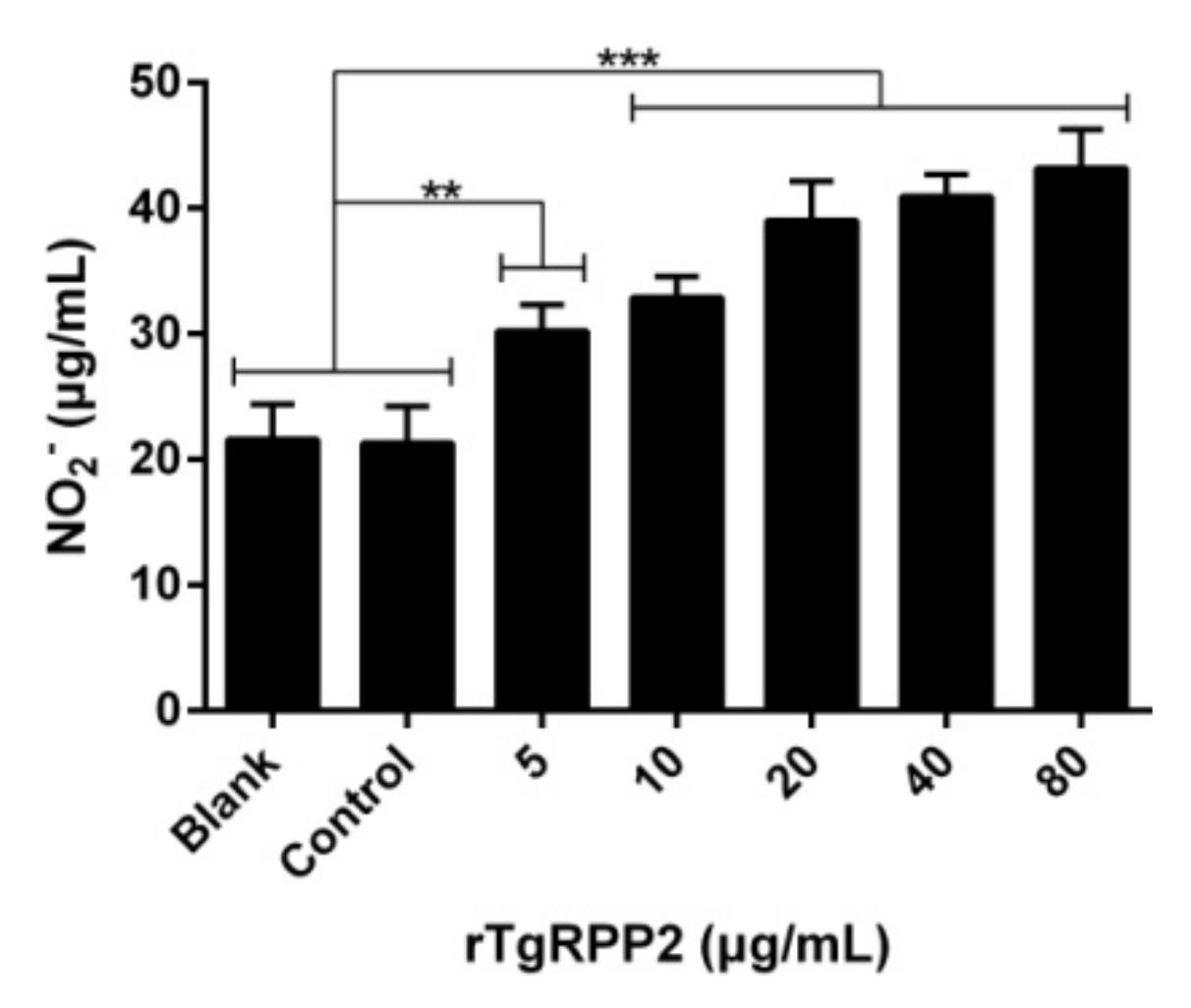
3.7. Enhanced NO Secretion of Murine Macrophages Triggered by Recombinant TgRPP2
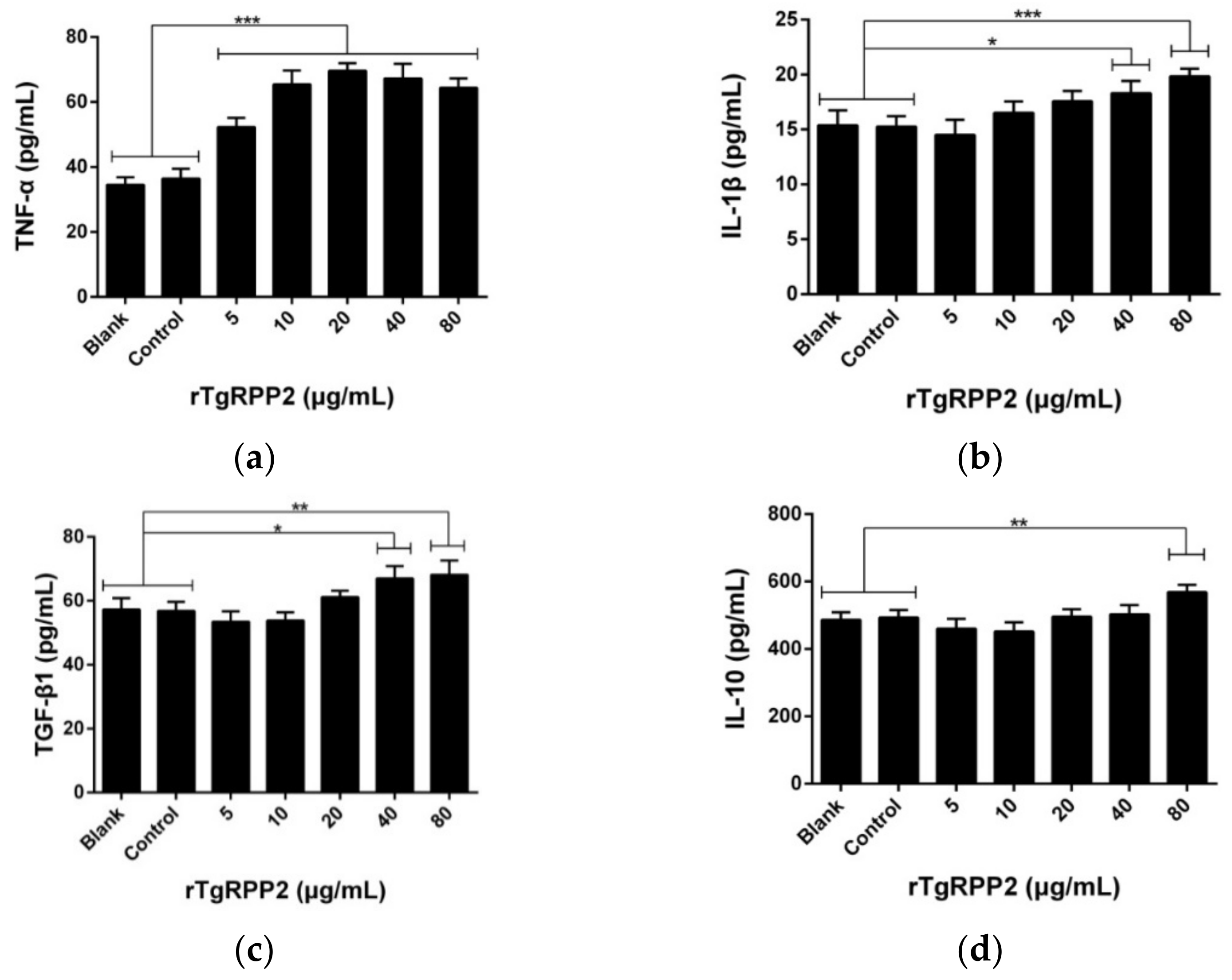
3.8. Modulation Effects in the Cytokine Secretion of Murine Macrophages
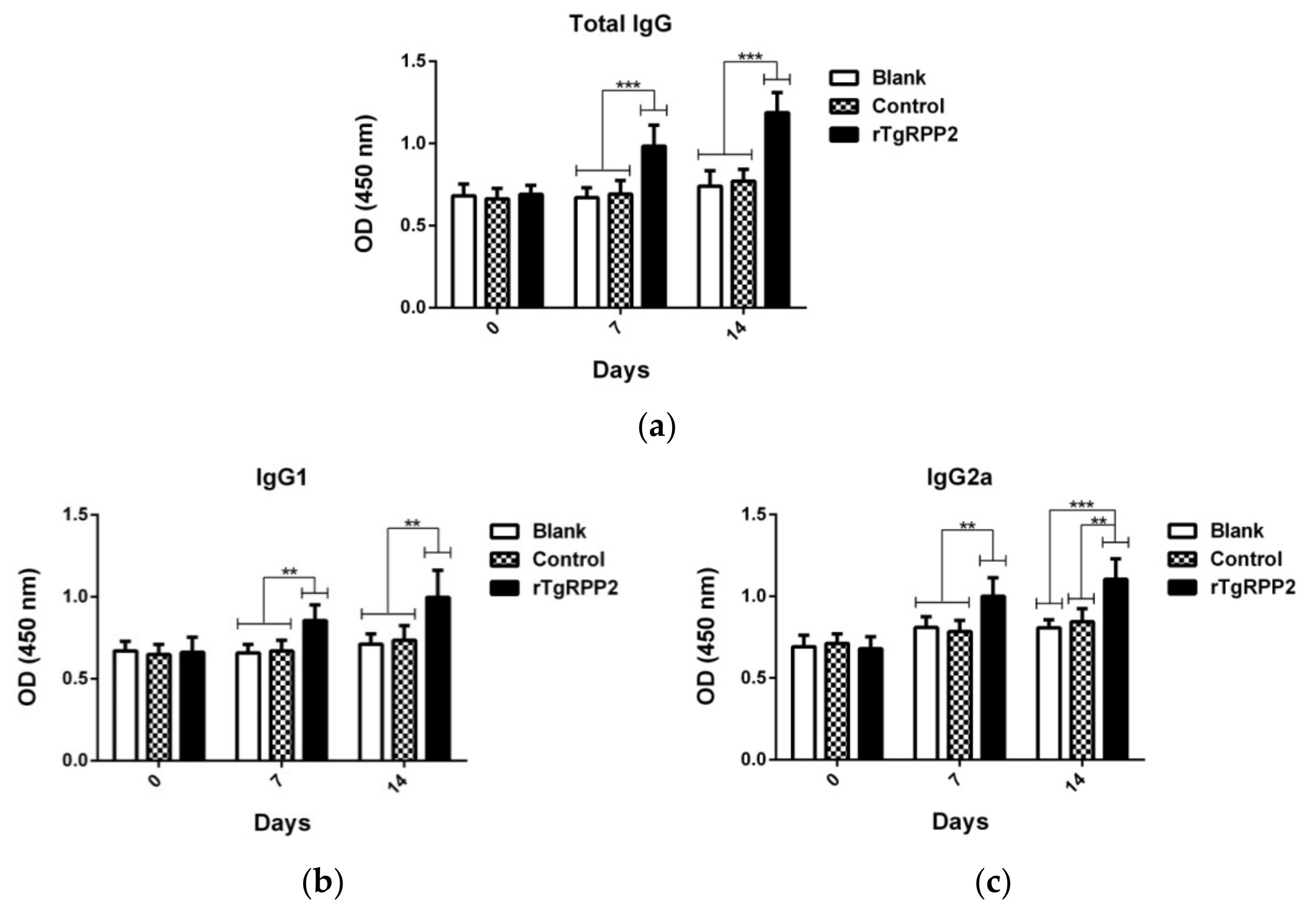
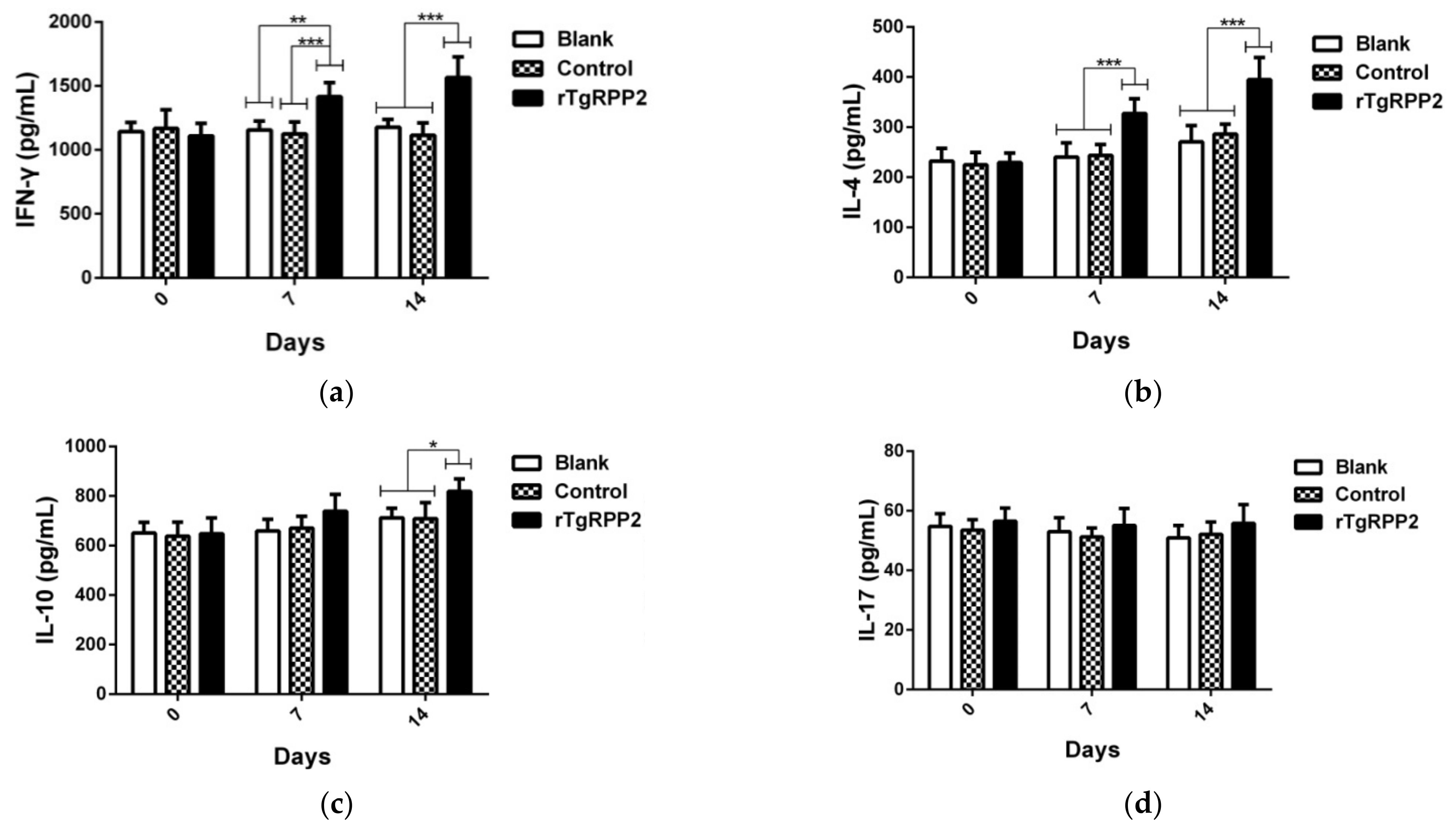
3.9. Modulation Effects in Antibodies and Cytokine Secretion In Vivo
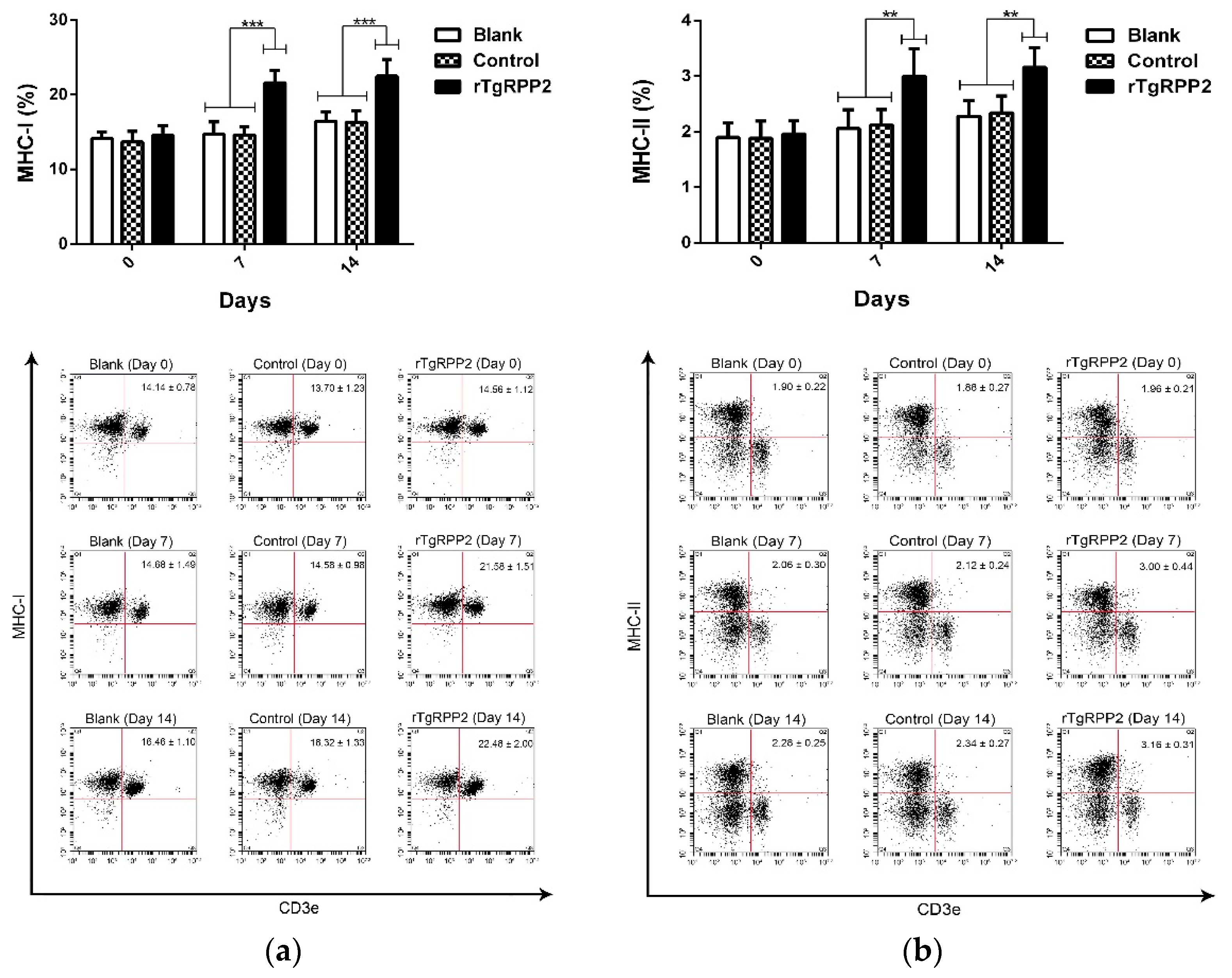
3.10. The Major Histocompatibility Complex Molecule Changes in Murine Spleen Lymphocytes
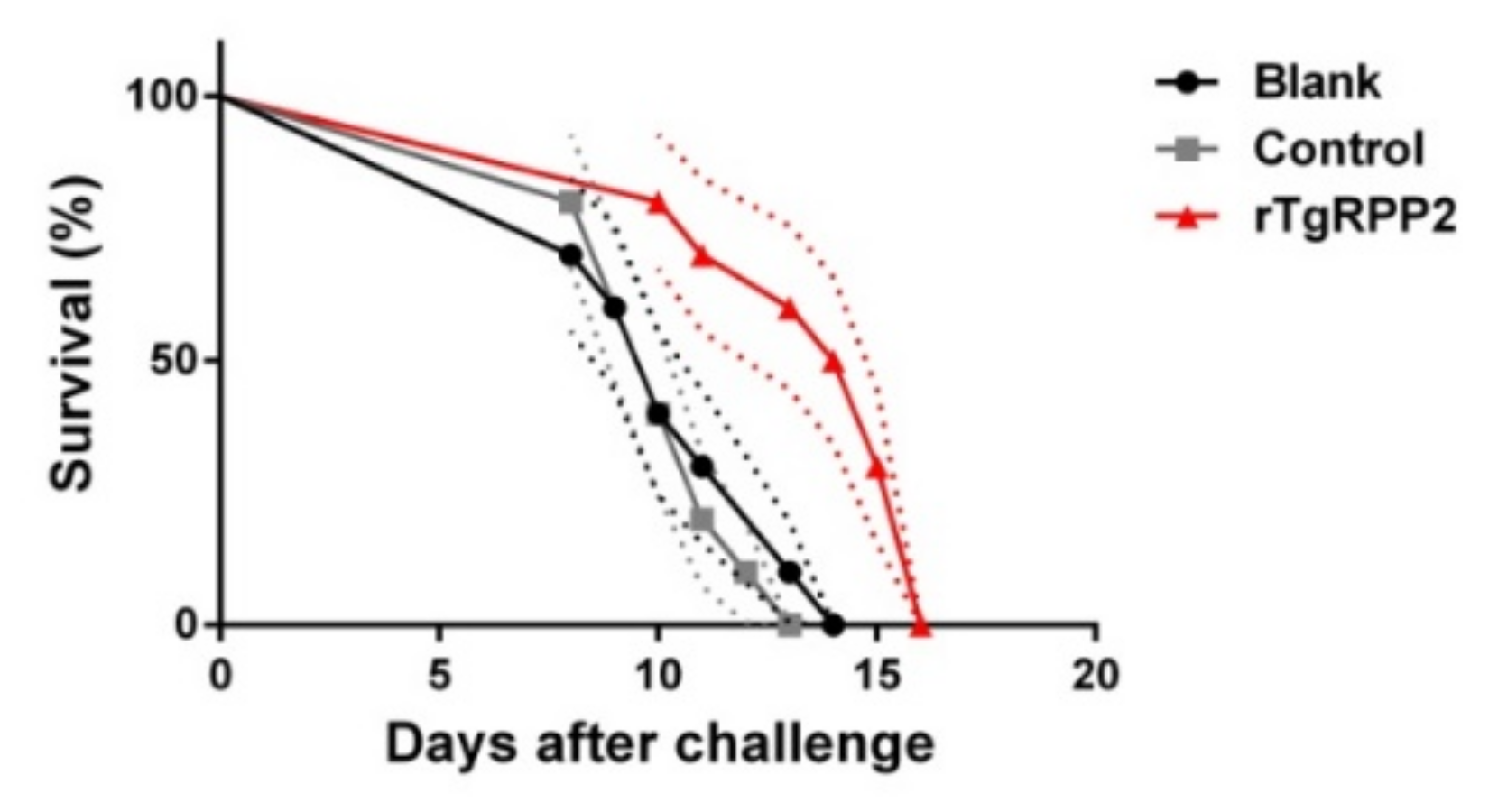
3.11. Immune Protection of Vaccinated Mice against Acute T. gondii Infections
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Zheng, J.; Cheng, Z.; Jia, H.; Zheng, Y. Characterization of aspartyl aminopeptidase from Toxoplasma gondii. Sci. Rep. 2016, 6, 34448. [Google Scholar] [CrossRef]
- Dubey, J.P. History of the discovery of the life cycle of Toxoplasma gondii. Int. J. Parasitol. 2009, 39, 877–882. [Google Scholar] [CrossRef] [PubMed]
- Montoya, J.G.; Liesenfeld, O. Toxoplasmosis. Lancet 2004, 363, 1965–1976. [Google Scholar] [CrossRef]
- Jeffers, V.; Tampaki, Z.; Kim, K.; Sullivan, W.J., Jr. A latent ability to persist: Differentiation in Toxoplasma gondii. Cell Mol. Life Sci. 2018, 75, 2355–2373. [Google Scholar] [CrossRef] [PubMed]
- Ahmadpour, E.; Daryani, A.; Sharif, M.; Sarvi, S.; Aarabi, M.; Mizani, A.; Rahimi, M.T.; Shokri, A. Toxoplasmosis in immunocompromised patients in Iran: A systematic review and meta-analysis. J. Infect. Dev. Countr. 2014, 8, 1503–1510. [Google Scholar] [CrossRef]
- Sroka, J.; Bilska-Zajac, E.; Wojcik-Fatla, A.; Zajac, V.; Dutkiewicz, J.; Karamon, J.; Piotrowska, W.; Cencek, T. Detection and Molecular Characteristics of Toxoplasma gondii DNA in Retail Raw Meat Products in Poland. Foodborne Pathog. Dis. 2019, 16, 195–204. [Google Scholar] [CrossRef]
- Marques, C.S.; Sousa, S.; Castro, A.; da Costa, J.M.C. Detection of Toxoplasma gondii oocysts in fresh vegetables and berry fruits. Parasit. Vectors 2020, 13, 180. [Google Scholar] [CrossRef] [PubMed]
- Hernandez-Cortazar, I.B.; Acosta-Viana, K.Y.; Guzman-Marin, E.; Ortega-Pacheco, A.; Segura-Correa, J.C.; Jimenez-Coello, M. Presence of Toxoplasma gondii in Drinking Water from an Endemic Region in Southern Mexico. Foodborne Pathog. Dis. 2017, 14, 288–292. [Google Scholar] [CrossRef]
- Khan, K.; Khan, W. Congenital toxoplasmosis: An overview of the neurological and ocular manifestations. Parasitol. Int. 2018, 67, 715–721. [Google Scholar] [CrossRef]
- Olariu, T.R.; Remington, J.S.; McLeod, R.; Alam, A.; Montoya, J.G. Severe congenital toxoplasmosis in the United States: Clinical and serologic findings in untreated infants. Pediatr. Infect. Dis. J. 2011, 30, 1056–1061. [Google Scholar] [CrossRef]
- Fallahi, S.; Rostami, A.; Nourollahpour Shiadeh, M.; Behniafar, H.; Paktinat, S. An updated literature review on maternal-fetal and reproductive disorders of Toxoplasma gondii infection. J. Gynecol. Obstet. Hum. Reprod. 2018, 47, 133–140. [Google Scholar] [CrossRef]
- Brown, A.S.; Schaefer, C.A.; Quesenberry, C.P., Jr.; Liu, L.; Babulas, V.P.; Susser, E.S. Maternal exposure to toxoplasmosis and risk of schizophrenia in adult offspring. Am. J. Psychiatry 2005, 162, 767–773. [Google Scholar] [CrossRef] [PubMed]
- Buxton, D.; Innes, E.A. A commercial vaccine for ovine toxoplasmosis. Parasitology 1995, 110, S11–S16. [Google Scholar] [CrossRef]
- Dittmar, A.J.; Drozda, A.A.; Blader, I.J. Drug Repurposing Screening Identifies Novel Compounds That Effectively Inhibit Toxoplasma gondii Growth. Msphere 2016, 1, e00042-15. [Google Scholar] [CrossRef]
- Lyons, R.E.; McLeod, R.; Roberts, C.W. Toxoplasma gondii tachyzoite-bradyzoite interconversion. Trends Parasitol. 2002, 18, 198–201. [Google Scholar] [CrossRef]
- Ben-Harari, R.R.; Goodwin, E.; Casoy, J. Adverse Event Profile of Pyrimethamine-Based Therapy in Toxoplasmosis: A Systematic Review. Drugs R D 2017, 17, 523–544. [Google Scholar] [CrossRef] [PubMed]
- Mandelbrot, L.; Kieffer, F.; Sitta, R.; Laurichesse-Delmas, H.; Winer, N.; Mesnard, L.; Berrebi, A.; Le Bouar, G.; Bory, J.P.; Cordier, A.G.; et al. Prenatal therapy with pyrimethamine + sulfadiazine vs spiramycin to reduce placental transmission of toxoplasmosis: A multicenter, randomized trial. Am. J. Obstet. Gynecol. 2018, 219, 386.e1–386.e9. [Google Scholar] [CrossRef] [PubMed]
- Sasai, M.; Pradipta, A.; Yamamoto, M. Host immune responses to Toxoplasma gondii. Int. Immunol. 2018, 30, 113–119. [Google Scholar] [CrossRef]
- Liu, Y.C.; Zou, X.B.; Chai, Y.F.; Yao, Y.M. Macrophage polarization in inflammatory diseases. Int. J. Biol. Sci. 2014, 10, 520–529. [Google Scholar] [CrossRef] [PubMed]
- Labonte, A.C.; Tosello-Trampont, A.C.; Hahn, Y.S. The role of macrophage polarization in infectious and inflammatory diseases. Mol Cells 2014, 37, 275–285. [Google Scholar] [CrossRef]
- Gordon, S.; Taylor, P.R. Monocyte and macrophage heterogeneity. Nat. Rev. Immunol. 2005, 5, 953–964. [Google Scholar] [CrossRef]
- Gordon, S. Alternative activation of macrophages. Nat. Rev. Immunol. 2003, 3, 23–35. [Google Scholar] [CrossRef] [PubMed]
- Liu, X.; Li, J.; Peng, X.; Lv, B.; Wang, P.; Zhao, X.; Yu, B. Geraniin Inhibits LPS-Induced THP-1 Macrophages Switching to M1 Phenotype via SOCS1/NF-κB Pathway. Inflammation 2016, 39, 1421–1433. [Google Scholar] [CrossRef]
- Ndao, O.; Puech, P.H.; Berard, C.; Limozin, L.; Rabhi, S.; Azas, N.; Dubey, J.P.; Dumetre, A. Dynamics of Toxoplasma gondii Oocyst Phagocytosis by Macrophages. Front. Cell Infect. Microbiol. 2020, 10, 207. [Google Scholar] [CrossRef] [PubMed]
- Park, J.; Hunter, C.A. The role of macrophages in protective and pathological responses to Toxoplasma gondii. Parasite Immunol. 2020, 42, e12712. [Google Scholar] [CrossRef] [PubMed]
- Barragan, A.; Sibley, L.D. Transepithelial migration of Toxoplasma gondii is linked to parasite motility and virulence. J. Exp. Med. 2002, 195, 1625–1633. [Google Scholar] [CrossRef] [PubMed]
- Lambert, H.; Hitziger, N.; Dellacasa, I.; Svensson, M.; Barragan, A. Induction of dendritic cell migration upon Toxoplasma gondii infection potentiates parasite dissemination. Cell Microbiol. 2006, 8, 1611–1623. [Google Scholar] [CrossRef] [PubMed]
- Tchórzewski, M. The acidic ribosomal P proteins. Int. J. Biochem. Cell Biol. 2002, 34, 911–915. [Google Scholar] [CrossRef]
- Lacombe, A.; Maclean, A.E.; Ovciarikova, J.; Tottey, J.; Muhleip, A.; Fernandes, P.; Sheiner, L. Identification of the Toxoplasma gondii mitochondrial ribosome, and characterisation of a protein essential for mitochondrial translation. Mol. Microbiol. 2019, 112, 1235–1252. [Google Scholar] [CrossRef]
- Nilsen, A.; LaCrue, A.N.; White, K.L.; Forquer, I.P.; Cross, R.M.; Marfurt, J.; Mather, M.W.; Delves, M.J.; Shackleford, D.M.; Saenz, F.E.; et al. Quinolone-3-diarylethers: A new class of antimalarial drug. Sci. Transl. Med. 2013, 5, 177ra137. [Google Scholar] [CrossRef]
- Phillips, M.A.; Lotharius, J.; Marsh, K.; White, J.; Dayan, A.; White, K.L.; Njoroge, J.W.; El Mazouni, F.; Lao, Y.; Kokkonda, S.; et al. A long-duration dihydroorotate dehydrogenase inhibitor (DSM265) for prevention and treatment of malaria. Sci. Transl. Med. 2015, 7, 296ra111. [Google Scholar] [CrossRef] [PubMed]
- Ke, H.; Mather, M.W. +Targeting Mitochondrial Functions as Antimalarial Regime, What Is Next? Curr. Clin. Micro Rpt. 2017, 4, 175–191. [Google Scholar] [CrossRef]
- Sudarsan, R.; Chopra, R.K.; Khan, M.A.; Sharma, S. Ribosomal protein P2 localizes to the parasite zoite-surface and is a target for invasion inhibitory antibodies in Toxoplasma gondii and Plasmodium falciparum. Parasitol. Int. 2015, 64, 43–49. [Google Scholar] [CrossRef] [PubMed]
- Szuster-Ciesielska, A.; Wawiorka, L.; Krokowski, D.; Grankowski, N.; Jarosz, L.; Lisiecka, U.; Tchorzewski, M. Immunogenic Evaluation of Ribosomal P-Protein Antigen P0, P1, and P2 and Pentameric Protein Complex P0-(P1-P2)2 of Plasmodium falciparum in a Mouse Model. J. Immunol. Res. 2019, 2019, 9264217. [Google Scholar] [CrossRef] [PubMed]
- Das, S.; Basu, H.; Korde, R.; Tewari, R.; Sharma, S. Arrest of nuclear division in Plasmodium through blockage of erythrocyte surface exposed ribosomal protein P2. PLoS Pathog. 2012, 8, e1002858. [Google Scholar] [CrossRef]
- Mishra, P.; Dmello, C.; Sengupta, D.; Chandrabhan Singh, S.; Kirkise, N.; Hosur, R.V.; Sharma, S. Molecular study of binding of Plasmodium ribosomal protein P2 to erythrocytes. Biochimie 2020, 176, 181–191. [Google Scholar] [CrossRef]
- Lamarque, M.; Besteiro, S.; Papoin, J.; Roques, M.; Vulliez-Le Normand, B.; Morlon-Guyot, J.; Dubremetz, J.F.; Fauquenoy, S.; Tomavo, S.; Faber, B.W.; et al. The RON2-AMA1 interaction is a critical step in moving junction-dependent invasion by apicomplexan parasites. PLoS Pathog. 2011, 7, e1001276. [Google Scholar] [CrossRef] [PubMed]
- Guang-Wei, Z.; Bo, S.; Qing, X.; Li-Xin, X.U.; Ruo-Feng, Y.; Xiao-Kai, S.; Adam, H.I.; Xiang-Rui, L.I. Isolation and Molecular Characterization of Toxoplasma gondii from Chickens in China. J. Integr. Agric. 2012, 11, 1347–1353. [Google Scholar] [CrossRef]
- Hassan, I.A.; Wang, S.; Xu, L.; Yan, R.; Song, X.; Li, X. Immunoglobulin and cytokine changes induced following immunization with a DNA vaccine encoding Toxoplasma gondii selenium-dependent glutathione reductase protein. Exp. Parasitol. 2014, 146, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Song, H.; Yan, R.; Xu, L.; Song, X.; Shah, M.A.; Zhu, H.; Li, X. Efficacy of DNA vaccines carrying Eimeria acervulina lactate dehydrogenase antigen gene against coccidiosis. Exp. Parasitol. 2010, 126, 224–231. [Google Scholar] [CrossRef] [PubMed]
- Green, L.C.; Wagner, D.A.; Glogowski, J.; Skipper, P.L.; Wishnok, J.S.; Tannenbaum, S.R. Analysis of nitrate, nitrite, and [15N]nitrate in biological fluids. Anal. Biochem. 1982, 126, 131–138. [Google Scholar] [CrossRef]
- Tang, X.M.; Yin, G.W.; Qin, M.; Tao, G.R.; Suo, J.X.; Liu, X.Y.; Suo, X. Transgenic Eimeria tenella as a vaccine vehicle: Expressing TgSAG1 elicits protective immunity against Toxoplasma gondii infections in chickens and mice. Sci. Rep.-Uk 2016, 6, 29379. [Google Scholar] [CrossRef] [PubMed]
- Di Carlo, F.J.; Fiore, J.V. On the composition of zymosan. Science 1958, 127, 756–757. [Google Scholar] [CrossRef] [PubMed]
- Mosser, D.M.; Edwards, J.P. Exploring the full spectrum of macrophage activation. Nat. Rev. Immunol. 2008, 8, 958–969. [Google Scholar] [CrossRef]
- Mantovani, A.; Biswas, S.K.; Galdiero, M.R.; Sica, A.; Locati, M. Macrophage plasticity and polarization in tissue repair and remodelling. J. Pathol. 2013, 229, 176–185. [Google Scholar] [CrossRef] [PubMed]
- Brunet, J.; Pfaff, A.W.; Abidi, A.; Unoki, M.; Nakamura, Y.; Guinard, M.; Klein, J.P.; Candolfi, E.; Mousli, M. Toxoplasma gondii exploits UHRF1 and induces host cell cycle arrest at G2 to enable its proliferation. Cell Microbiol. 2008, 10, 908–920. [Google Scholar] [CrossRef] [PubMed]
- Lavine, M.D.; Arrizabalaga, G. Induction of mitotic S-phase of host and neighboring cells by Toxoplasma gondii enhances parasite invasion. Mol. Biochem. Parasitol. 2009, 164, 95–99. [Google Scholar] [CrossRef] [PubMed]
- Kim, M.J.; Jung, B.K.; Cho, J.; Song, H.; Pyo, K.H.; Lee, J.M.; Kim, M.K.; Chai, J.Y. Exosomes Secreted by Toxoplasma gondii-Infected L6 Cells: Their Effects on Host Cell Proliferation and Cell Cycle Changes. Korean J. Parasitol. 2016, 54, 147–154. [Google Scholar] [CrossRef] [PubMed]
- Liu, X.C.; Li, X.Y.; Wang, Q.Q.; Sun, X.N.; Lu, M.M.; Ehsan, M.; Xu, L.X.; Yan, R.F.; Song, X.K.; Li, X.R. Toxoplasma gondii Histone 4 Affects Some Functions of Murine Ana-1 Macrophages In Vitro. J. Eukaryot. Microbiol. 2018, 65, 860–869. [Google Scholar] [CrossRef] [PubMed]
- Mammari, N.; Halabi, M.A.; Yaacoub, S.; Chlala, H.; Dardé, M.L.; Courtioux, B. Toxoplasma gondii Modulates the Host Cell Responses: An Overview of Apoptosis Pathways. Biomed. Res. Int. 2019, 2019, 6152489. [Google Scholar] [CrossRef]
- D’Arcy, M.S. Cell death: A review of the major forms of apoptosis, necrosis and autophagy. Cell Biol Int 2019, 43, 582–592. [Google Scholar] [CrossRef] [PubMed]
- Xu, X.; Lai, Y.; Hua, Z.C. Apoptosis and apoptotic body: Disease message and therapeutic target potentials. Biosci. Rep. 2019, 39, BSR20180992. [Google Scholar] [CrossRef]
- Payne, T.M.; Molestina, R.E.; Sinai, A.P. Inhibition of caspase activation and a requirement for NF-κB function in the Toxoplasma gondii-mediated blockade of host apoptosis. J. Cell. Sci. 2003, 116, 4345–4358. [Google Scholar] [CrossRef] [PubMed]
- Goebel, S.; Luder, C.G.; Gross, U. Invasion by Toxoplasma gondii protects human-derived HL-60 cells from actinomycin D-induced apoptosis. Med. Microbiol. Immunol. 1999, 187, 221–226. [Google Scholar] [CrossRef] [PubMed]
- Medina, C.B.; Mehrotra, P.; Arandjelovic, S.; Perry, J.S.A.; Guo, Y.; Morioka, S.; Barron, B.; Walk, S.F.; Ghesquière, B.; Krupnick, A.S.; et al. Metabolites released from apoptotic cells act as tissue messengers. Nature 2020, 580, 130–135. [Google Scholar] [CrossRef] [PubMed]
- Petrovsky, N.; Aguilar, J.C. Vaccine adjuvants: Current state and future trends. Immunol. Cell Biol. 2004, 82, 488–496. [Google Scholar] [CrossRef] [PubMed]
- Brake, D.A. Vaccinology for control of apicomplexan parasites: A simplified language of immune programming and its use in vaccine design. Int. J. Parasitol. 2002, 32, 509–515. [Google Scholar] [CrossRef]
- Qu, D.F.; Han, J.Z.; Du, A.F. Enhancement of protective immune response to recombinant Toxoplasma gondii ROP18 antigen by ginsenoside Re. Exp. Parasitol. 2013, 135, 234–239. [Google Scholar] [CrossRef] [PubMed]
- Sanchez, V.R.; Pitkowski, M.N.; Cuppari, A.V.F.; Rodriguez, F.M.; Fenoy, I.M.; Frank, F.M.; Goldman, A.; Corral, R.S.; Martin, V. Combination of CpG-oligodeoxynucleotides with recombinant ROP2 or GRA4 proteins induces protective immunity against Toxoplasma gondii infection. Exp. Parasitol. 2011, 128, 448–453. [Google Scholar] [CrossRef] [PubMed]
- Zheng, B.; Lu, S.H.; Tong, Q.B.; Kong, Q.M.; Lou, D. The virulence-related rhoptry protein 5 (ROP5) of Toxoplasma gondii is a novel vaccine candidate against toxoplasmosis in mice. Vaccine 2013, 31, 4578–4584. [Google Scholar] [CrossRef] [PubMed]
- Wang, T.; Yin, H.Q.; Li, Y.; Zhao, L.Z.; Sun, X.H.; Cong, H. Vaccination with recombinant adenovirus expressing multi-stage antigens of Toxoplasma gondii by the mucosal route induces higher systemic cellular and local mucosal immune responses than with other vaccination routes. Parasite 2017, 24, 12. [Google Scholar] [CrossRef][Green Version]
- Chen, J.; Zhou, D.H.; Li, Z.Y.; Petersen, E.; Huang, S.Y.; Song, H.Q.; Zhu, X.Q. Toxoplasma gondii: Protective immunity induced by rhoptry protein 9 (TgROP9) against acute toxoplasmosis. Exp. Parasitol. 2014, 139, 42–48. [Google Scholar] [CrossRef]
- Sonaimuthu, P.; Ching, X.T.; Fong, M.Y.; Kalyanasundaram, R.; Lau, Y.L. Induction of Protective Immunity against Toxoplasmosis in BALB/c Mice Vaccinated with Toxoplasma gondii Rhoptry-1. Front Microbiol. 2016, 7, 808. [Google Scholar] [CrossRef]
- Ghaffarifar, F.; Jafarimodrek, M.; Vazini, H.; Sharifi, Z.; Dalimi, A.; Dayer, M.S. Assessment of DNA vaccine encoding Toxoplasma gondii microneme complete gene and IL-12 as adjuvant in BALB/c mice. Iran J. Basic Med. Sci. 2019, 22, 901–907. [Google Scholar] [CrossRef] [PubMed]
- Wu, M.M.; An, R.; Chen, Y.; Chen, T.Y.; Wen, H.Y.; Yan, Q.; Shen, J.L.; Chen, L.J.; Du, J. Vaccination with recombinant Toxoplasma gondii CDPK3 induces protective immunity against experimental toxoplasmosis. Acta Trop. 2019, 199, 105148. [Google Scholar] [CrossRef] [PubMed]
- Stern, P.L. Key steps in vaccine development. Ann. Allergy Asthma Immunol. 2020, 125, 17–27. [Google Scholar] [CrossRef] [PubMed]
- Wipasa, J.; Riley, E.M. The immunological challenges of malaria vaccine development. Expert Opin. Biol. Ther. 2007, 7, 1841–1852. [Google Scholar] [CrossRef]
- Penner, R.C. Backbone Free Energy Estimator Applied to Viral Glycoproteins. J. Comput. Biol. 2020, 27, 1495–1508. [Google Scholar] [CrossRef]
- Sanchez-Trincado, J.L.; Gomez-Perosanz, M.; Reche, P.A. Fundamentals and Methods for T- and B-Cell Epitope Prediction. J. Immunol. Res. 2017, 2017, 2680160. [Google Scholar] [CrossRef]
- Correa, D.; Canedo-Solares, I.; Ortiz-Alegria, L.B.; Caballero-Ortega, H.; Rico-Torres, C.P. Congenital and acquired toxoplasmosis: Diversity and role of antibodies in different compartments of the host. Parasite Immunol. 2007, 29, 651–660. [Google Scholar] [CrossRef] [PubMed]
- Kang, H.; Remington, J.S.; Suzuki, Y. Decreased resistance of B cell-deficient mice to infection with Toxoplasma gondii despite unimpaired expression of IFN-gamma, TNF-alpha, and inducible nitric oxide synthase. J. Immunol. 2000, 164, 2629–2634. [Google Scholar] [CrossRef]
- Sayles, P.C.; Gibson, G.W.; Johnson, L.L. B cells are essential for vaccination-induced resistance to virulent Toxoplasma gondii. Infect. Immun. 2000, 68, 1026–1033. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.L.; Elsheikha, H.M.; Zhu, W.N.; Chen, K.; Li, T.T.; Yue, D.M.; Zhang, X.X.; Huang, S.Y.; Zhu, X.Q. Immunization with Toxoplasma gondii GRA17 Deletion Mutant Induces Partial Protection and Survival in Challenged Mice. Front. Immunol. 2017, 8, 730. [Google Scholar] [CrossRef]
- Grzybowski, M.M.; Dziadek, B.; Gatkowska, J.M.; Dzitko, K.; Dlugonska, H. Towards vaccine against toxoplasmosis: Evaluation of the immunogenic and protective activity of recombinant ROP5 and ROP18 Toxoplasma gondii proteins. Parasitol. Res. 2015, 114, 4553–4563. [Google Scholar] [CrossRef] [PubMed]
- Kugler, D.G.; Mittelstadt, P.R.; Ashwell, J.D.; Sher, A.; Jankovic, D. CD4(+) T cells are trigger and target of the glucocorticoid response that prevents lethal immunopathology in toxoplasma infection. J. Exp. Med. 2013, 210, 1919–1927. [Google Scholar] [CrossRef] [PubMed]
- Dupont, C.D.; Christian, D.A.; Selleck, E.M.; Pepper, M.; Leney-Greene, M.; Harms Pritchard, G.; Koshy, A.A.; Wagage, S.; Reuter, M.A.; Sibley, L.D.; et al. Parasite fate and involvement of infected cells in the induction of CD4+ and CD8+ T cell responses to Toxoplasma gondii. PLoS Pathog. 2014, 10, e1004047. [Google Scholar] [CrossRef] [PubMed]
- Lyons, R.E.; Anthony, J.P.; Ferguson, D.J.; Byrne, N.; Alexander, J.; Roberts, F.; Roberts, C.W. Immunological studies of chronic ocular toxoplasmosis: Up-regulation of major histocompatibility complex class I and transforming growth factor beta and a protective role for interleukin-6. Infect. Immun. 2001, 69, 2589–2595. [Google Scholar] [CrossRef]
- Sturge, C.R.; Benson, A.; Raetz, M.; Wilhelm, C.L.; Mirpuri, J.; Vitetta, E.S.; Yarovinsky, F. TLR-independent neutrophil-derived IFN-γ is important for host resistance to intracellular pathogens. Proc. Natl. Acad. Sci. USA 2013, 110, 10711–10716. [Google Scholar] [CrossRef] [PubMed]
- Matowicka-Karna, J.; Dymicka-Piekarska, V.; Kemona, H. Does Toxoplasma gondii Infection Affect the Levels of IgE and Cytokines (IL-5, IL-6, IL-10, IL-12, and TNF-alpha)? Clin. Dev. Immunol. 2009, 2009, 374696. [Google Scholar] [CrossRef]
- Ching, X.T.; Fong, M.Y.; Lau, Y.L. Evaluation of Immunoprotection Conferred by the Subunit Vaccines of GRA2 and GRA5 against Acute Toxoplasmosis in BALB/c Mice. Front. Microbiol. 2016, 7, 609. [Google Scholar] [CrossRef] [PubMed]
- Zheng, B.; Ding, J.; Chen, X.; Yu, H.; Lou, D.; Tong, Q.; Kong, Q.; Lu, S. Immuno-Efficacy of a T. gondii Secreted Protein with an Altered Thrombospondin Repeat (TgSPATR) As a Novel DNA Vaccine Candidate against Acute Toxoplasmosis in BALB/c Mice. Front. Microbiol. 2017, 8, 216. [Google Scholar] [CrossRef] [PubMed]
- Dubey, J.P. Mouse pathogenicity of Toxoplasma gondii isolated from a goat. Am. J. Vet. Res. 1980, 41, 427–429. [Google Scholar] [PubMed]
- Howe, D.K.; Honore, S.; Derouin, F.; Sibley, L.D. Determination of genotypes of Toxoplasma gondii strains isolated from patients with toxoplasmosis. J. Clin. Microbiol. 1997, 35, 1411–1414. [Google Scholar] [CrossRef] [PubMed]
- Melchor, S.J.; Ewald, S.E. Disease Tolerance in Toxoplasma Infection. Front. Cell Infect. Microbiol. 2019, 9, 185. [Google Scholar] [CrossRef] [PubMed]











Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yu, Z.; Lu, Y.; Liu, Z.; Aleem, M.T.; Liu, J.; Luo, J.; Yan, R.; Xu, L.; Song, X.; Li, X. Recombinant Toxoplasma gondii Ribosomal Protein P2 Modulates the Functions of Murine Macrophages In Vitro and Provides Immunity against Acute Toxoplasmosis In Vivo. Vaccines 2021, 9, 357. https://doi.org/10.3390/vaccines9040357
Yu Z, Lu Y, Liu Z, Aleem MT, Liu J, Luo J, Yan R, Xu L, Song X, Li X. Recombinant Toxoplasma gondii Ribosomal Protein P2 Modulates the Functions of Murine Macrophages In Vitro and Provides Immunity against Acute Toxoplasmosis In Vivo. Vaccines. 2021; 9(4):357. https://doi.org/10.3390/vaccines9040357
Chicago/Turabian StyleYu, Zhengqing, Yujia Lu, Zhaoyi Liu, Muhammad Tahir Aleem, Junlong Liu, Jianxun Luo, Ruofeng Yan, Lixin Xu, Xiaokai Song, and Xiangrui Li. 2021. "Recombinant Toxoplasma gondii Ribosomal Protein P2 Modulates the Functions of Murine Macrophages In Vitro and Provides Immunity against Acute Toxoplasmosis In Vivo" Vaccines 9, no. 4: 357. https://doi.org/10.3390/vaccines9040357
APA StyleYu, Z., Lu, Y., Liu, Z., Aleem, M. T., Liu, J., Luo, J., Yan, R., Xu, L., Song, X., & Li, X. (2021). Recombinant Toxoplasma gondii Ribosomal Protein P2 Modulates the Functions of Murine Macrophages In Vitro and Provides Immunity against Acute Toxoplasmosis In Vivo. Vaccines, 9(4), 357. https://doi.org/10.3390/vaccines9040357