

The Stability and Efficency of CPB Cells Were Acclimated for Virus Proliferation
Abstract
1. Introduction
2. Material and Methods
2.1. Cells and Virus Strains
2.2. Cells Acclimation
2.3. Morphological Characteristics of CPB Cells
2.4. The Growth Characteristics of CPB Cells
2.5. Transfection Efficiency
2.6. Virus Susceptibility
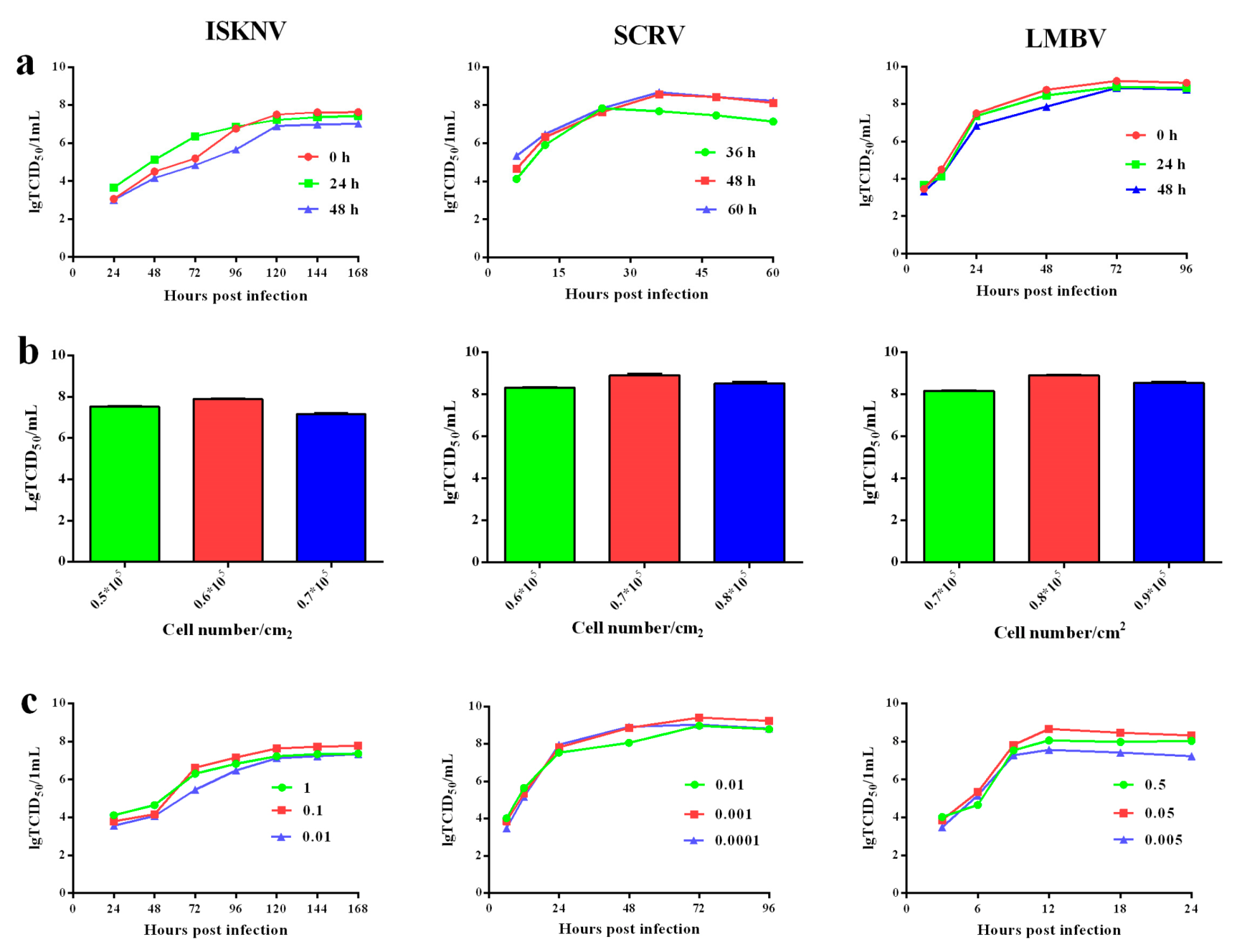
2.7. Virus Multiplication Conditions
2.8. The Time and Economic Cost of Viral Multiplication
2.9. Statistical Analyses
3. Results
3.1. The Passage of CPB Cells
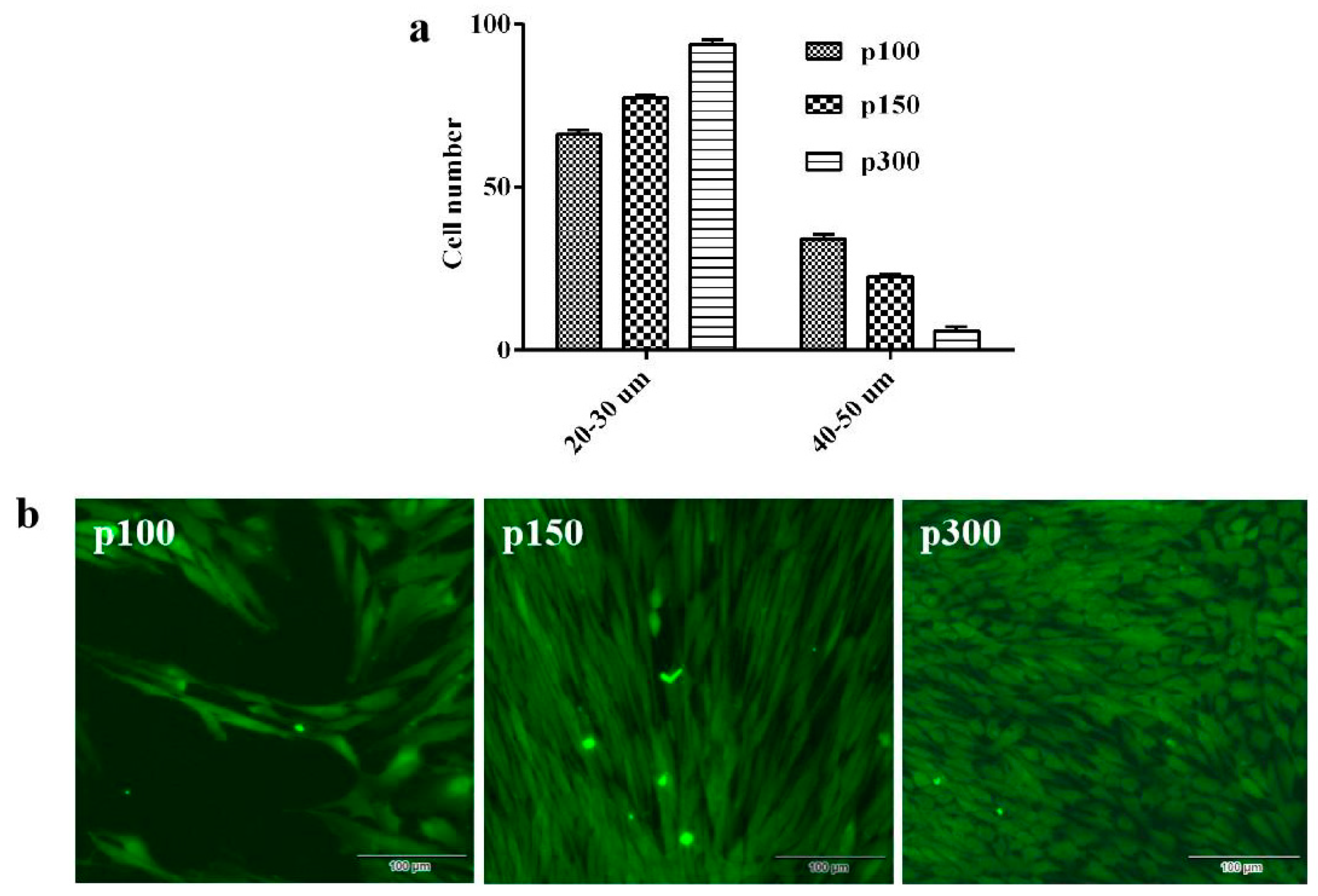
3.2. Identification of Morphological Features
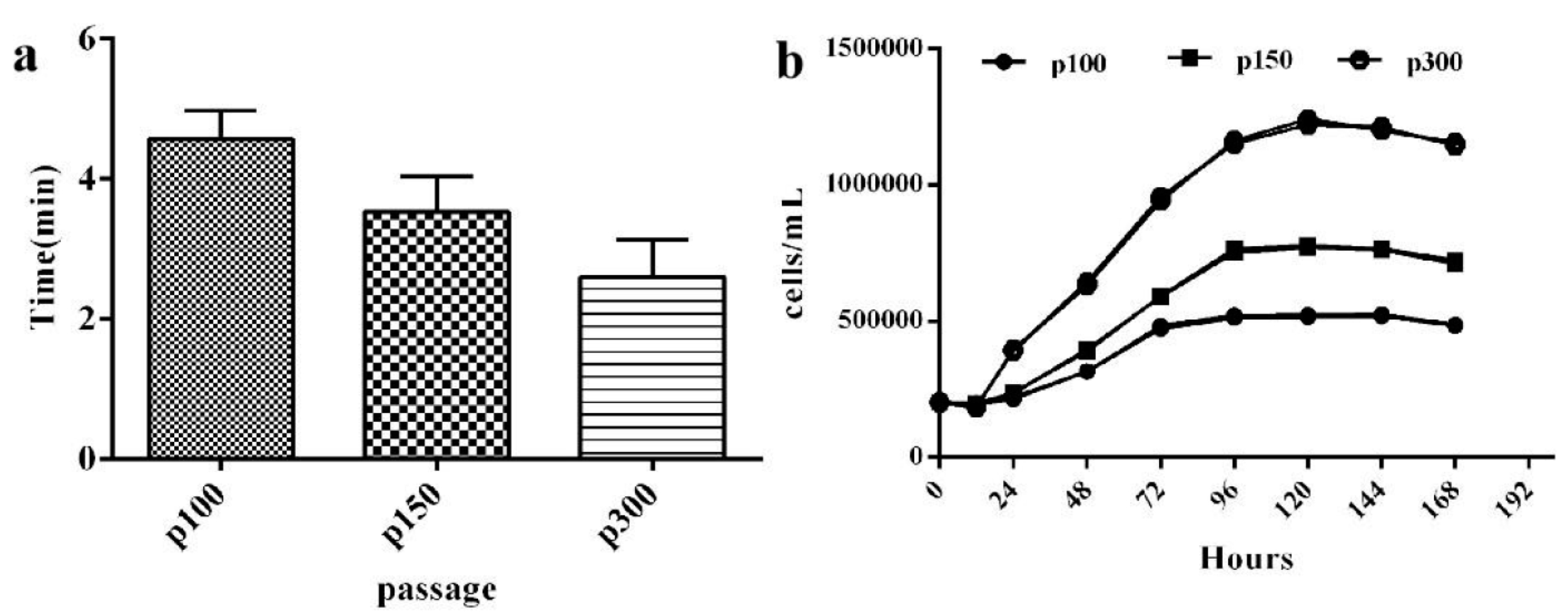
3.3. The Growth Characteristics of CPB Cells
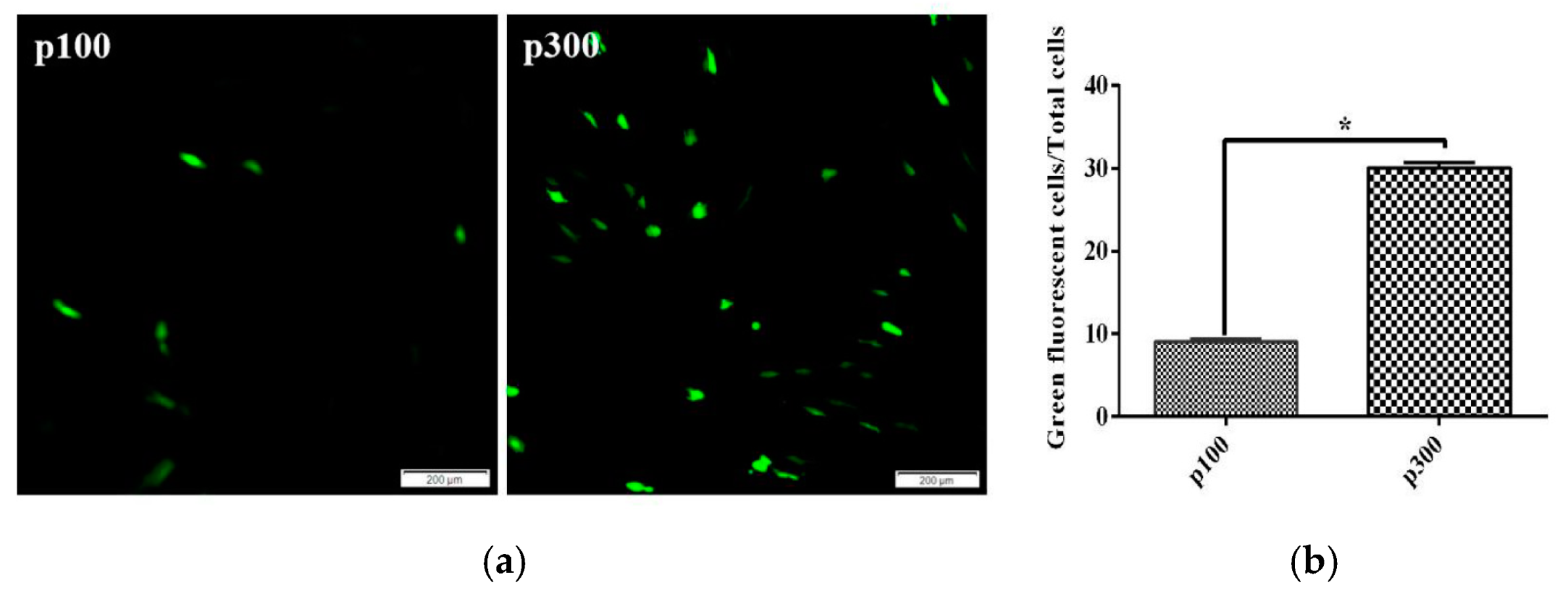
3.4. Transfection Efficiency
3.5. Virus Susceptibility of CPB p300
3.6. The Optimal Conditions of Viral Multiplication in CPBp300 Cells
3.7. The Time and Economic Cost of Viral Multiplication
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Liang, X.F.; Oku, H.; Ogata, H.Y.; Liu, J.; He, X. Weaning Chinese perch Siniperca chuatsi (Basilewsky) onto artificial diets based upon its specific sensory modality in feeding. Aquac. Res. 2001, 32, 76–82. [Google Scholar] [CrossRef]
- Xie, S.; Yin, P.; Tian, L.; Yu, Y.; Liu, Y.; Niu, J. Dietary Supplementation of Astaxanthin Improved the Growth Performance, Antioxidant Ability and Immune Response of Juvenile Largemouth Bass (Micropterus salmoides) Fed High-Fat Diet. Mar. Drugs 2020, 18, 642. [Google Scholar] [CrossRef] [PubMed]
- Lin, Q.; Fu, X.; Li, N.; Wan, Q.; Chen, W.; Huang, Y.; Huang, Z.; Li, J.; Zhao, L.; Lin, L. Co-infections of infectious spleen and kidney necrosis virus and Siniperca chuatsi rhabdovirus in Chinese perch (Siniperca chuatsi). Microb. Pathog. 2017, 111, 422–430. [Google Scholar] [CrossRef] [PubMed]
- Wang, P.; Huo, X.; Zhao, F.; Liu, Q.; Tian, Q.; Yang, C.; Su, J. Vitamin D3 can effectively and rapidly clear largemouth bass ranavirus by immunoregulation. Fish Shellfish Immunol. 2023, 143, 109213. [Google Scholar] [CrossRef] [PubMed]
- Talbi, C.; Cabon, J.; Baud, M.; Bourjaily, M.; de Boisséson, C.; Castric, J.; Bigarré, L. Genetic diversity of perch rhabdoviruses isolates based on the nucleoprotein and glycoprotein genes. Arch. Virol. 2011, 156, 2133–2144. [Google Scholar] [CrossRef]
- Ma, D.; Deng, G.; Bai, J.; Li, S.; Yu, L.; Quan, Y.; Yang, X.; Jiang, X.; Zhu, Z.; Ye, X. A Strain of Siniperca chuatsi Rhabdovirus Causes High Mortality among Cultured Largemouth Bass in South China. J. Aquat. Anim. Health 2013, 25, 197–204. [Google Scholar] [CrossRef] [PubMed]
- Xu, Z.; Liao, J.; Zhang, D.; Liu, S.; Zhang, L.; Kang, S.; Xu, L.; Chen, H.; Peng, W.; Zhou, S.; et al. Isolation, Characterization, and Transcriptome Analysis of an ISKNV-Like Virus from Largemouth Bass. Viruses 2023, 15, 398. [Google Scholar] [CrossRef] [PubMed]
- Fu, X.; Li, N.; Liu, L.; Lin, Q.; Wang, F.; Lai, Y.; Jiang, H.; Pan, H.; Shi, C.; Wu, S. Genotype and host range analysis of infectious spleen and kidney necrosis virus (ISKNV). Virus Genes 2011, 42, 97–109. [Google Scholar] [CrossRef]
- Neal, J.W.; Eggleton, M.A.; Goodwin, A.E. The effects of largmouth bass virus on a quality largemouth bass population arkansas. J. Wildl. Dis. 2009, 45, 766–771. [Google Scholar] [CrossRef]
- Liu, X.; Zhang, Y.; Zhang, Z.; An, Z.; Zhang, X.; Vakharia, V.N.; Lin, L. Isolation, identification and the pathogenicity characterization of a Santee-Cooper ranavirus and its activation on immune responses in juvenile largemouth bass (Micropterus salmoides). Fish Shellfish Immunol. 2023, 135, 108641. [Google Scholar] [CrossRef]
- Ahmed, R.; Burton, D.R. Viral vaccines: Past successes and future challenges. Curr. Opin. Virol. 2013, 3, 307–308. [Google Scholar] [CrossRef]
- Adams, A. Progress, challenges and opportunities in fish vaccine development. Fish Shellfish Immunol. 2019, 90, 210–214. [Google Scholar] [CrossRef]
- Dai, X.; Zhang, X.; Ostrikov, K.; Abrahamyan, L. Host receptors: The key to establishing cells with broad viral tropism for vaccine production. Crit. Rev. Microbiol. 2020, 46, 147–168. [Google Scholar] [CrossRef]
- Aubrit, F.; Perugi, F.; Léon, A.; Guéhenneux, F.; Champion-Arnaud, P.; Lahmar, M.; Schwamborn, K. Cell substrates for the production of viral vaccines. Vaccine 2015, 33, 5905–5912. [Google Scholar] [CrossRef]
- Fu, X.; Li, N.; Lai, Y.; Luo, X.; Wang, Y.; Shi, C.; Huang, Z.; Wu, S.; Su, J. A novel fish cell line derived from the brain of Chinese perch Siniperca chuatsi: Development and characterization. J. Fish Biol. 2015, 86, 32–45. [Google Scholar] [CrossRef]
- Niu, Y.; Zhang, P.; Wang, L.; Li, N.; Lin, Q.; Liu, L.; Liang, H.; Huang, Z.; Fu, X. Development of double-antibody sandwich ELISA for rapidly quantitative detection of antigen concentration in inactivated SCRV vaccine. Aquaculture 2020, 520, 734671. [Google Scholar] [CrossRef]
- Lijuan, Z.; Ningqiu, L.; Qiang, L.; Lihui, L.; Hongru, L.; Zhibin, H.; Xiaozhe, F. An avirulent Micropterus salmoides rhabdovirus vaccine candidate protects Chinese perch against rhabdovirus infection. Fish Shellfish Immunol. 2018, 77, 474–480. [Google Scholar] [CrossRef] [PubMed]
- Luo, X.; Niu, Y.; Fu, X.; Lin, Q.; Liang, H.; Liu, L.; Li, N. Large-Scale Microcarrier Culture of Chinese Perch Brain Cell for Viral Vaccine Production in a Stirred Bioreactor. Vaccines 2021, 9, 1003. [Google Scholar] [CrossRef] [PubMed]
- Chen, C.; Wang, Z.; Sun, Z.; Li, W.; Dimitrov, D.S. Development of an efficient method for selection of stable cell pools for protein expression and surface display with Expi293F cells. Cell Biochem. Funct. 2023, 41, 355–364. [Google Scholar] [CrossRef] [PubMed]
- Baust, J.M.; Buehring, G.C.; Campbell, L.; Elmore, E.; Harbell, J.; Nims, R.W.; Price, P.; Reid, Y.A.; Simione, F. Best practices in cell culture an overview. Animal 2017, 2017, 669–672. [Google Scholar] [CrossRef] [PubMed]
- Sharfstein, S.T. Advances in Cell Culture Process Development: Tools and Techniques for Improving Cell Line Development and Process Optimization. Biotechnol. Prog. 2008, 24, 727–734. [Google Scholar] [CrossRef]
- Siddiqui, M.S.I.; Islam, M.R.; Chowdhury, E.H. Growth kinetics of a Vero cells adapted Bangladeshi strain of peste des petits ruminants (PPR) virus in cell culture. Arch. Microbiol. 2021, 203, 1587–1593. [Google Scholar] [CrossRef]
- Genzel, Y.; Behrendt, I.; König, S.; Sann, H.; Reichl, U. Metabolism of MDCK cells during cell growth and influenza virus production in large-scale microcarrier culture. Vaccine 2004, 22, 2202–2208. [Google Scholar] [CrossRef]
- Jin, Y.; Bergmann, S.M.; Mai, Q.; Yang, Y.; Liu, W.; Sun, D.; Chen, Y.; Yu, Y.; Liu, Y.; Cai, W.; et al. Simultaneous Isolation and Identification of Largemouth Bass Virus and Rhabdovirus from Moribund Largemouth Bass (Micropterus salmoides). Viruses 2022, 14, 1643. [Google Scholar] [CrossRef]
- Cooper Robbins, S.C.; Ward, K.; Skinner, S.R. School-based vaccination: A systematic review of process evaluations. Vaccine 2011, 29, 9588–9599. [Google Scholar] [CrossRef]
- Razali, S.A.; Shamsir, M.S.; Ishak, N.F.; Low, C.-F.; Azemin, W.-A. Riding the wave of innovation: Immunoinformatics in fish disease control. PeerJ 2023, 11, e16419. [Google Scholar] [CrossRef] [PubMed]
- Goh, K.W.; Abdul Kari, Z.; Wee, W.; Zakaria, N.N.A.; Rahman, M.M.; Kabir, M.A.; Abdul Hamid, N.K.; Tahiluddin, A.B.; Kamarudin, A.S.; Téllez–Isaías, G.; et al. Exploring the roles of phytobiotics in relieving the impacts of Edwardsiella tarda infection on fish: A mini-review. Front. Vet. Sci. 2023, 10, 1149514. [Google Scholar] [CrossRef] [PubMed]
- Genzel, Y. Designing cell lines for viral vaccine production: Where do we stand? Biotechnol. J. 2015, 10, 728–740. [Google Scholar] [CrossRef] [PubMed]
- Kiesslich, S.; Kamen, A.A. Vero cell upstream bioprocess development for the production of viral vectors and vaccines. Biotechnol. Adv. 2020, 44, 107608. [Google Scholar] [CrossRef]
- Yun, S.-I.; Lee, Y.-M. Japanese encephalitis. Hum. Vaccines Immunother. 2013, 10, 263–279. [Google Scholar] [CrossRef]
- Hallman, W.K.; Hallman, W.K.; Hallman, E.E. Cell-based, cell-cultured, cell-cultivated, cultured, or cultivated. What is the best name for meat, poultry, and seafood made directly from the cells of animals? npj Sci. Food 2023, 7, 62. [Google Scholar] [CrossRef] [PubMed]
- Colosimo, A.; Goncz, K.; Holmes, A.; Kunzelmann, K.; Novelli, G.; Malone, R.; Bennett, M.; Gruenert, D. Transfer and Expression of Foreign Genes in Mammalian Cells. BioTechniques 2000, 2000, 314–331. [Google Scholar] [CrossRef] [PubMed]
- Hoenen, T.; Groseth, A. Virus–Host Cell Interactions. Cells 2022, 11, 804. [Google Scholar] [CrossRef] [PubMed]
- Ferris, N.P.; Hutchings, G.H.; Moulsdale, H.J.; Golding, J.; Clarke, J.B. Sensitivity of primary cells immortalised by oncogene transfection for the detection and isolation of foot-and-mouth disease and swine vesicular disease viruses. Vet. Microbiol. 2002, 2002, 307–316. [Google Scholar] [CrossRef]
- Petiot, E.; Jacob, D.; Lanthier, S.; Lohr, V.; Ansorge, S.; Kamen, A.A. Metabolic and Kinetic analyses of influenza production in perfusion HEK293 cell culture. BMC Biotechnol. 2011, 11, 84. [Google Scholar] [CrossRef]
- Dill, V.; Zimmer, A.; Beer, M.; Eschbaumer, M. Investigation of cell culture conditions for optimal foot-and-mouth disease virus production. BMC Biotechnol. 2019, 19, 33. [Google Scholar] [CrossRef]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| T | 0 h | 24 h | 48 h | 72 h | 96 h | 120 h | 144 h | 168 h | Doubling | Multiplication |
|---|---|---|---|---|---|---|---|---|---|---|
| p100 | 2 × 105 | 2.17 × 105 | 3.15 × 105 | 4.77 × 105 | 5.16 × 105 | 5.19 × 105 | 5.20 × 105 | 4.85 × 105 | 60 h | 2.616 |
| p150 | 2 × 105 | 2.36 × 105 | 3.90 × 105 | 5.89 × 105 | 7.58 × 105 | 7.74 × 105 | 7.63 × 105 | 7.16 × 105 | 48 h | 3.61 |
| p300 | 2 × 105 | 3.92 × 105 | 6.36 × 105 | 9.48 × 105 | 1.16 × 106 | 1.23 × 106 | 1.20 × 106 | 1.15 × 106 | 24 h | 6.2 |
| Viruses | p100(TCID50/y) | p300(TCID50/y) | p100(TCID50/d) | p300(TCID50/d) |
|---|---|---|---|---|
| ISKNV | (2.25 ± 0.15) × 107 | (4.0 ± 0.15) × 107 | (2.52 ± 0.35) × 107 | (4.73 ± 0.22) × 107 |
| SCRV | (1.17 ± 0.16) × 108 | (2.24 ± 0.25) × 109 | (2.01 ± 0.35) × 108 | (4.84 ± 0.51) × 109 |
| LMBV | (2.00 ± 0.15) × 108 | (2.64 ± 0.36) × 109 | (3.77 ± 0.32) × 108 | (5.43 ± 0.35) × 109 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Niu, Y.; Ma, S.; Liang, H.; Fu, X.; Ma, B.; Lin, Q.; Luo, X.; Li, N. The Stability and Efficency of CPB Cells Were Acclimated for Virus Proliferation. Vaccines 2024, 12, 220. https://doi.org/10.3390/vaccines12030220
Niu Y, Ma S, Liang H, Fu X, Ma B, Lin Q, Luo X, Li N. The Stability and Efficency of CPB Cells Were Acclimated for Virus Proliferation. Vaccines. 2024; 12(3):220. https://doi.org/10.3390/vaccines12030220
Chicago/Turabian StyleNiu, Yinjie, Saiya Ma, Hongru Liang, Xiaozhe Fu, Baofu Ma, Qiang Lin, Xia Luo, and Ningqiu Li. 2024. "The Stability and Efficency of CPB Cells Were Acclimated for Virus Proliferation" Vaccines 12, no. 3: 220. https://doi.org/10.3390/vaccines12030220
APA StyleNiu, Y., Ma, S., Liang, H., Fu, X., Ma, B., Lin, Q., Luo, X., & Li, N. (2024). The Stability and Efficency of CPB Cells Were Acclimated for Virus Proliferation. Vaccines, 12(3), 220. https://doi.org/10.3390/vaccines12030220