
Deciphering the Molecular Basis for Attenuation of Flavobacterium columnare Strain Fc1723 Used as Modified Live Vaccine against Columnaris Disease


Abstract
1. Introduction
2. Materials and Methods
2.1. Bacterial Strains and Growth Conditions
2.2. Genomic DNA Extraction
2.3. Genomic Sequencing and Assembly
2.4. Gene Annotation and SNP Identification
2.5. Biofilm Quantification Assay
3. Results
3.1. Draft Genome of F. columnare B27 and 1723
3.2. Single-Nucleotide Polymorphisms Identified in Fc1723
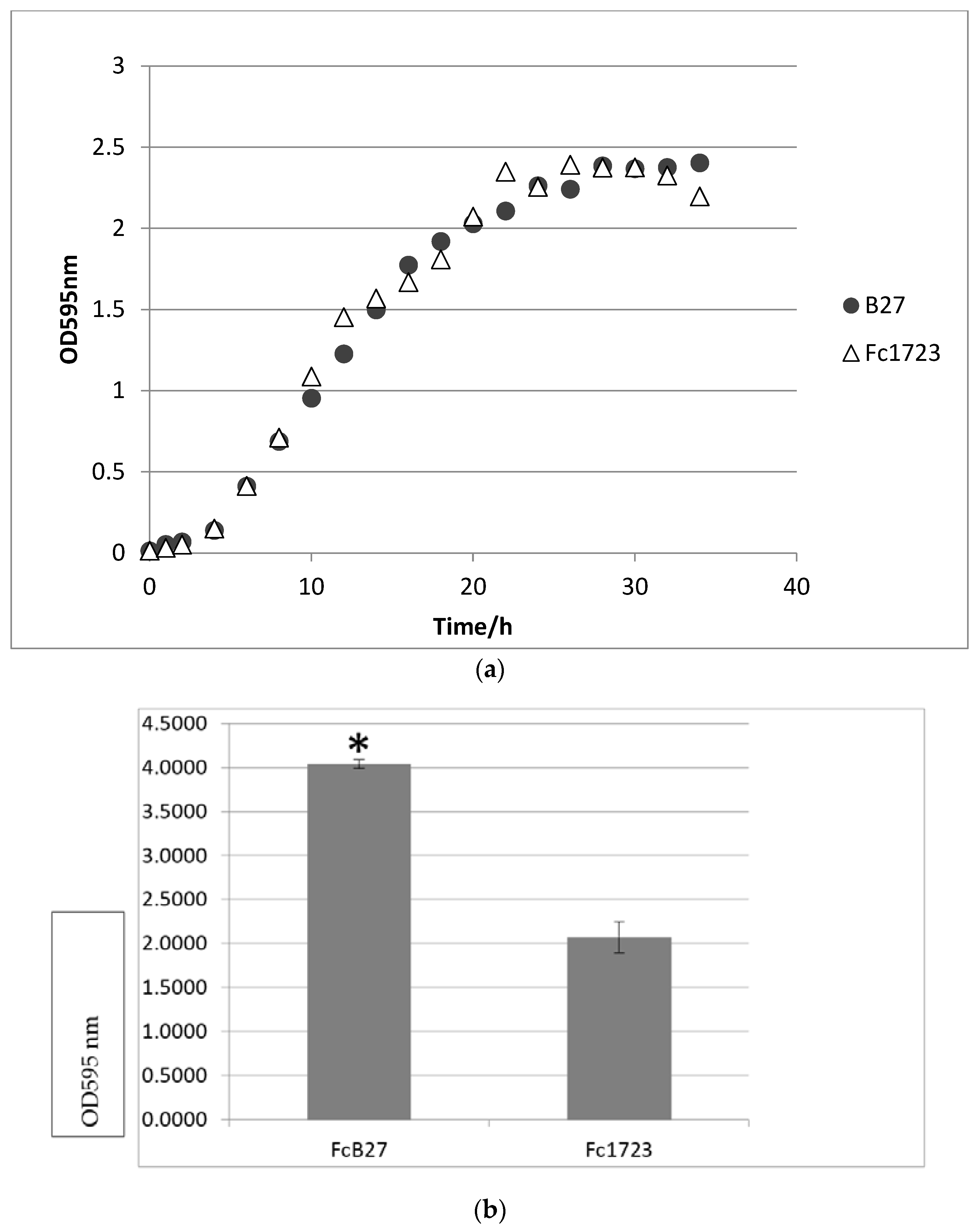
3.3. Growth, Gliding Motility, and Biofilm Formation
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Austin, B.; Austin, D.A. Bacterial Fish Pathogens: Disease of Farmed and Wild Fish; Springer: New York, NY, USA, 1999. [Google Scholar]
- USDA. Part I: Reference of Catfish Health and Production Practices in the United States, 2009; USDA, APHIS, VC, NAHMS: Fort Collins, CO, USA, 2010.
- Kunttu, H.; Sundberg, L.-R.; Pulkkinen, K.; Valtonen, E.T. Environment may be the source of Flavobacterium columanre outbreaks at fish farms. Environ. Microbiol. Rep. 2012, 4, 398–402. [Google Scholar] [CrossRef] [PubMed]
- Cai, W.; De La Fuente, L.; Arias, C.R. Biofilm formation by the fish pathogen Flavobacterium columnare: Development and parameters affecting surface attachment. Appl. Environ. Microbiol. 2013, 79, 5633–5642. [Google Scholar] [CrossRef] [PubMed]
- Arias, C.R.; LaFrentz, S.; Cai, W.; Olivares-Fuster, O. Adaptive response to starvation in the fish pathogen Flavobacterium columnare: Cell viability and ultrastructural changes. BMC Microbiol. 2012, 12, 266. [Google Scholar] [CrossRef]
- Tekedar, H.C.; Karsi, A.; Reddy, J.S.; Nho, S.W.; Kalindamar, S.; Lawrence, M.L. Comparative genomics and transcriptional analysis of Flavobacterium columnare strain ATCC 49512. Front. Microbiol. 2017, 8, 588. [Google Scholar] [CrossRef]
- Brudeseth, B.E.; Wiulsrod, R.; Fredriksen, B.N.; Kindmo, K.; Lokling, K.-E.; Bordevik, M.; Steine, N.; Klevan, A.; Gravningen, K. Status and future perspectives of vaccines for industrialised fin-fish farming. Fish Shellfish Immunol. 2013, 35, 1759–1768. [Google Scholar] [CrossRef] [PubMed]
- Woodrow, K.A.; Bennett, K.M.; Lo, D.D. Mucosal vaccine design and delivery. Annu. Rev. Biomed. Eng. 2012, 14, 17–46. [Google Scholar] [CrossRef]
- Shoemaker, C.A.; Klesius, P.H.; Evans, J.J.; Arias, C.R. Use of modified live vacccine in aquaculture. J. World Aquac. Soc. 2009, 40, 573–585. [Google Scholar] [CrossRef]
- Mohammed, H.; Olivares-Fuster, O.; LaFrentz, S.W.; Arias, C.R. New attenuated vaccine against columnaris disease in fish: Choosing the right parental strain is critical for vaccine efficacy. Vaccine 2013, 31, 5276–5280. [Google Scholar] [CrossRef]
- Triyanto, K.; Wakabayashi, H. Genotypic diversity of strains of Flavobacterium columnare from diseased fishes. Fish Pathol. 1999, 34, 65–71. [Google Scholar] [CrossRef]
- LaFrentz, B.R.; Waldbieser, G.C.; Welch, T.J.; Shoemaker, C.A. Intragenomic heterogeneity in the 16S rRNA genes of Flavobacterium columnare and standard protocols for genomovar assigment. J. Fish Dis. 2014, 37, 657–669. [Google Scholar] [CrossRef]
- Olivares-Fuster, O.; Baker, J.L.; Terhune, J.S.; Shoemaker, C.A.; Klesius, P.H.; Arias, C.R. Host-specific association between Flavobacterium columnare genomovars and fish species. Syst. Appl. Microbiol. 2007, 30, 624–633. [Google Scholar] [CrossRef] [PubMed]
- Shoemaker, C.A.; Olivares-Fuster, O.; Arias, C.R.; Klesius, P.H. Flavobacterium columnare genomovar influences mortality in channel catfish (Ictalurus punctatus). Vet. Microbiol. 2008, 127, 353–359. [Google Scholar] [CrossRef] [PubMed]
- Olivares-Fuster, O.; Arias, C.R. Development and characterization of rifampicin-resistant mutants from high virulent strains of Flavobacterium columnare. J. Fish Dis. 2011, 34, 385–394. [Google Scholar] [CrossRef] [PubMed]
- Shoemaker, C.A.; Arias, C.R.; Klesius, P.H.; Welker, T.L. Technique for identifying Flavobacterium columnare using whole-cell fatty acid profiles. J. Aquat. Anim. Health 2005, 17, 267–274. [Google Scholar] [CrossRef]
- Wei, Z.-G.; Zhang, S.-W. NPBSS: A new PacBio sequencing simulator for generating the continuous long reads with an empirical model. BMC Bioinform. 2018, 19, 177. [Google Scholar] [CrossRef]
- Overbeek, R.; Olson, R.; Pusch, G.D.; Olsen, G.J.; Davis, J.J.; Disz, T.; Edwards, R.A.; Gerdes, S.; Parrello, B.; Shukla, M. The SEED and the Rapid Annotation of microbial genomes using Subsystems Technology (RAST). Nucleic Acids Res. 2013, 42, D206–D214. [Google Scholar] [CrossRef]
- Aziz, R.K.; Bartels, D.; Best, A.A.; DeJongh, M.; Disz, T.; Edwards, R.A.; Formsma, K.; Gerdes, S.; Glass, E.M.; Kubal, M. The RAST Server: Rapid annotations using subsystems technology. BMC Genom. 2008, 9, 75. [Google Scholar] [CrossRef]
- Darling, A.C.; Mau, B.; Blattner, F.R.; Perna, N.T. Mauve: Multiple alignment of conserved genomic sequence with rearrangements. Genome Res. 2004, 14, 1394–1403. [Google Scholar] [CrossRef]
- Thorvaldsdóttir, H.; Robinson, J.T.; Mesirov, J.P. Integrative Genomics Viewer (IGV): High-performance genomics data visualization and exploration. Brief. Bioinform. 2013, 14, 178–192. [Google Scholar] [CrossRef]
- Choi, Y.; Chan, A.P. PROVEAN web server: A tool to predict the functional effect of amino acid substitutions and indels. Bioinformatics 2015, 31, 2745–2747. [Google Scholar] [CrossRef]
- Goldstein, B.P. Resistance to rifampicin: A review. J. Antibiot. 2014, 67, 625. [Google Scholar] [CrossRef] [PubMed]
- Alvarez, B.; Secades, P.; Prieto, M.; McBride, M.J.; Guijarro, J.A. A mutation in Flavobacterium psychrophilum tlpB inhibits gliding motility and induce biofilm formation. Appl. Environ. Microbiol. 2006, 72, 4044–4053. [Google Scholar] [CrossRef] [PubMed]
- Minor, P.D. Live attenuated vaccines: Historical successes and current challenges. Virology 2015, 479, 379–392. [Google Scholar] [CrossRef] [PubMed]
- Shoemaker, C.A.; Klesius, P.H. Protective immunity against enteric septicaemia in channel catfish, Ictalurus punctatus (Rafinesque), following controlled exposure to Edwardsiella ictaluri. J. Fish Dis. 1997, 20, 361–368. [Google Scholar] [CrossRef]
- Thune, R.L.; Fernandez, D.H.; Battista, J.R. An aroA Mutant of Edwardsiella ictaluri is Safe and Efficacious as a Live, Attenuated Vaccine. J. Aquat. Anim. Health 1999, 11, 358–372. [Google Scholar] [CrossRef]
- Detmer, A.; Glenting, J. Live bacterial vaccines—A review and identification of potential hazards. Microb. Cell Fact. 2006, 5, 23. [Google Scholar] [CrossRef]
- Montaraz, J.A.; Winter, A.J. Comparison of living and nonliving vaccines for Brucella abortus in BALB/c mice. Infect. Immun. 1986, 53, 245–251. [Google Scholar] [CrossRef]
- Dietrich, G.; Collioud, A.; Rothen, S.A. Developing and manufacturing attenuated live bacterial vaccines. BioPharm Int. 2008, 2008. [Google Scholar]
- Gliniewicz, K.; Wildung, M.; Orfe, L.H.; Wiens, G.D.; Cain, K.D.; Lahmers, K.K.; Snekvik, K.R.; Call, D.R. Potential mechanisms of attenuation for rifampicin-passaged strains of Flavobacterium psychrophilum. BMC Microbiol. 2015, 15, 179. [Google Scholar] [CrossRef]
- Schurig, G.G.; Roop, R.M., II; Bagchi, T.; Boyle, S.; Buhrman, D.; Sriranganathan, N. Biological properties of RB51; a stable rough strain of Brucella abortus. Vet. Microbiol. 1991, 28, 171–188. [Google Scholar] [CrossRef]
- LaFrentz, B.R.; LaPatra, S.E.; Call, D.R.; Cain, K.D. Isolation of rifampicin resistant Flavobacterium psychrophilum strains and their potential as live attenuated vaccine candidates. Vaccine 2008, 26, 5582–5589. [Google Scholar] [CrossRef] [PubMed]
- Klesius, P.H.; Shoemaker, C.A. Development and use of modified live Edwardsiella ictaluri vaccine against enteric septicemia of catfish. Adv. Vet. Med. 1999, 41, 523–537. [Google Scholar] [PubMed]
- Shoemaker, C.A.; Klesius, P.H.; Drennan, J.D.; Evans, J.J. Efficacy of a modified live Flavobacterium columnare vaccine in fish. Fish Shellfish Immunol. 2011, 30, 304–308. [Google Scholar] [CrossRef] [PubMed]
- Wehrli, W.; Knüsel, F.; Schmid, K.; Staehelin, M. Interaction of rifamycin with bacterial RNA polymerase. Proc. Natl. Acad. Sci. USA 1968, 61, 667–673. [Google Scholar] [CrossRef] [PubMed]
- Ovchinnikov, Y.A.; Monastyrskaya, G.S.; Guriev, S.O.; Kalinina, N.F.; Sverdlov, E.D.; Gragerov, A.I.; Bass, I.A.; Kiver, I.F.; Moiseyeva, E.P.; Igumnov, V.N. RNA polymerase rifampicin-resistance mutations in Escherichia coli: Sequence changes and dominance. Mol. Gen. Genet. 1983, 190, 344–348. [Google Scholar] [CrossRef]
- Jin, D.J.; Gross, C.A. Mapping and sequencing of mutations in the Escherichia coli rpoB gene that lead to rifampicin resistance. J. Mol. Biol. 1988, 202, 45–58. [Google Scholar] [CrossRef]
- Arias, C.R.; Shoemaker, C.A.; Evans, J.J.; Klesius, P.H. A comparative study of Edwardsiella ictaluri parent (EILO) and E. ictaluri rifampicin-mutant (RE-33) isolates using lipopolysaccharides, outer membrane proteins, fatty acids, Biolog, API 20E and genomic analyses. J. Dis. 2003, 26, 415–421. [Google Scholar] [CrossRef]
- Vemulapalli, R.; Mc Quiston, J.R.; Schurig, G.G.; Sriranganathan, N.; Halling, S.M.; Boyle, S.M. Identification of an IS711 element interrupting the wboA Gene of Brucella abortus vaccine Strain RB51 and a PCR Assay to distinguish strain RB51 from other Brucella species and strains. Clin. Diagn. Lab. Immunol. 1999, 6, 760–764. [Google Scholar] [CrossRef]
- Weissman, S.J.; Moseley, S.L.; Dykhuizen, D.E.; Sokurenko, E.V. Enterobacterial adhesins and the case for studying SNPs in bacteria. Trends Microbiol. 2003, 11, 115–117. [Google Scholar] [CrossRef]
- de Koning, H.; Diallinas, G. Nucleobase transporters. Mol. Membr. Biol. 2000, 17, 75–94. [Google Scholar]
- Vogels, G.V.D.; Van der Drift, C. Degradation of purines and pyrimidines by microorganisms. Bacteriol. Rev. 1976, 40, 403. [Google Scholar] [CrossRef] [PubMed]
- Smith, S.W.; Weiss, S.B.; Kennedy, E.P. The enzymatic dephosphorylation of phosphatidic acids. J. Biol. Chem. 1957, 228, 915–922. [Google Scholar] [CrossRef]
- Andrews, D.; Beames, B.; Summers, M.; Park, W. Characterization of the lipid acyl hydrolase activity of the major potato (Solanum tuberosum) tuber protein, patatin, by cloning and abundant expression in a baculovirus vector. Biochem. J. 1988, 252, 199–206. [Google Scholar] [CrossRef] [PubMed]
- Sato, H.; Frank, D.W.; Hillard, C.J.; Feix, J.B.; Pankhaniya, R.R.; Moriyama, K.; Finck-Barbançon, V.; Buchaklian, A.; Lei, M.; Long, R.M. The mechanism of action of the Pseudomonas aeruginosa-encoded type III cytotoxin, ExoU. EMBO J. 2003, 22, 2959–2969. [Google Scholar] [CrossRef] [PubMed]
- Phillips, R.M.; Six, D.A.; Dennis, E.A.; Ghosh, P. In vivo phospholipase activity of the Pseudomonas aeruginosa cytotoxin ExoU and protection of mammalian cells with phospholipase A2 inhibitors. J. Biol. Chem. 2003, 278, 41326–41332. [Google Scholar] [CrossRef]
- Banerji, S.; Flieger, A. Patatin-like proteins: A new family of lipolytic enzymes present in bacteria? Microbiology 2004, 150, 522–525. [Google Scholar] [CrossRef]
- Hermanson, G.T. Bioconjugate Techniques; Academic Press: Cambridge, MA, USA, 2013. [Google Scholar]
- Peek, J.A.; Taylor, R.K. Characterization of a periplasmic thiol: Disulfide interchange protein required for the functional maturation of secreted virulence factors of Vibrio cholerae. Proc. Natl. Acad. Sci. USA 1992, 89, 6210–6214. [Google Scholar] [CrossRef]
- Suzuki, T.; Itoh, A.; Ichihara, S.; Mizushima, S. Characterization of the sppA gene coding for protease IV, a signal peptide peptidase of Escherichia coli. J. Bacteriol. 1987, 169, 2523–2528. [Google Scholar] [CrossRef]
- Engel, L.S.; Hill, J.M.; Caballero, A.R.; Green, L.C.; O’Callaghan, R.J. Protease IV, a unique extracellular protease and virulence factor from Pseudomonas aeruginosa. J. Biol. Chem. 1998, 273, 16792–16797. [Google Scholar] [CrossRef]
- Kunttu, H.; Jokinen, E.; Valtonen, E.; Sundberg, L.R. Virulent and nonvirulent Flavobacterium columnare colony morphologies: Characterization of chondroitin AC lyase activity and adhesion to polystyrene. J. Appl. Microbiol. 2011, 111, 1319–1326. [Google Scholar] [CrossRef]
- Mount, D.W. Using the basic local alignment search tool (BLAST). Cold Spring Harb. Protoc. 2007, 2007, pdb.top17. [Google Scholar] [CrossRef] [PubMed]


{kind=link}
| SNP Contig | SNP Position a | Reference Position b | SNP Pattern c | Amino Acid Change d | Annotation e | Functional Group |
|---|---|---|---|---|---|---|
| scf7180000000036 | 125027 | 2071493 | G > A | Asn469Lys | DNA-directed RNA polymerase beta subunit | Gene transcription |
| scf7180000000036 | 125076 | 2071592 | G > T | His486Tyr | DNA-directed RNA polymerase beta subunit | Gene transcription |
| scf7180000000039 | 83013 | 1656107 | C > A | Gly240stop | Nucleoside permease | Nucleosides transporter |
| scf7180000000044 | 91216 | 121892 | G > T | Glu194Stop | Phosphoesterase | Lipid metabolism |
| scf7180000000039 | 132995 | 1700354 | G > A | Gly386Arg | Protein-disulfide reductase | Protein biosynthesis |
| scf7180000000044 | 114715 | 145396 | C > A | Phe577Cys | Patatin | Energy storage |
| scf7180000000043 | 500199 | 2767414 | C > A | Ala470Asp | Protease IV | Toxin secretion |
| scf7180000000036 | 99642 | 2048192 | T > G | Ile2902Met | Thioredoxin reductase | Redox regulation/biofilm |
| scf7180000000044 | 41423 | 74284 | T > C | Phe238Ser | Dynein heavy chain | Unknown |
| scf7180000000044 | 41416 | 74277 | C > T | Leu236Leu | Dynein heavy chain | Unknown |
| scf7180000000043 | 534202 | 2801439 | C > T | Glu167Glu | Heme O synthase/protoheme IX farnesyltransferase | Protein biosynthesis |
| scf7180000000047 | 741630 | 1267503 | G > A | NA | Hypothetical protein | NA |
| scf7180000000044 | 1361 | NF | C > T | NA | Unknown region | NA |
| scf7180000000044 | 1147 | NF | A > G | NA | Unknown region | NA |
| scf7180000000044 | 1363 | NF | G > A | NA | Unknown region | NA |
| scf7180000000047 | 249370 | 804839 | G > T | NA | Unknown region | NA |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Cai, W.; Arias, C.R. Deciphering the Molecular Basis for Attenuation of Flavobacterium columnare Strain Fc1723 Used as Modified Live Vaccine against Columnaris Disease. Vaccines 2021, 9, 1370. https://doi.org/10.3390/vaccines9111370
Cai W, Arias CR. Deciphering the Molecular Basis for Attenuation of Flavobacterium columnare Strain Fc1723 Used as Modified Live Vaccine against Columnaris Disease. Vaccines. 2021; 9(11):1370. https://doi.org/10.3390/vaccines9111370
Chicago/Turabian StyleCai, Wenlong, and Covadonga R. Arias. 2021. "Deciphering the Molecular Basis for Attenuation of Flavobacterium columnare Strain Fc1723 Used as Modified Live Vaccine against Columnaris Disease" Vaccines 9, no. 11: 1370. https://doi.org/10.3390/vaccines9111370
APA StyleCai, W., & Arias, C. R. (2021). Deciphering the Molecular Basis for Attenuation of Flavobacterium columnare Strain Fc1723 Used as Modified Live Vaccine against Columnaris Disease. Vaccines, 9(11), 1370. https://doi.org/10.3390/vaccines9111370