Humoral Immune Response of Thai Dogs after Oral Vaccination against Rabies with the SPBN GASGAS Vaccine Strain

 , , and
, , and
Abstract
1. Introduction
2. Materials and Methods
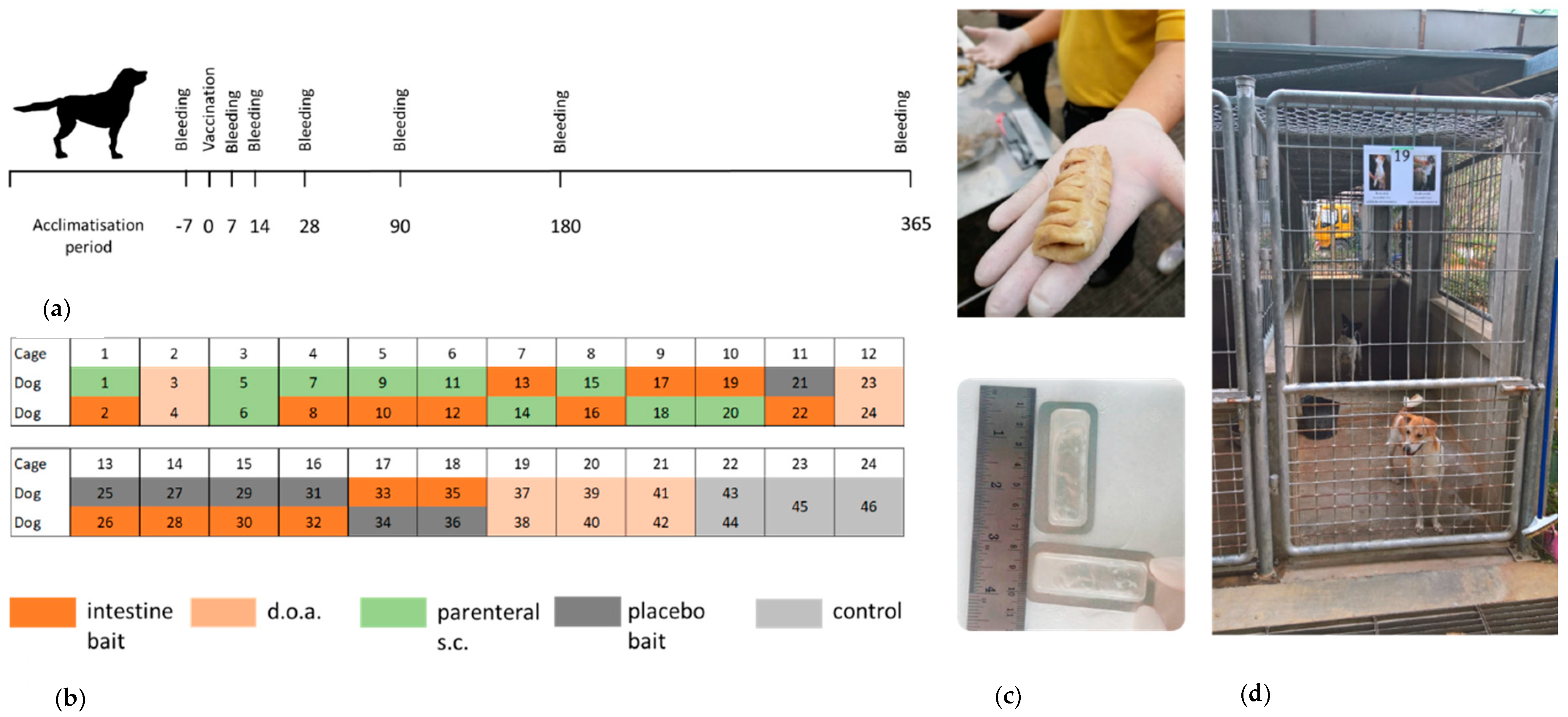
2.1. Animals and Housing
2.2. Study Design
2.3. Vaccination
2.4. Sampling and Testing Procedure
2.5. Diagnostic Assays
2.6. Statistical Analysis
3. Results
3.1. Bait Acceptance and Safety of the Vaccine
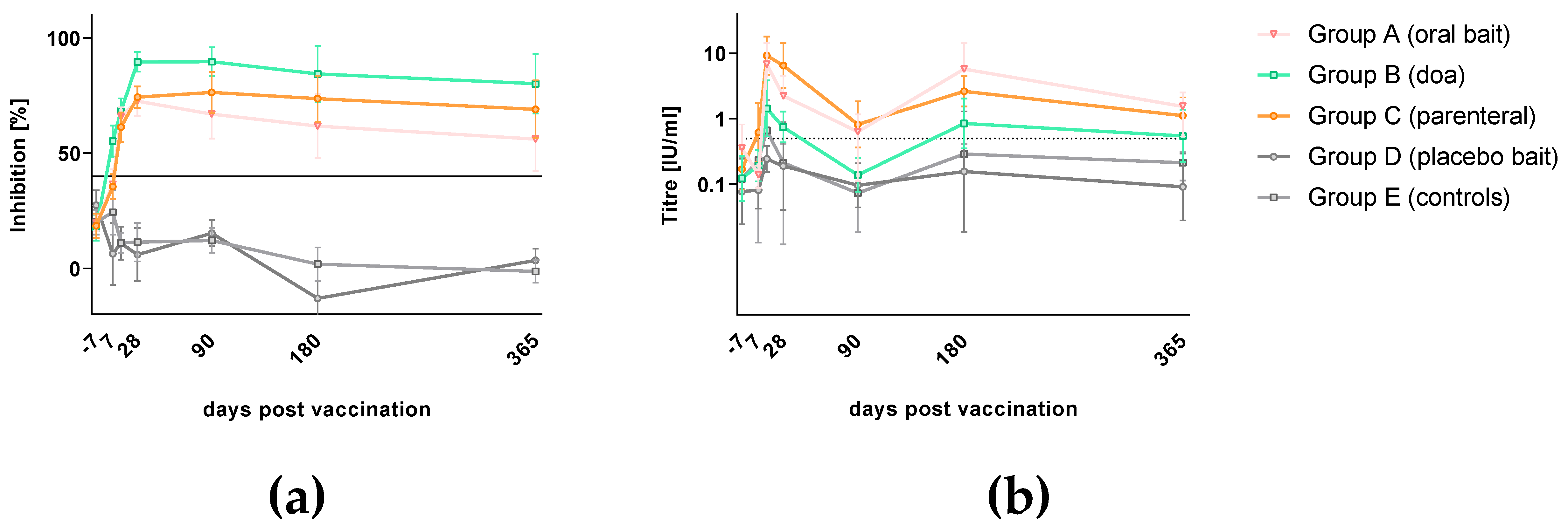
3.2. Rabies Specific Immune Response
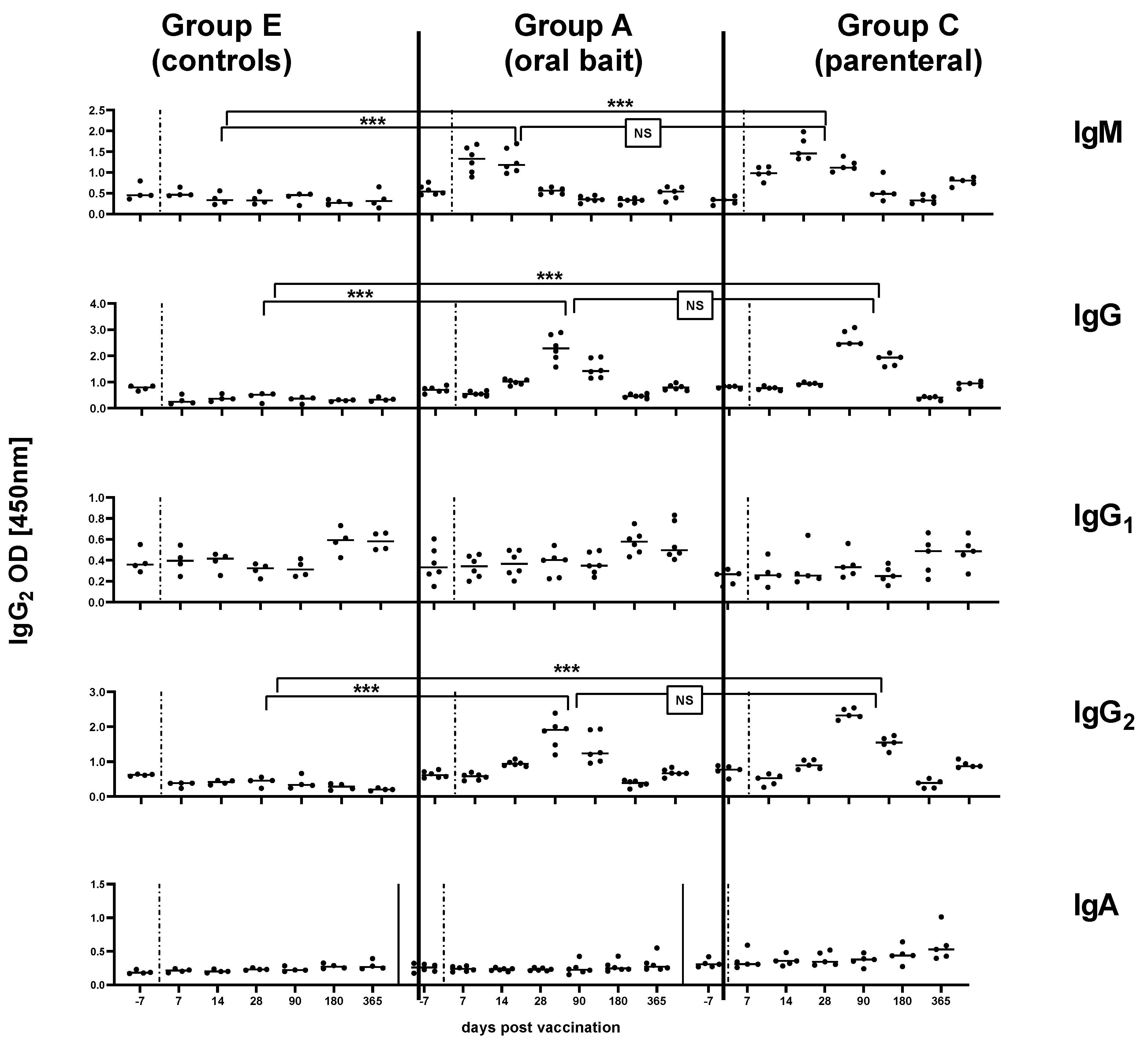
3.3. Kinetics of Immunoglobulin Classes and Isotypes
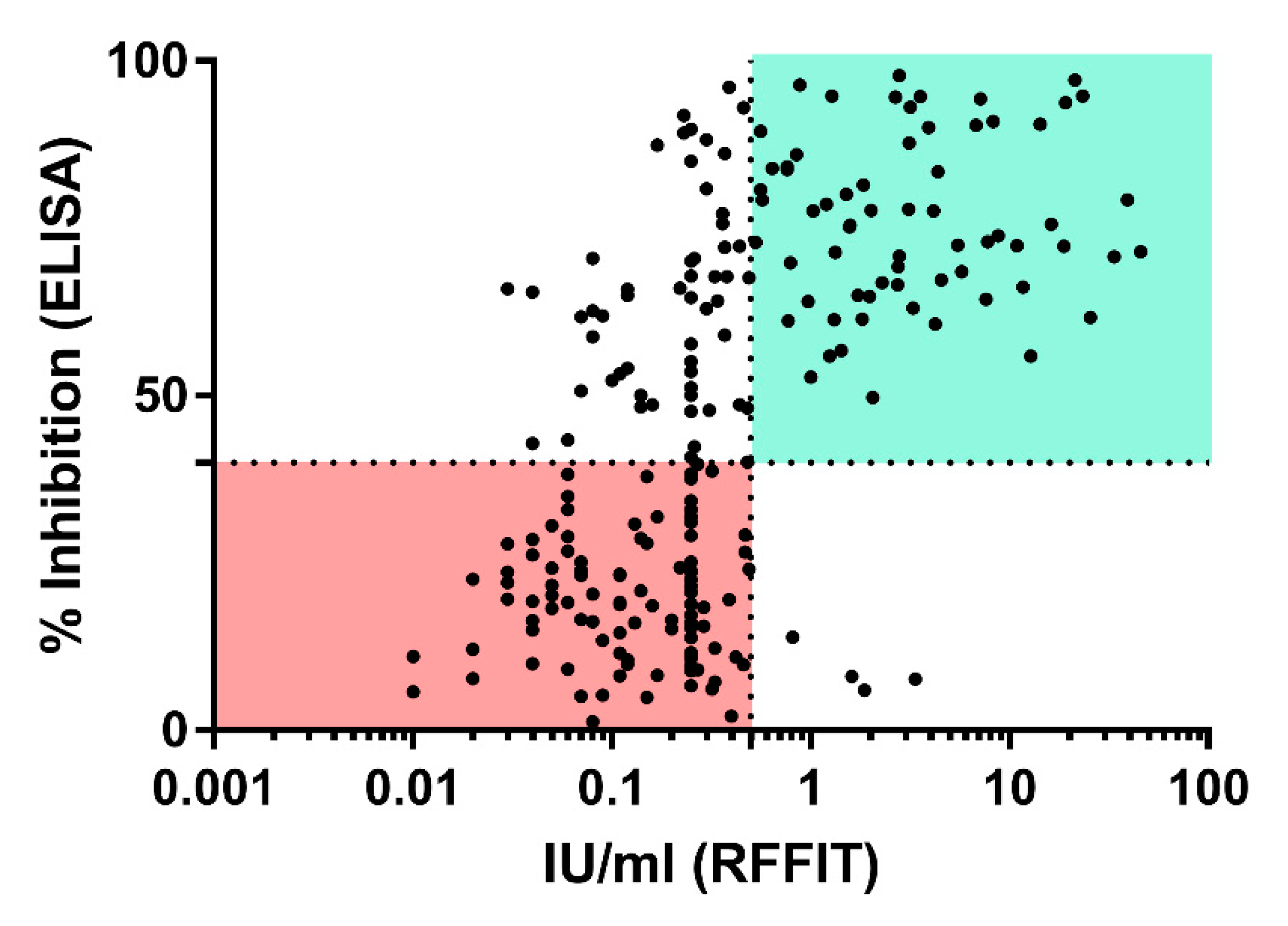
3.4. rVBA Versus rVNA
3.5. Seroconversion Rates
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Abbreviations
| mdv | mass dog vaccination |
| ORV | oral rabies vaccination |
| dpv | days post vaccination |
| doa | direct oral application |
| sc | subcutaneously |
| rVNA | Rabies virus neutralizing antibodies |
| rVBA | Rabies virus binding antibodies |
| RFFIT | Rapid Fluorescent Focus Inhibition Test |
| ELISA | Enzyme-linked Immunosorbent Assay |
References
- Hampson, K.; Coudeville, L.; Lembo, T.; Sambo, M.; Kieffer, A.; Attlan, M.; Barrat, J.; Blanton, J.D.; Briggs, D.J.; Cleaveland, S.; et al. Estimating the global burden of endemic canine rabies. PLoS Neglect. Trop. Dis. 2015, 9, e0003709. [Google Scholar] [CrossRef]
- Minghui, R.; Stone, M.; Semedo, M.H.; Nel, L. New global strategic plan to eliminate dog-mediated rabies by 2030. Lancet Glob. Health 2018, 6, e828–e829. [Google Scholar] [CrossRef]
- Abela-Ridder, B.; Balogh de, K.; Kessels, J.A.; Dieuzy-Labaye, I.; Torres, G. Global rabies control: The role of international organisations and the Global Strategic Plan to eliminate dog-mediated human rabies. Rev. Sci. Tech. 2018, 37, 741–749. [Google Scholar] [CrossRef] [PubMed]
- Shwiff, S.; Hampson, K.; Anderson, A. Potential economic benefits of eliminating canine rabies. Antivir. Res. 2013, 98, 352–356. [Google Scholar] [CrossRef] [PubMed]
- Vigilato, M.A.; Clavijo, A.; Knobl, T.; Silva, H.M.; Cosivi, O.; Schneider, M.C.; Leanes, L.F.; Belotto, A.J.; Espinal, M.A. Progress towards eliminating canine rabies: Policies and perspectives from Latin America and the Caribbean. Philos. Trans. R. Soc. Lond. B Biol. Sci. 2013, 368, 20120143. [Google Scholar] [CrossRef] [PubMed]
- Wallace, R.M.; Undurraga, E.A.; Blanton, J.D.; Cleaton, J.; Franka, R. Elimination of Dog-Mediated Human Rabies Deaths by 2030: Needs Assessment and Alternatives for Progress Based on Dog Vaccination. Front. Vet. Sci. 2017, 4, 9. [Google Scholar] [CrossRef]
- Cliquet, F.; Guiot, A.L.; Aubert, M.; Robardet, E.; Rupprecht, C.E.; Meslin, F.X. Oral vaccination of dogs: A well-studied and undervalued tool for achieving human and dog rabies elimination. Vet. Res. 2018, 49, 61. [Google Scholar] [CrossRef]
- Rupprecht, C.E.; Kuzmin, I.V.; Yale, G.; Nagarajan, T.; Meslin, F.X. Priorities in applied research to ensure programmatic success in the global elimination of canine rabies. Vaccine 2019, 37, A77–A84. [Google Scholar] [CrossRef]
- Rupprecht, C.E.; Abela-Ridder, B.; Abila, R.; Amparo, A.C.; Banyard, A.; Blanton, J.; Chanachai, K.; Dallmeier, K.; de Balogh, K.; Del Rio Vilas, V.; et al. Towards rabies elimination in the Asia-Pacific region: From theory to practice. Biologicals 2020, 64, 83–95. [Google Scholar] [CrossRef]
- Mills, G. Oral vaccines could be key to mass rabies dog vaccination. Vet. Rec. 2019, 184, 464–465. [Google Scholar] [CrossRef]
- Vos, A. Oral vaccination of dogs against rabies. Intern Anim. Health J. 2019, 6, 25–29. [Google Scholar]
- Wallace, R.M.; Undurraga, E.A.; Gibson, A.; Boone, J.; Pieracci, E.G.; Gamble, L.; Blanton, J.D. Estimating the effectiveness of vaccine programs in dog populations. Epidemiol. Infect. 2019, 147, e247. [Google Scholar] [CrossRef] [PubMed]
- Gibson, A.D.; Yale, G.; Vos, A.; Corfmat, J.; Airikkala-Otter, I.; King, A.; Wallace, R.M.; Gamble, L.; Handel, I.G.; Mellanby, R.J.; et al. Oral bait handout as a method to access roaming dogs for rabies vaccination in Goa, India: A proof of principle study. Vaccine X 2019, 1, 100015. [Google Scholar] [CrossRef]
- Lembo, T.; Hampson, K.; Kaare, M.T.; Ernest, E.; Knobel, D.; Kazwala, R.R.; Haydon, D.T.; Cleaveland, S. The feasibility of canine rabies elimination in Africa: Dispelling doubts with data. PLoS Neglect. Trop. Dis. 2010, 4, e626. [Google Scholar] [CrossRef] [PubMed]
- Gibson, A.D.; Mazeri, S.; Yale, G.; Desai, S.; Naik, V.; Corfmat, J.; Ortmann, S.; King, A.; Müller, T.; Handel, I.; et al. Development of a Non-Meat-Based, Mass Producible and Effective Bait for Oral Vaccination of Dogs against Rabies in Goa State, India. Trop. Med. Infect. Dis. 2019, 4, 118. [Google Scholar] [CrossRef]
- Bender, S.; Bergman, D.; Vos, A.; Martin, A.; Chipman, R. Field Studies Evaluating Bait Acceptance and Handling by Dogs in Navajo Nation, USA. Trop. Med. Infect. Dis. 2017, 2, 17. [Google Scholar] [CrossRef]
- Kasemsuwan, S.; Chanachai, K.; Pinyopummintr, T.; Leelalapongsathon, K.; Sujit, K.; Vos, A. Field Studies Evaluating Bait Acceptance and Handling by Free-Roaming Dogs in Thailand. Vet. Sci. 2018, 5, 47. [Google Scholar] [CrossRef]
- Darkaoui, S.; Boue, F.; Demerson, J.M.; Fassi Fihri, O.; Yahia, K.I.; Cliquet, F. First trials of oral vaccination with rabies SAG2 dog baits in Morocco. Clin. Exp. Vaccine Res. 2014, 3, 220–226. [Google Scholar] [CrossRef]
- Head, J.R.; Vos, A.; Blanton, J.; Müller, T.; Chipman, R.; Pieracci, E.G.; Cleaton, J.; Wallace, R. Environmental distribution of certain modified live-virus vaccines with a high safety profile presents a low-risk, high-reward to control zoonotic diseases. Sci. Rep. 2019, 9, 6783. [Google Scholar] [CrossRef]
- Gibson, A.D.; Wallace, R.M.; Rahman, A.; Bharti, O.K.; Isloor, S.; Lohr, F.; Gamble, L.; Mellanby, R.J.; King, A.; Day, M.J. Reviewing Solutions of Scale for Canine Rabies Elimination in India. Trop. Med. Infect. Dis. 2020, 5, 47. [Google Scholar] [CrossRef]
- Undurraga, E.A.; Millien, M.F.; Allel, K.; Etheart, M.D.; Cleaton, J.; Ross, Y.; Wallace, R.M. Costs and effectiveness of alternative dog vaccination strategies to improve dog population coverage in rural and urban settings during a rabies outbreak. Vaccine 2020, 38, 6162–6173. [Google Scholar] [CrossRef] [PubMed]
- Müller, W.; Güzel, T.; Auylan, O.; Kaya, C.; Cox, J.; Schneider, L. The feasibility of oral vaccination of dogs in Turkey—A European Union supported project. J. Etlik Vet. Microbiol. 1998, 9, 61–73. [Google Scholar]
- Aylan, O.; Vos, A. Efficacy of oral rabies vaccine baits in indigenous Turkish dogs. Infect. Dis. Rev. 2000, 2, 74–77. [Google Scholar]
- Aylan, O.; Vos, A. Efficacy studies with SAD B19 in Turkish dogs. J. Etlik Vet. Microbiol. 1998, 9, 93–102. [Google Scholar]
- Cliquet, F.; Gurbuxani, J.P.; Pradhan, H.K.; Pattnaik, B.; Patil, S.S.; Regnault, A.; Begouen, H.; Guiot, A.L.; Sood, R.; Mahl, P.; et al. The safety and efficacy of the oral rabies vaccine SAG2 in Indian stray dogs. Vaccine 2007, 25, 257–264. [Google Scholar] [CrossRef]
- Cliquet, F.; Guiot, A.L.; Schumacher, C.; Maki, J.; Cael, N.; Barrat, J. Efficacy of a square presentation of V-RG vaccine baits in red fox, domestic dog and raccoon dog. Dev. Biol. 2008, 131, 257–264. [Google Scholar]
- Hammami, S.; Schumacher, C.; Cliquet, F.; Tlatli, A.; Aubert, A.; Aubert, M. Vaccination of Tunisian dogs with the lyophilised SAG2 oral rabies vaccine incorporated into the DBL2 dog bait. Vet. Res. 1999, 30, 607–613. [Google Scholar]
- Faber, M.; Faber, M.L.; Papaneri, A.; Bette, M.; Weihe, E.; Dietzschold, B.; Schnell, M.J. A Single Amino Acid Change in Rabies Virus Glycoprotein Increases Virus Spread and Enhances Virus Pathogenicity. J. Virol. 2005, 79, 14141–14148. [Google Scholar] [CrossRef]
- Faber, M.; Pulmanausahakul, R.; Hodawadekar, S.S.; Spitsin, S.; McGettigan, J.P.; Schnell, M.J.; Dietzschold, B. Overexpression of the rabies virus glycoprotein results in enhancement of apoptosis and antiviral immune response. J. Virol. 2002, 76, 3374–3381. [Google Scholar] [CrossRef]
- Freuling, C.M.; Eggerbauer, E.; Finke, S.; Kaiser, C.; Kaiser, C.; Kretzschmar, A.; Nolden, T.; Ortmann, S.; Schröder, C.; Teifke, J.P.; et al. Efficacy of the oral rabies virus vaccine strain SPBN GASGAS in foxes and raccoon dogs. Vaccine 2019, 37, 4750–4757. [Google Scholar] [CrossRef]
- Freuling, C.M.; Kamp, V.T.; Klein, A.; Günther, M.; Zaeck, L.; Potratz, M.; Eggerbauer, E.; Bobe, K.; Kaiser, C.; Kretzschmar, A.; et al. Long-Term Immunogenicity and Efficacy of the Oral Rabies Virus Vaccine Strain SPBN GASGAS in Foxes. Viruses 2019, 11, 790. [Google Scholar] [CrossRef] [PubMed]
- Moore, S.; Gilbert, A.; Vos, A.; Freuling, C.M.; Ellis, C.; Kliemt, J.; Müller, T. Rabies Virus Antibodies from Oral Vaccination as a Correlate of Protection against Lethal Infection in Wildlife. Trop. Med. Infect. Dis. 2017, 2, 31. [Google Scholar] [CrossRef] [PubMed]
- Wasniewski, M.; Guiot, A.L.; Schereffer, J.L.; Tribout, L.; Mähar, K.; Cliquet, F. Evaluation of an ELISA to detect rabies antibodies in orally vaccinated foxes and raccoon dogs sampled in the field. J. Virol. Methods 2013, 187, 264–270. [Google Scholar] [CrossRef] [PubMed]
- Sagaraaeranee, P.; Puanghat, A.; Kasempimolparn, S.; Khawplod, P. Efficacy of oral rabies vaccine in dogs in Thailand. In Rabies Control in Asia; Dodet, B., Meslin, F.X., Eds.; John Libbey Eurotext: Montrouge, France, 2001. [Google Scholar]
- Vos, A.; Freuling, C.M.; Hundt, B.; Kaiser, C.; Nemitz, S.; Neubert, A.; Nolden, T.; Teifke, J.P.; Te Kamp, V.; Ulrich, R.; et al. Oral vaccination of wildlife against rabies: Differences among host species in vaccine uptake efficiency. Vaccine 2017, 35, 3938–3944. [Google Scholar] [CrossRef] [PubMed]
- Te Kamp, V.; Freuling, C.M.; Vos, A.; Schuster, P.; Kaiser, C.; Ortmann, S.; Kretzschmar, A.; Nemitz, S.; Eggerbauer, E.; Ulrich, R.; et al. Responsiveness of various reservoir species to oral rabies vaccination correlates with differences in vaccine uptake of mucosa associated lymphoid tissues. Sci. Rep. 2020, 10, 2919. [Google Scholar] [CrossRef]
- Vidarsson, G.; Dekkers, G.; Rispens, T. IgG subclasses and allotypes: From structure to effector functions. Front. Immunol. 2014, 5, 520. [Google Scholar] [CrossRef]
- Aubert, M.F. Practical significance of rabies antibodies in cats and dogs. Rev. Sci. Tech. 1992, 11, 735–760. [Google Scholar] [CrossRef]
- OIE. Chapter 3.1.17. Rabies (Infection with Rabies Virus and Other Lyssaviruses) (NB: Version Adopted in May 2018). In Manual of Diagnostic Tests and Vaccines for Terrestrial Animals; OIE: Paris, France, 2018. [Google Scholar]
- Moore, S.M.; Hanlon, C.A. Rabies-Specific Antibodies: Measuring Surrogates of Protection against a Fatal Disease. PLoS Neglect. Trop. Dis. 2010, 4, e595. [Google Scholar] [CrossRef]
- Smith, T.G.; Millien, M.; Vos, A.; Fracciterne, F.A.; Crowdis, K.; Chirodea, C.; Medley, A.; Chipman, R.; Qin, Y.; Blanton, J.; et al. Evaluation of immune responses in dogs to oral rabies vaccine under field conditions. Vaccine 2019, 37, 4743–4749. [Google Scholar] [CrossRef]
- Ortmann, S.; Kretzschmar, A.; Kaiser, C.; Lindner, T.; Freuling, C.; Schuster, P.; Mueller, T.; Vos, A. In Vivo Safety Studies with SPBN GASGAS in the Frame of Oral Vaccination of Foxes and Raccoon Dogs against Rabies. Front. Vet. Sci. 2018, 5, 91. [Google Scholar] [CrossRef]
- Knowlton, F.F.; Roetto, M.; Briggs, D. Serological responses of coyotes to two commercial rabies vaccines. J. Wildl. Dis. 2001, 37, 798–802. [Google Scholar] [CrossRef] [PubMed]
- Randall, K.L. Generating humoral immune memory following infection or vaccination. Expert Rev. Vaccines 2010, 9, 1083–1093. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Groups | ||||||
|---|---|---|---|---|---|---|
| dpv | Assay | A SPBN GASGAS (bait) | B SPBN GASGAS (doa) | C Bayovac * R (sc) | D Placebo Bait | E Control |
| −7 | ELISA | 0/15 | 0/10 | 0/10 | 0/7 | 0/4 |
| RFFIT | 1/15 | 0/10 | 1/10 | 0/7 | 0/4 | |
| 7 | ELISA | 5/15 | 3/10 | 10/10 | 1/7 * | 0/4 |
| RFFIT | 0/15 | 0/10 | 4/10 | 0/7 | 0/4 | |
| 14 | ELISA | 14/15 | 10/10 | 10/10 | 0/7 | 0/4 |
| RFFIT | 10/15 | 8/10 | 10/10 | 0/7 | 1/4 | |
| 28 | ELISA | 15/15 | 10/10 | 10/10 | 0/7 | 0/4 |
| RFFIT | 8/15 | 7/10 | 10/10 | 1/7 | 0/4 | |
| 90 | ELISA | 14/15 ** | 10/10 | 10/10 | 0/6 | 0/4 |
| RFFIT | 4/15 | 1/10 | 5/10 | 0/6 | 0/4 | |
| 180 | ELISA | 14/15 | 9/10 | 10/10 | 0/6 | 0/4 |
| RFFIT | 8/15 | 6/10 | 10/10 | 1/6 *** | 0/4 | |
| 365 | ELISA | 13/15 | 8/10 | 9/10 | 0/6 | 0/4 |
| RFFIT | 7/15 | 4/10 | 7/10 | 0/6 | 0/4 | |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Leelahapongsathon, K.; Kasemsuwan, S.; Pinyopummintr, T.; Boodde, O.; Phawaphutayanchai, P.; Aiyara, N.; Bobe, K.; Vos, A.; Friedrichs, V.; Müller, T.; et al. Humoral Immune Response of Thai Dogs after Oral Vaccination against Rabies with the SPBN GASGAS Vaccine Strain. Vaccines 2020, 8, 573. https://doi.org/10.3390/vaccines8040573
Leelahapongsathon K, Kasemsuwan S, Pinyopummintr T, Boodde O, Phawaphutayanchai P, Aiyara N, Bobe K, Vos A, Friedrichs V, Müller T, et al. Humoral Immune Response of Thai Dogs after Oral Vaccination against Rabies with the SPBN GASGAS Vaccine Strain. Vaccines. 2020; 8(4):573. https://doi.org/10.3390/vaccines8040573
Chicago/Turabian StyleLeelahapongsathon, Kansuda, Suwicha Kasemsuwan, Tanu Pinyopummintr, Orawan Boodde, Parinya Phawaphutayanchai, Nirut Aiyara, Katharina Bobe, Ad Vos, Virginia Friedrichs, Thomas Müller, and et al. 2020. "Humoral Immune Response of Thai Dogs after Oral Vaccination against Rabies with the SPBN GASGAS Vaccine Strain" Vaccines 8, no. 4: 573. https://doi.org/10.3390/vaccines8040573
APA StyleLeelahapongsathon, K., Kasemsuwan, S., Pinyopummintr, T., Boodde, O., Phawaphutayanchai, P., Aiyara, N., Bobe, K., Vos, A., Friedrichs, V., Müller, T., Freuling, C. M., & Chanachai, K. (2020). Humoral Immune Response of Thai Dogs after Oral Vaccination against Rabies with the SPBN GASGAS Vaccine Strain. Vaccines, 8(4), 573. https://doi.org/10.3390/vaccines8040573