

Expression of E. coli FimH Enhances Trafficking of an Orally Delivered Lactobacillus acidophilus Vaccine to Immune Inductive Sites via Antigen-Presenting Cells

Abstract
1. Introduction
2. Materials and Methods
2.1. FimH Plasmid Construction and Lactobacillus acidophilus Transformation
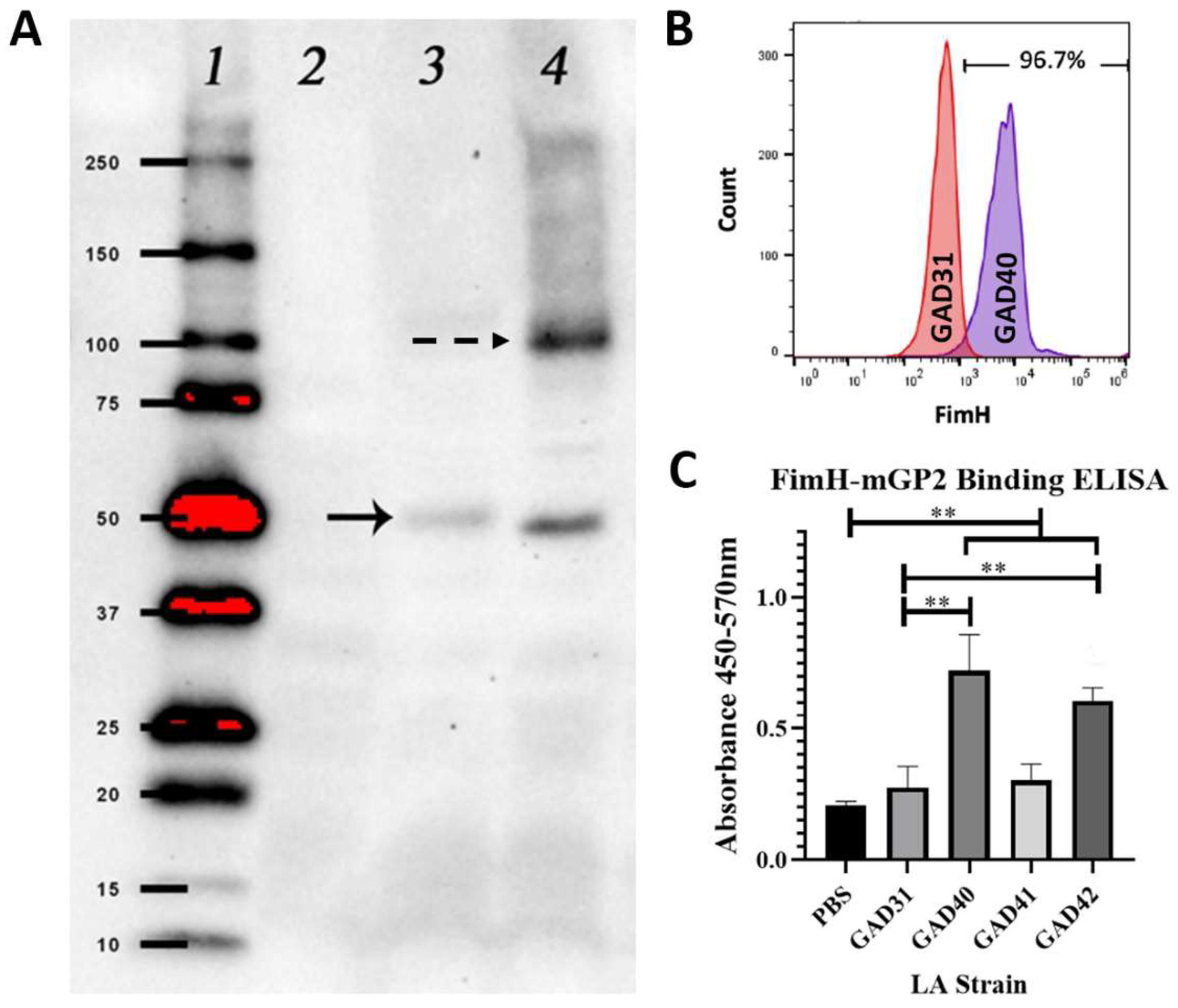
2.2. N-Term FimH Western Blot
2.3. N-Term FimH rLA Surface Expression Flow Cytometry
2.4. Murine GP2 Expression and Purification
2.5. rLA FimH-Mouse GP2-Binding Assay
2.6. Ethics Statement and Mouse Usage
2.7. Intestinal Loop Peyer’s Patch rLA Uptake and Mesenteric Lymph Node Trafficking
2.8. Mesenteric Lymph Node Antigen-Presenting Cell Flow Cytometry
2.9. N-Term FimH TLR4 Activation Assay
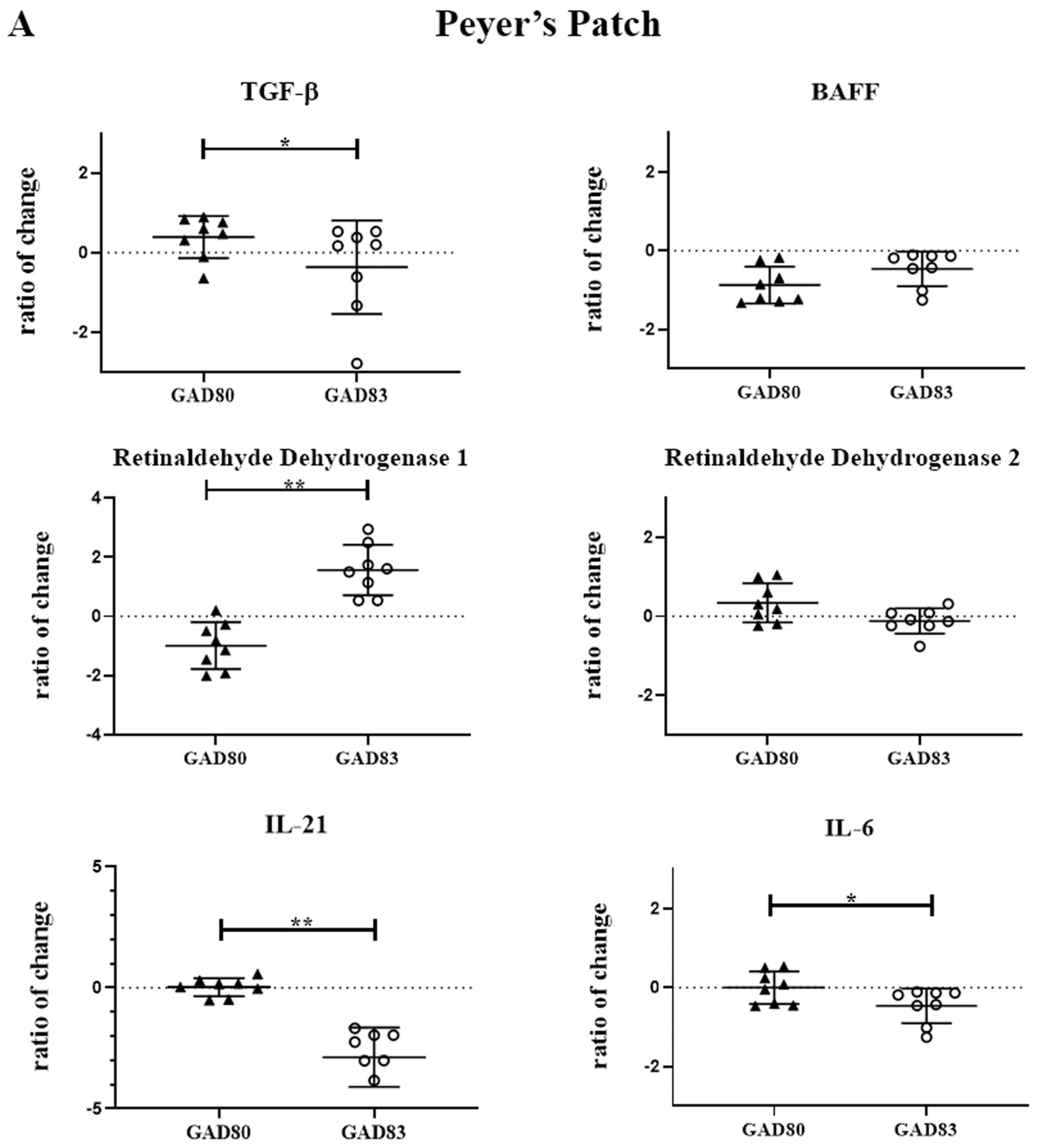
2.10. Peyer’s Patch and Mesenteric Lymph Node Quantitative Real-Time PCR Cytokine Evaluation
2.11. Statistics
3. Results
3.1. The N-Terminal Domain of E. coli FimH Can Be Detected on the Surface of Lactobacillus acidophilus by Flow Cytometry and Binds Murine GP2 In Vitro
3.2. N-Term FimH Increases Trafficking of rLA to Mesenteric Lymph Nodes, Which Is Mediated by Macrophages and Dendritic Cells
3.3. N-Term FimH Displayed on Surface of rLA Does Not Activate TLR4
3.4. N-Term FimH Increases Expression of Retinaldehyde Dehydrogenase 1 and Decreases Expression of IL-21 in Peyer’s Patches and Mesenteric Lymph Nodes
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Lavelle, E.C.; Ward, R.W. Mucosal vaccines—Fortifying the frontiers. Nat. Rev. Immunol. 2022, 22, 236–250. [Google Scholar] [CrossRef]
- Lynn, D.J.; Benson, S.C.; Lynn, M.A.; Pulendran, B. Modulation of immune responses to vaccination by the microbiota: Implications and potential mechanisms. Nat. Rev. Immunol. 2022, 22, 33–46. [Google Scholar] [CrossRef]
- Kajikawa, A.; Masuda, K.; Katoh, M.; Igimi, S. Adjuvant effects for oral immunization provided by recombinant Lactobacillus casei secreting biologically active murine interleukin-1β. Clin. Vaccine Immunol. 2010, 17, 43–48. [Google Scholar] [CrossRef]
- Kajikawa, A.; Nordone, S.K.; Zhang, L.; Stoeker, L.L.; LaVoy, A.S.; Klaenhammer, T.R.; Dean, G.A. Dissimilar properties of two recombinant Lactobacillus acidophilus strains displaying Salmonella FliC with different anchoring motifs. Appl. Environ. Microbiol. 2011, 77, 6587–6596. [Google Scholar] [CrossRef]
- Kajikawa, A.; Zhang, L.; LaVoy, A.; Bumgardner, S.; Klaenhammer, T.R.; Dean, G.A. Mucosal Immunogenicity of Genetically Modified Lactobacillus acidophilus Expressing an HIV-1 Epitope within the Surface Layer Protein. PLoS ONE 2015, 10, e0141713. [Google Scholar] [CrossRef] [PubMed]
- Bien, J.; Sokolova, O.; Bozko, P. Role of Uropathogenic Escherichia coli Virulence Factors in Development of Urinary Tract Infection and Kidney Damage. Int. J. Nephrol. 2012, 2012, 681473. [Google Scholar] [CrossRef] [PubMed]
- Azizi, A.; Kumar, A.; Diaz-Mitoma, F.; Mestecky, J. Enhancing oral vaccine potency by targeting intestinal M cells. PLoS Pathog. 2010, 6, e1001147. [Google Scholar] [CrossRef]
- Fransen, F.; Zagato, E.; Mazzini, E.; Fosso, B.; Manzari, C.; El Aidy, S.; Chiavelli, A.; D’Erchia, A.M.; Sethi, M.K.; Pabst, O.; et al. BALB/c and C57BL/6 Mice Differ. in Polyreactive IgA Abundance, which Impacts the Generation of Antigen-Specific IgA and Microbiota Diversity. Immunity 2015, 43, 527–540. [Google Scholar] [CrossRef]
- Martinoli, C.; Chiavelli, A.; Rescigno, M. Entry route of Salmonella typhimurium directs the type of induced immune response. Immunity 2007, 27, 975–984. [Google Scholar] [CrossRef]
- Mossman, K.L.; Mian, M.F.; Lauzon, N.M.; Gyles, C.L.; Lichty, B.; Mackenzie, R.; Gill, N.; Ashkar, A.A. Cutting edge: FimH adhesin of type 1 fimbriae is a novel TLR4 ligand. J. Immunol. 2008, 181, 6702–6706. [Google Scholar] [CrossRef] [PubMed]
- Baorto, D.M.; Gao, Z.; Malaviya, R.; Dustin, M.L.; van der Merwe, A.; Lublin, D.M.; Abraham, S.N. Survival of FimH-expressing enterobacteria in macrophages relies on glycolipid traffic. Nature 1997, 389, 636–639. [Google Scholar] [CrossRef]
- Fan, X.; Yue, Y.; Xiong, S. Incorporation of a bi-functional protein FimH enhances the immunoprotection of chitosan-pVP1 vaccine against coxsackievirus B3-induced myocarditis. Antiviral Res. 2017, 140, 121–132. [Google Scholar] [CrossRef] [PubMed]
- Zhang, W.; Xu, L.; Park, H.B.; Hwang, J.; Kwak, M.; Lee, P.C.W.; Liang, G.; Zhang, X.; Xu, J.; Jin, J.O. Escherichia coli adhesion portion FimH functions as an adjuvant for cancer immunotherapy. Nat. Commun. 2020, 11, 1187. [Google Scholar] [CrossRef]
- Douglas, G.L.; Goh, Y.J.; Klaenhammer, T.R. Integrative food grade expression system for lactic acid bacteria. Methods Mol. Biol. 2011, 765, 373–387. [Google Scholar] [CrossRef] [PubMed]
- Walker, D.C.; Aoyama, K.; Klaenhammer, T.R. Electrotransformation of lactobacillus acidophilus group A1. FEMS Microbiol. Lett. 1996, 138, 233–237. [Google Scholar] [CrossRef] [PubMed]
- Stoeker, L.; Nordone, S.; Gunderson, S.; Zhang, L.; Kajikawa, A.; LaVoy, A.; Miller, M.; Klaenhammer, T.R.; Dean, G.A. Assessment of Lactobacillus gasseri as a candidate oral vaccine vector. Clin. Vaccine Immunol. 2011, 18, 1834–1844. [Google Scholar] [CrossRef]
- Fukuda, S.; Hase, K.; Ohno, H. Application of a mouse ligated Peyer’s patch intestinal loop assay to evaluate bacterial uptake by M cells. J. Vis. Exp. 2011, 58, e3225. [Google Scholar] [CrossRef]
- Nolan, T.; Hands, R.E.; Ogunkolade, W.; Bustin, S.A. SPUD: A quantitative PCR assay for the detection of inhibitors in nucleic acid preparations. Anal. Biochem. 2006, 351, 308–310. [Google Scholar] [CrossRef]
- Choudhury, D.; Thompson, A.; Stojanoff, V.; Langermann, S.; Pinkner, J.; Hultgren, S.J.; Knight, S.D. X-ray structure of the FimC-FimH chaperone-adhesin complex from uropathogenic Escherichia coli. Science 1999, 285, 1061–1066. [Google Scholar] [CrossRef]
- Schembri, M.A.; Hasman, H.; Klemm, P. Expression and purification of the mannose recognition domain of the FimH adhesin. FEMS Microbiol. Lett. 2000, 188, 147–151. [Google Scholar] [CrossRef]
- Kandasamy, M.; Selvakumari Jayasurya, A.; Moochhala, S.; Huat Bay, B.; Kun Lee, Y.; Mahendran, R. Lactobacillus rhamnosus GG secreting an antigen and Interleukin-2 translocates across the gastrointestinal tract and induces an antigen specific immune response. Microbiol. Immunol. 2011, 55, 704–714. [Google Scholar] [CrossRef]
- Cerovic, V.; Bain, C.C.; Mowat, A.M.; Milling, S.W. Intestinal macrophages and dendritic cells: What’s the difference? Trends Immunol. 2014, 35, 270–277. [Google Scholar] [CrossRef] [PubMed]
- Tamoutounour, S.; Henri, S.; Lelouard, H.; de Bovis, B.; de Haar, C.; van der Woude, C.J.; Woltman, A.M.; Reyal, Y.; Bonnet, D.; Sichien, D.; et al. CD64 distinguishes macrophages from dendritic cells in the gut and reveals the Th1-inducing role of mesenteric lymph node macrophages during colitis. Eur. J. Immunol. 2012, 42, 3150–3166. [Google Scholar] [CrossRef] [PubMed]
- Holmgren, J.; Czerkinsky, C. Mucosal immunity and vaccines. Nat. Med. 2005, 11, S45–S53. [Google Scholar] [CrossRef] [PubMed]
- Wang, Z.; Yu, Q.; Gao, J.; Yang, Q. Mucosal and systemic immune responses induced by recombinant Lactobacillus spp. expressing the hemagglutinin of the avian influenza virus H5N1. Clin. Vaccine Immunol. 2012, 19, 174–179. [Google Scholar] [CrossRef]
- Mohamadzadeh, M.; Durmaz, E.; Zadeh, M.; Pakanati, K.C.; Gramarossa, M.; Cohran, V.; Klaenhammer, T.R. Targeted expression of anthrax protective antigen by Lactobacillus gasseri as an anthrax vaccine. Future Microbiol. 2010, 5, 1289–1296. [Google Scholar] [CrossRef]
- Kajikawa, A.; Zhang, L.; Long, J.; Nordone, S.; Stoeker, L.; LaVoy, A.; Bumgardner, S.; Klaenhammer, T.; Dean, G. Construction and immunological evaluation of dual cell surface display of HIV-1 gag and Salmonella enterica serovar Typhimurium FliC in Lactobacillus acidophilus for vaccine delivery. Clin. Vaccine Immunol. 2012, 19, 1374–1381. [Google Scholar] [CrossRef]
- Moeini, H.; Rahim, R.A.; Omar, A.R.; Shafee, N.; Yusoff, K. Lactobacillus acidophilus as a live vehicle for oral immunization against chicken anemia virus. Appl. Microbiol. Biotechnol. 2011, 90, 77–88. [Google Scholar] [CrossRef]
- Hongying, F.; Xianbo, W.; Fang, Y.; Yang, B.; Beiguo, L. Oral immunization with recombinant Lactobacillus acidophilus expressing the adhesin Hp0410 of Helicobacter pylori induces mucosal and systemic immune responses. Clin. Vaccine Immunol. 2014, 21, 126–132. [Google Scholar] [CrossRef]
- O’Flaherty, S.; Klaenhammer, T.R. Multivalent Chromosomal Expression of the Clostridium botulinum Serotype A Neurotoxin Heavy-Chain Antigen and the Bacillus anthracis Protective Antigen in Lactobacillus acidophilus. Appl. Environ. Microbiol. 2016, 82, 6091–6101. [Google Scholar] [CrossRef]
- Lin, R.; Zhang, Y.; Long, B.; Li, Y.; Wu, Y.; Duan, S.; Zhu, B.; Wu, X.; Fan, H. Oral Immunization with Recombinant Lactobacillus acidophilus Expressing espA-Tir-M Confers Protection against Enterohemorrhagic Escherichia coli O157:H7 Challenge in Mice. Front. Microbiol. 2017, 8, 417. [Google Scholar] [CrossRef]
- Chu, H.; Kang, S.; Ha, S.; Cho, K.; Park, S.M.; Han, K.H.; Kang, S.K.; Lee, H.; Han, S.H.; Yun, C.H.; et al. Lactobacillus acidophilus expressing recombinant K99 adhesive fimbriae has an inhibitory effect on adhesion of enterotoxigenic Escherichia coli. Microbiol. Immunol. 2005, 49, 941–948. [Google Scholar] [CrossRef] [PubMed]
- Zhu, L.F.; Long, B.G.; Luo, J.; Jiang, R.; Fang, H.Y. Construction of a recombinant Lactobacillus acidophilus expressing high levels of Helicobacter pylori adhesin Hp0410. Nan Fang Yi Ke Da Xue Xue Bao 2010, 30, 334–337. [Google Scholar] [PubMed]
- Vilander, A.C.; Dean, G.A. Adjuvant Strategies for Lactic Acid Bacterial Mucosal Vaccines. Vaccines 2019, 7, 150. [Google Scholar] [CrossRef] [PubMed]
- Yanagihara, S.; Kanaya, T.; Fukuda, S.; Nakato, G.; Hanazato, M.; Wu, X.R.; Yamamoto, N.; Ohno, H. Uromodulin-SlpA binding dictates Lactobacillus acidophilus uptake by intestinal epithelial M cells. Int. Immunol. 2017, 29, 357–363. [Google Scholar] [CrossRef]
- Chang, S.Y.; Ko, H.J.; Kweon, M.N. Mucosal dendritic cells shape mucosal immunity. Exp. Mol. Med. 2014, 46, e84. [Google Scholar] [CrossRef]
- Cerutti, A. The regulation of IgA class switching. Nat. Rev. Immunol. 2008, 8, 421–434. [Google Scholar] [CrossRef]
- Ehrhardt, R.O.; Strober, W.; Harriman, G.R. Effect of transforming growth factor (TGF)-beta 1 on IgA isotype expression. TGF-beta 1 induces a small increase in sIgA+ B cells regardless of the method of B cell activation. J. Immunol. 1992, 148, 3830–3836. [Google Scholar] [CrossRef]
- Konkel, J.E.; Chen, W. Balancing acts: The role of TGF-beta in the mucosal immune system. Trends Mol. Med. 2011, 17, 668–676. [Google Scholar] [CrossRef] [PubMed]
- Chen, C.C.; Louie, S.; Shi, H.N.; Walker, W.A. Preinoculation with the probiotic Lactobacillus acidophilus early in life effectively inhibits murine Citrobacter rodentium colitis. Pediatr Res. 2005, 58, 1185–1191. [Google Scholar] [CrossRef]
- Seo, G.Y.; Youn, J.; Kim, P.H. IL-21 ensures TGF-beta 1-induced IgA isotype expression in mouse Peyer’s patches. J. Leukoc. Biol. 2009, 85, 744–750. [Google Scholar] [CrossRef]
- Cao, A.T.; Yao, S.; Gong, B.; Nurieva, R.I.; Elson, C.O.; Cong, Y. Interleukin (IL)-21 promotes intestinal IgA response to microbiota. Mucosal Immunol. 2015, 8, 1072–1082. [Google Scholar] [CrossRef] [PubMed]
- Li, S.N.; Wang, W.; Fu, S.P.; Wang, J.F.; Liu, H.M.; Xie, S.S.; Liu, B.R.; Li, Y.; Lv, Q.K.; Li, Z.Q.; et al. IL-21 modulates release of proinflammatory cytokines in LPS-stimulated macrophages through distinct signaling pathways. Mediators Inflamm. 2013, 2013, 548073. [Google Scholar] [CrossRef] [PubMed]
- Iwata, M.; Hirakiyama, A.; Eshima, Y.; Kagechika, H.; Kato, C.; Song, S.Y. Retinoic acid imprints gut-homing specificity on T cells. Immunity 2004, 21, 527–538. [Google Scholar] [CrossRef] [PubMed]
- Fischer, H.; Yamamoto, M.; Akira, S.; Beutler, B.; Svanborg, C. Mechanism of pathogen-specific TLR4 activation in the mucosa: Fimbriae, recognition receptors and adaptor protein selection. Eur. J. Immunol. 2006, 36, 267–277. [Google Scholar] [CrossRef] [PubMed]
- Habibi, M.; Asadi Karam, M.R.; Bouzari, S. In silico Study of Toll-Like Receptor 4 Binding Site of FimH from Uropathogenic Escherichia coli. J. Med. Microbiol. Infect. Dis. 2014, 2, 35–39. [Google Scholar]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Plasmids | ||||
| Name | Surface-Expressed Protein | Tag | Surface Anchor | |
| pD001 | FimH | No tag | Amub | |
| pD002 | FimH | 5′-FLAG-tag | Amub | |
| pD003 | FimH | 3′-FLAG-tag | Amub | |
| LA Strains | ||||
| Name | slpA Epitope Insert | Plasmid | Antibiotic Resistance | Reference |
| NCK56 | None | None | None | NCFM Strain |
| GAD31 | HIV-1 MPER | None | Erythromycin | [5] |
| GAD40 | HIV-1 MPER | pD001 | Erythromycin | Here |
| GAD41 | HIV-1 MPER | pD002 | Erythromycin | Here |
| GAD42 | HIV-1 MPER | pD003 | Erythromycin | Here |
| GAD80 | VP8-10pep | None | Erythromycin | Here |
| GAD83 | VP8-10pep | pD001 | Erythromycin | Here |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Vilander, A.C.; Shelton, K.; LaVoy, A.; Dean, G.A. Expression of E. coli FimH Enhances Trafficking of an Orally Delivered Lactobacillus acidophilus Vaccine to Immune Inductive Sites via Antigen-Presenting Cells. Vaccines 2023, 11, 1162. https://doi.org/10.3390/vaccines11071162
Vilander AC, Shelton K, LaVoy A, Dean GA. Expression of E. coli FimH Enhances Trafficking of an Orally Delivered Lactobacillus acidophilus Vaccine to Immune Inductive Sites via Antigen-Presenting Cells. Vaccines. 2023; 11(7):1162. https://doi.org/10.3390/vaccines11071162
Chicago/Turabian StyleVilander, Allison C., Kimberly Shelton, Alora LaVoy, and Gregg A. Dean. 2023. "Expression of E. coli FimH Enhances Trafficking of an Orally Delivered Lactobacillus acidophilus Vaccine to Immune Inductive Sites via Antigen-Presenting Cells" Vaccines 11, no. 7: 1162. https://doi.org/10.3390/vaccines11071162
APA StyleVilander, A. C., Shelton, K., LaVoy, A., & Dean, G. A. (2023). Expression of E. coli FimH Enhances Trafficking of an Orally Delivered Lactobacillus acidophilus Vaccine to Immune Inductive Sites via Antigen-Presenting Cells. Vaccines, 11(7), 1162. https://doi.org/10.3390/vaccines11071162