Alterations in the Human Plasma Lipidome in Response to Tularemia Vaccination

 , , , , , and
, , , , , and
Abstract
1. Introduction
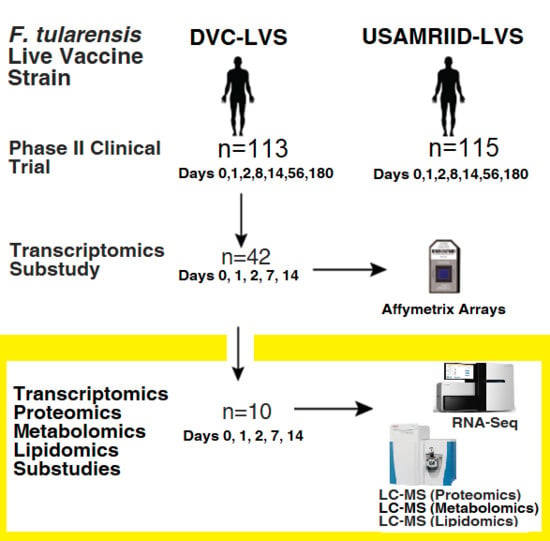
2. Materials and Methods
2.1. Study Design
2.2. Standards
2.3. Lipid Extraction
2.4. Mass Spectrometry
2.5. Lipid Quantification
2.6. Data Analysis
Missing Values and Baseline Calculations
2.7. Statistics
2.8. Transcriptomics Data
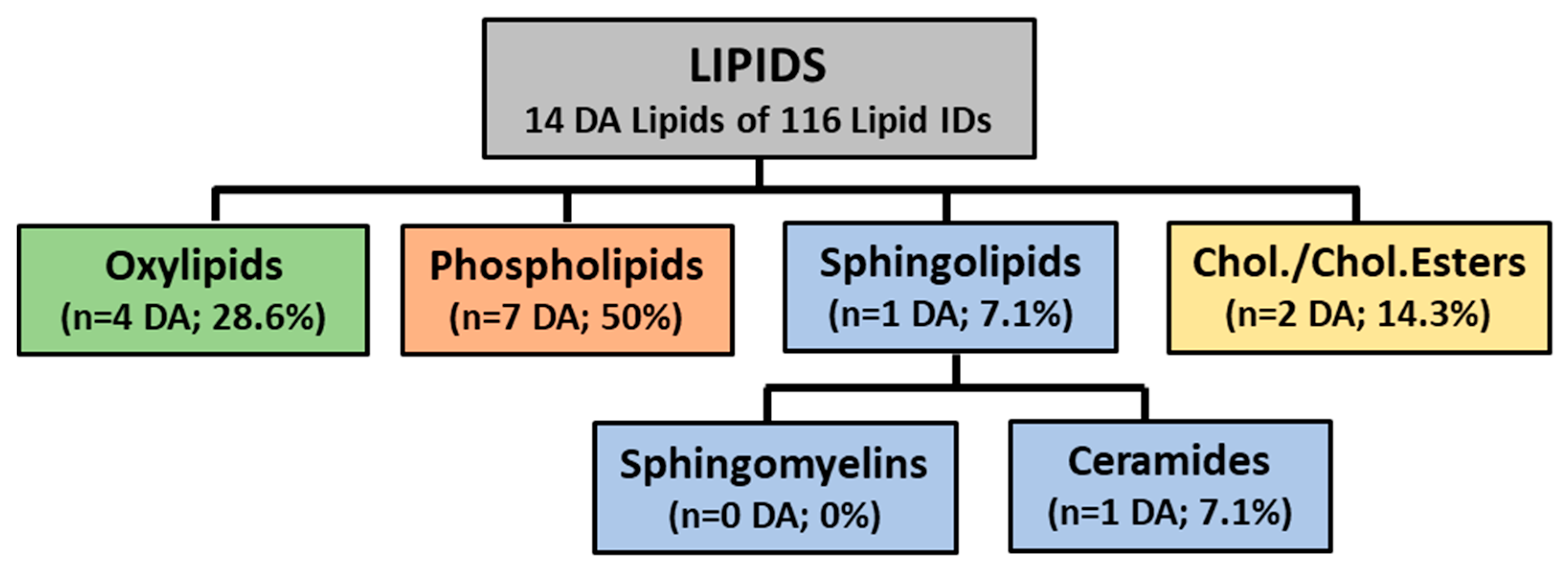
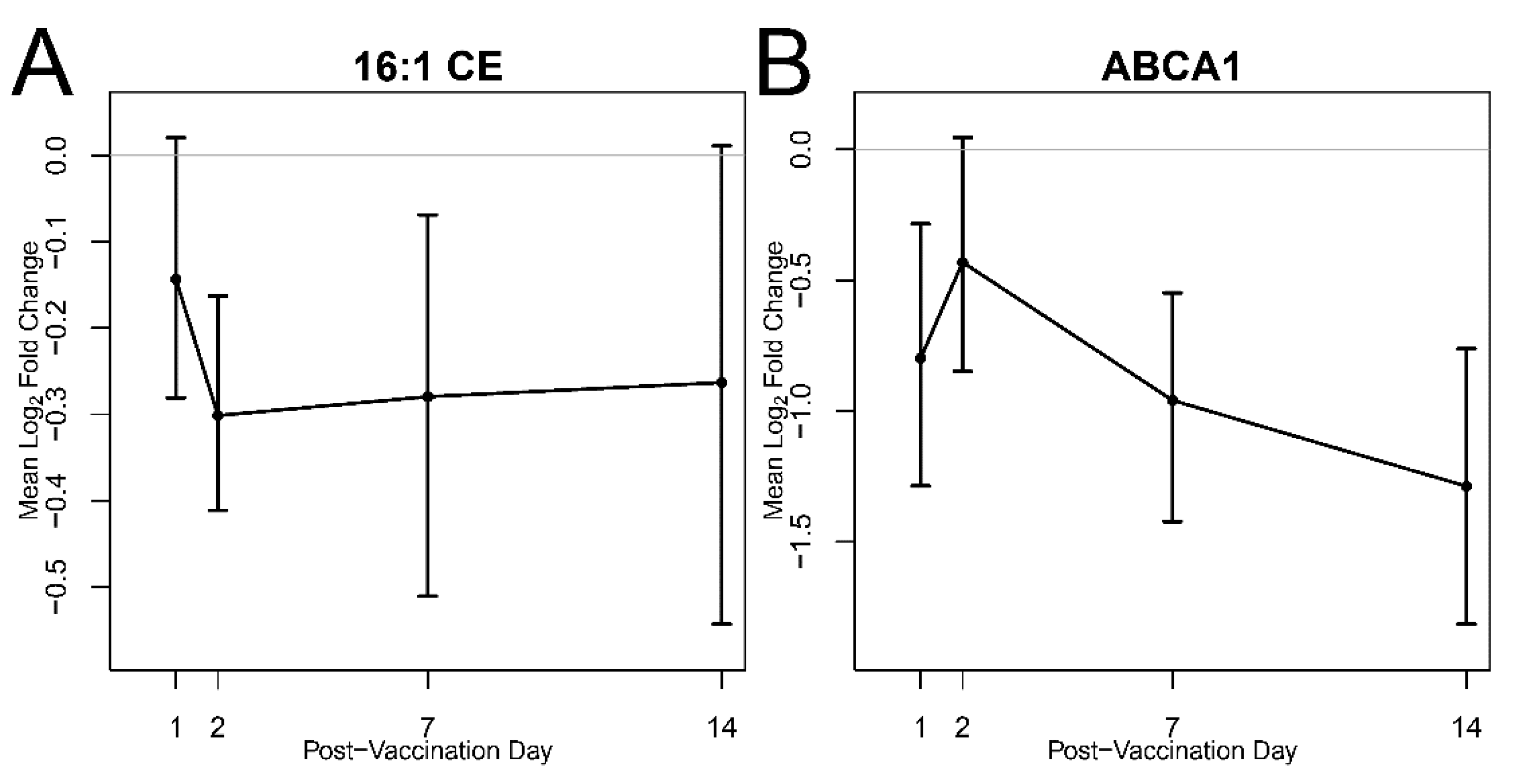
3. Results
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Clem, A.S. Fundamentals of Vaccine Immunology. J. Glob. Infect. Dis. 2011, 3, 73–78. [Google Scholar] [CrossRef] [PubMed]
- Querec, T.D.; Akondy, R.S.; Lee, E.K.; Cao, W.; Nakaya, H.I.; Teuwen, D.; Pirani, A.; Gernert, K.; Deng, J.; Marzolf, B.; et al. Systems biology approach predicts immunogenicity of the yellow fever vaccine in humans. Nat. Immunol. 2009, 10, 116–125. [Google Scholar] [CrossRef] [PubMed]
- Edupuganti, S.; Eidex, R.B.; Keyserling, H.; Akondy, R.S.; Lanciotti, R.; Orenstein, W.; del Rio, C.; Pan, Y.; Querec, T.; Lipman, H.; et al. A Randomized, Double-Blind, Controlled Trial of the 17D Yellow Fever VirusVaccine Given in Combination with Immune Globulin or Placebo: Comparative Viremia and Immunogenicity. Am. J. Trop. Med. Hyg. 2013, 88, 172–177. [Google Scholar] [CrossRef]
- Hagan, T.; Cortese, M.; Rouphael, N.; Boudreau, C.; Linde, C.; Maddur, M.S.; Das, J.; Wang, H.; Guthmiller, J.; Zheng, N.Y.; et al. Antibiotics-Driven Gut Microbiome Perturbation Alters Immunity to Vaccines in Humans. Cell 2019, 178, 1313–1328. [Google Scholar] [CrossRef] [PubMed]
- Itoe, M.A.; Sampaio, J.L.; Cabal, G.G.; Real, E.; Zuzarte-Luis, V.; March, S.; Bhatia, S.N.; Frischknecht, F.; Thiele, C.; Shevchenko, A.; et al. Host Cell Phosphatidylcholine Isa Key Mediator of Malaria Parasite Survival during Liver Stage Infection. Cell Host Microbe. 2014, 16, 778–786. [Google Scholar] [CrossRef]
- Wenk, M.R. Lipidomics of host-pathogen interactions. FEBS Lett. 2006, 580, 5541–5551. [Google Scholar] [CrossRef]
- Francis, E. Deer-fly fever, or Pahvant Valley Plague. A disease of man of hitherto unknown etiology. Public Health Rep. 1919, 34, 2061–2062. [Google Scholar] [CrossRef]
- Mulligan, M.J. Tularemia vaccine: Safety, reactogenicity, “Take” skin reactions, and antibody responses following vaccination with a new lot of the Francisella tularensis live vaccine strain—A phase 2 randomized clinical Trial. Vaccine 2017, 35, 4730–4737. [Google Scholar] [CrossRef]
- Mandell, G.L.; Bennett, J.E. Principles and Practices of Infectious Diseases, 4th ed.; Churchill Livingstone: London, UK, 1994. [Google Scholar]
- Chang, Y.-H.; Duong, D.M.; Goll, J.B.; Wood, D.C.; Jensen, T.L.; Yin, L.; Gelber, C.E.; Seyfried, N.T.; Anderson, E.; Natrajan, M.S.; et al. Proteomic Analysis of Human Immune Responses to Live-Attenuated Tularemia Vaccine. Vaccines, accepted.
- Goll, J.B.; Li, S.; Edwards, J.L.; Bosinger, S.; Jensen, T.L.; Wang, Y.; Hooper, W.F.; Gelber, C.E.; Sanders, K.L.; Anderson, E.J.; et al. Transcriptomic and metabolic responses to a live-attenuated Francisella tularensis vaccine. Vaccines, accepted.
- Bligh, E.G.; Dyer, W.J. A Rapid Method of Total Lipid Extraction and Purification. Can. J. Biochem. Physiol. 1959, 37, 911–917. [Google Scholar] [CrossRef]
- Han, X.; Gross, R.W. Shotgun Lipidomics: Electrospray Ionization Mass Spectrometric Analysis and Quantitation of Cellular Lipidomes Directly from Crud Extracts of Biological Samples. Mass Spectrom. Rev. 2005, 24, 367–412. [Google Scholar] [CrossRef]
- Gouveia-Figueira, S.; Späth, J.; Zivkovic, A.M.; Nording, M.L. Profiling the Oxylipin and Endocannabinoid Metabolome by UPLC-ESI-MS/MS in Human Plasma to Monitor Postprandial Inflammation. PLoS ONE 2015, 10, e0132042. [Google Scholar] [CrossRef]
- Yang, J.; Schmelzer, K.; Georgi, K.; Hammock, B.D. Quantitative Profiling Method for Oxylipin Metabolome by Liquid Chromatography Electrospray Ionization Tandem Mass Spectrometry. Anal. Chem. 2009, 81, 8085–8093. [Google Scholar] [CrossRef]
- Strassburg, K.; Huijbrechts, A.M.L.; Kortekaas, K.A.; Lindeman, J.H.; Pedersen, T.L.; Dane, A.; Berger, R.; Brenkman, A.; Hankemeier, T.; Duynhoven, J.v.; et al. Quantitative profiling of oxylipins through comprehensive LC-MS/MS analysis: Application in cardiac surgery. Anal. Bioanal. Chem. 2012, 404, 1413–1426. [Google Scholar] [CrossRef]
- Zoerner, A.A.; Gutzki, F.-M.; Batkai, S.; May, M.; Rakers, C.; Engeli, S.; Jordan, J.; Tsikas, D. Quantification of endocannabinoids in biological systems by chromatography and mass spectrometry: A comprehensive review from an analytical and biological perspective. Biochim. Biophys. Acta 2011, 1811, 706–723. [Google Scholar] [CrossRef] [PubMed]
- Blanksby, S.J.; Mitchell, T.W. Advances in mass spectrometry for lipidomics. Annu. Rev. Anal. Chem. 2010, 3, 433–465. [Google Scholar] [CrossRef] [PubMed]
- Quehenberger, O.; Armando, A.M.; Brown, A.H.; Milne, S.B.; Myers, D.S.; Merrill, A.H.; Bandyopadhyay, S.; Jones, K.N.; Kelly, S.; Shaner, R.L.; et al. Lipidomics reveals a remarkable diversity of lipids in human plasma. J. Lipid. Res. 2010, 51, 3299–3305. [Google Scholar] [CrossRef] [PubMed]
- Brugger, B. Lipidomics: Analysis of the Lipid Composition of Cells and Subcellular Organelles by Electrospray Ionization Mass Spectrometry. Annu. Rev. Biochem. 2014, 83, 79–98. [Google Scholar] [CrossRef]
- Pulfer, M.; Murphy, R.C. Electrospray Mass Spectrometry of Phospholipids. Mass Spectrom. Rev. 2003, 22, 332–364. [Google Scholar] [CrossRef]
- Han, X.; Yang, K.; Cheng, H.; Fikes, K.N.; Gross, R.W. Shotgun lipidomics of phosphoethanolamine-containing lipids in biological samples after one-step in situ derivatization. J. Lipid Res. 2005, 46, 1548–1560. [Google Scholar] [CrossRef]
- Bowden, J.A.; Albert, C.J.; Barnaby, O.S.; Ford, D.A. Analysis of cholesteryl esters and diacylglycerols using lithiated adducts and electrospray ionization-tandem mass spectrometry. Anal. Biochem. 2011, 417, 202–210. [Google Scholar] [CrossRef]
- Han, X. Characterization and Direct Quantitation of Ceramide Molecular Species from Lipid Extracts of Biological Samples by Electrospray Ionization Tandem Mass Spectrometry. Anal. Biochem. 2002, 302, 199–212. [Google Scholar] [CrossRef] [PubMed]
- Gabbs, M.; Leng, S.; Devassy, J.G.; Monirujjaman, M.; Aukema, H.M. Advances in Our Understanding of Oxylipins Derived from Dietary PUFAs. Adv. Nutr. 2015, 6, 513–540. [Google Scholar] [CrossRef] [PubMed]
- Hastie, T.; Tibshirani, R.; Narasimhan, B.; Chu, G. Impute: Imputation for microarray data. Bioinformatics 2001, 17, 520–525. [Google Scholar]
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2013. [Google Scholar]
- Gentleman, R.C.; Carey, V.J.; Bates, D.M.; Bolstad, B.; Dettling, M.; Dudoit, S.; Ellis, B.; Gautier, L.; Ge, Y.; Gentry, J.; et al. Bioconductor: Open software development for computational biology and bioinformatics. Genome Biol. 2004, 5, R80. [Google Scholar] [CrossRef] [PubMed]
- Quehenberger, O. The Human Plasma Lipidome. N. Engl. J. Med. 2011, 365, 1812–1823. [Google Scholar] [CrossRef] [PubMed]
- McNamara, D.J.; Kolb, R.; Parker, T.S.; Batwin, H.; Samuel, P.; Brown, C.D.; Ahrens, E.H., Jr. Heterogeneity of cholesterol homeostasis in man. Response to changes in dietary fat quality and cholesterol quantity. J. Clin. Investig. 1987, 79, 1729–1739. [Google Scholar] [CrossRef]
- Nestel, P.; Poyser, A. Changes in cholesterol synthesis and excretion when cholesterol intake is increased. Metabolism 1976, 25, 1591–1599. [Google Scholar] [CrossRef]
- Alphonse, P.A.S.; Jones, P.J.H. Revisiting Human Cholesterol Synthesis and Absorption: The Reciprocity Paradigm and its Key Regulators. Lipids. 2016, 51, 519–536. [Google Scholar] [CrossRef]
- Tall, A.R.; Yvan-Charvet, L. Cholesterol, inflammation and innate immunity. Nat. Rev. Immunol. 2015, 15, 104–116. [Google Scholar] [CrossRef]
- Gomez-Muñoz, A.; Presa, N.; Gomez-Larrauri, A.; Rivera, I.; Trueba, M.; Ordoñez, M. Control of inflammatory responses by ceramide, sphingosine 1-phosphate and ceramide 1-phosphate. Prog. Lipid Res. 2016, 61, 51–62. [Google Scholar] [CrossRef]
- Powell, W.S.; Rokach, J. The eosinophil chemoattractant 5-oxo-ETE and the OXE receptor. Prog. Lipid Res. 2013, 52, 651–665. [Google Scholar] [CrossRef] [PubMed]
- Gail, G.E.; Rokach, J.; Powell, W.S. 5-Oxo-ETE and the OXE receptor. Prostaglandins Other Lipid Mediat. 2009, 89, 98–104. [Google Scholar]
- Szanto, A.; Nagy, L. The many faces of PPARg: Anti-inflammatory by any means? Immunobiology 2008, 213, 789–803. [Google Scholar] [CrossRef] [PubMed]
- Kersten, S.; Desvergne, B.; Wahli, W. Roles of PPARs in health and disease. Nature 2000, 405, 421–424. [Google Scholar] [CrossRef] [PubMed]
- Szanto, A.; Balint, B.L.; Nagy, Z.S.; Barta, E.; Dezso, B.; Pap, A.; Szeles, L.; Poliska, S.; Oros, M.; Evans, R.M.; et al. STAT6 transcription factor is a facilitator of the nuclear receptor PPARgamma-regulated gene expression in macrophages and dendritic cells. Immunity 2010, 33, 699–712. [Google Scholar] [CrossRef] [PubMed]
- Jourdan, T.; Godlewski, G.; Cinar, R.; Bertola, A.; Szanda, G.; Liu, J.; Tam, J.; Han, T.; Mukhopadhyay, B.; Skarulis, M.C.; et al. Activation of the Nlrp3 inflammasome in infiltratingmacrophages by endocannabinoids mediates beta cell loss in type 2 diabetes. Nat. Med. 2013, 19, 1132–1140. [Google Scholar] [CrossRef]
- Liu, J.; Wang, L.; Harvey-White, J.; Osei-Hyiaman, D.; Razdan, R.; Gong, Q.; Chan, A.C.; Zhou, Z.; Huang, B.X.; Kim, H.-Y.; et al. A biosynthetic pathway for anandamide. Proc. Natl. Acad. Sci. USA 2006, 103, 13345–13350. [Google Scholar] [CrossRef]
- Adaptive Immune Responses to Booster Vaccination against Yellow Fever Virus Are Much Reduced Compared to Those after Primary Vaccination. Available online: https://www.nature.com/articles/s41598-017-00798-1 (accessed on 10 July 2020).
- Vida, M.; Serrano, A.; Romero-Cuevas, M.; Pavon, F.J.; Gonzalez-Rodriguez, A.; Gavito, A.L.; Cuesta, A.L.; Angela, M.A.; Valverde, A.M.; de Fonseca, F.R.; et al. IL-6 cooperates with peroxisome proliferator-activatedreceptor-a-ligands to induce liver fatty acid binding protein (LFABP) up-regulation. Liver Int. 2013, 33, 1019–1028. [Google Scholar] [CrossRef]
- Yang, L.; Guo, H.; Li, Y.; Meng, X.; Ya, L.; Zhang, D.; Wu, S.; Zhou, H.; Peng, H.; Xie, Q.; et al. Oleoylethanolamide exerts anti-inflammatory effects on LPS-induced THP-1 cells by enhancing PPARα signaling and inhibiting the NF-κB and ERK1/2/AP-1/STAT3 pathways. Sci. Rep. 2016, 6, 34611. [Google Scholar] [CrossRef]
- Cucchi, D.; Camacho-Muñoz, D.; Certo, M.; Niven, J.; Smith, J.; Nicolaou, A.; Mauro, C. Omega-3 polyunsaturated fatty acids impinge on CD41 T cell motility and adipose tissue distribution via direct and lipid mediator-dependent effects. Cardiovasc. Res. 2019, 116, 1006–1020. [Google Scholar]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Lipid Class | Class Abbrev. | Ionization Mode | Characteristic Scan | Lipid IDs | Collision Energy |
|---|---|---|---|---|---|
| Phosphatidylethanolamine | PE | Negative | NL222 | 35 | −30 |
| Phosphatidylcholine | PC | Negative | NL50 | 42 | −30 |
| Sphingomyelin | SM | Negative | NL50 | 4 | −24 |
| Phosphatidylinositol | PI | Negative | Prec241 | 9 | −30 |
| Phosphatidylserine | PS | Negative | NL87 | 1 | −30 |
| Cholesteryl Esters | CE | Positive | NL368 | 14 | 25 |
| Ceramide | Cer | Negative | NL256 | 11 | −32 |
| Lipid Species | Abbreviation | m/z | Transition |
|---|---|---|---|
| Oleoylethanolamide | OEA | 325.5 | 325 < 62 |
| Arachidonoylethanolamine | AEA | 347.5 | 347 < 62 |
| Prostaglandin E2 Ethanolamide | PGE2 Ethanolamide | 395.5 | 378 < 62 |
| Prostaglandin F2a Ethanolamide | PGF2a Ethanolamide | 397.5 | 380 < 62 |
| 12(13)-EpOME/13-HODE | |||
| 12(13)epoxy-9-octadecenoic acid | 12(13)-EpOME | 296.5 | 296 < 168 |
| 13-hydroxy-9,11-octadecadienoic acid | 13-HODE | 296.5 | 296 < 168 |
| 9,10-dihydroxy-12-octadecenoic acid | 9,10-DiHOME | 314.5 | 314 < 172 |
| Thromboxane B2 | TXB2 | 370.5 | 370 < 147 |
| 12-hydroxy-5,8,10,14-eicosatetraenoic acid | 12-HETE | 320.5 | 320 < 87 |
| 9-HETE/11(12)-EET | |||
| 9-hydroxy-5,7,11,14-eicosatetraenoic acid | 9-HETE | 320.5 | 320 < 167 |
| 11(12)-epoxy-5,8,14-eicosatrienoic acid | 11(12)-EET | 320.5 | 320 < 167 |
| 20-hydroxy-5,8,11,14-eicosatetraenoic acid | 20-HETE | 320.5 | 320 < 289 |
| 5-hydroxy-6,8,11,14-eicosatetraenoic acid | 5-HETE | 320.5 | 320 < 301 |
| 8(9)-epoxy-5,11,14-eicosatrienoic acid | 8(9)-EET | 320.5 | 320 < 69 |
| 14(15)-epoxy-5,8,11-eicosatrienoic acid | 14(15)-EET | 320.5 | 320 < 220 |
| Summed DHET Species | |||
| 14,15-dihydroxy-5,8,11-eicosatrienoic acid | 14,15-DHET | 338.5 | 338 < 256, 170,184,82 |
| 11,12-dihydroxy-5,8,14-eicosatrienoic acid | 11,12-DHET | 338.5 | 338 < 256, 170,184,82 |
| 8,9-dihydroxy-5,11,14-eicosatrienoic acid | 8,9-DHET | 338.5 | 338 < 256, 170,184,82 |
| 5,6-dihydroxy-8,11,14-eicosatrienoic acid | 5,6-DHET | 338.5 | 338 < 256, 170,184,82 |
| Lipid Name | Lipid ID | Lipid Class | Day | Fold Change | t-Statistic | p-Value |
|---|---|---|---|---|---|---|
| 16:1 LPC | TULIPID017 | PC | Day 1 | 0.709 | −2.7 | 0.0137 |
| 20:3 LPC | TULIPID024 | PC | Day 1 | 0.74 | −5.5 | 0.002 |
| 36:3 PE | TULIPID103 | PE | Day 1 | 1.238 | 1.8 | 0.0469 |
| p40:6 PE | TULIPID119 | PE | Day 1 | 1.214 | 2.4 | 0.0449 |
| 16:1 CE | TULIPID078 | CE | Day 2 | 0.811 | −4.7 | 0.0039 |
| 16:1 LPC | TULIPID017 | PC | Day 2 | 0.597 | −2.9 | 0.0234 |
| 20:3 LPC | TULIPID024 | PC | Day 2 | 0.62 | −3.8 | 0.0078 |
| 22:3 CE | TULIPID089 | CE | Day 2 | 0.376 | −2.6 | 0.0371 |
| 34:1 PI | TULIPID058 | PI | Day 2 | 0.388 | −2.7 | 0.0254 |
| 34:2 PE | TULIPID099 | PE | Day 2 | 0.797 | −3.4 | 0.0098 |
| AEA | TULIPID002 | OXY | Day 2 | 0.686 | −2.7 | 0.0312 |
| OEA | TULIPID001 | OXY | Day 2 | 0.682 | −3.0 | 0.0156 |
| 16:1 CE | TULIPID078 | CE | Day 7 | 0.824 | −2.2 | 0.0371 |
| 16:1 LPC | TULIPID017 | PC | Day 7 | 0.534 | −2.6 | 0.0312 |
| 20:3 LPC | TULIPID024 | PC | Day 7 | 0.598 | −3.0 | 0.0195 |
| 22:0 Cer | TULIPID072 | Cer | Day 7 | 0.807 | −2.9 | 0.0254 |
| 38:3 PI | TULIPID064 | PI | Day 7 | 0.491 | −2.4 | 0.0469 |
| 5-HETE | TULIPID012 | OXY | Day 7 | 0.386 | −2.7 | 0.0312 |
| Summed DHET Species | TULIPID007 | OXY | Day 7 | 1.969 | 3.1 | 0.0215 |
| 5-HETE | TULIPID012 | OXY | Day 14 | 0.674 | −2.3 | 0.0488 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Maner-Smith, K.M.; Goll, J.B.; Khadka, M.; Jensen, T.L.; Colucci, J.K.; Gelber, C.E.; Albert, C.J.; Bosinger, S.E.; Franke, J.D.; Natrajan, M.; et al. Alterations in the Human Plasma Lipidome in Response to Tularemia Vaccination. Vaccines 2020, 8, 414. https://doi.org/10.3390/vaccines8030414
Maner-Smith KM, Goll JB, Khadka M, Jensen TL, Colucci JK, Gelber CE, Albert CJ, Bosinger SE, Franke JD, Natrajan M, et al. Alterations in the Human Plasma Lipidome in Response to Tularemia Vaccination. Vaccines. 2020; 8(3):414. https://doi.org/10.3390/vaccines8030414
Chicago/Turabian StyleManer-Smith, Kristal M., Johannes B. Goll, Manoj Khadka, Travis L. Jensen, Jennifer K. Colucci, Casey E. Gelber, Carolyn J. Albert, Steven E. Bosinger, Jacob D. Franke, Muktha Natrajan, and et al. 2020. "Alterations in the Human Plasma Lipidome in Response to Tularemia Vaccination" Vaccines 8, no. 3: 414. https://doi.org/10.3390/vaccines8030414
APA StyleManer-Smith, K. M., Goll, J. B., Khadka, M., Jensen, T. L., Colucci, J. K., Gelber, C. E., Albert, C. J., Bosinger, S. E., Franke, J. D., Natrajan, M., Rouphael, N., Johnson, R. A., Sanz, P., Anderson, E. J., Hoft, D. F., Mulligan, M. J., Ford, D. A., & Ortlund, E. A. (2020). Alterations in the Human Plasma Lipidome in Response to Tularemia Vaccination. Vaccines, 8(3), 414. https://doi.org/10.3390/vaccines8030414