
Neutralizing Antibody Responses Elicited by Inactivated Whole Virus and Genetic Vaccines against Dominant SARS-CoV-2 Variants during the Four Epidemic Peaks of COVID-19 in Colombia

 , , , and
, , , and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Patients and Samples
2.2. Virus Isolation
2.3. Microneutralization and Binding Antibody Assays
2.4. Statistical Analysis
3. Results
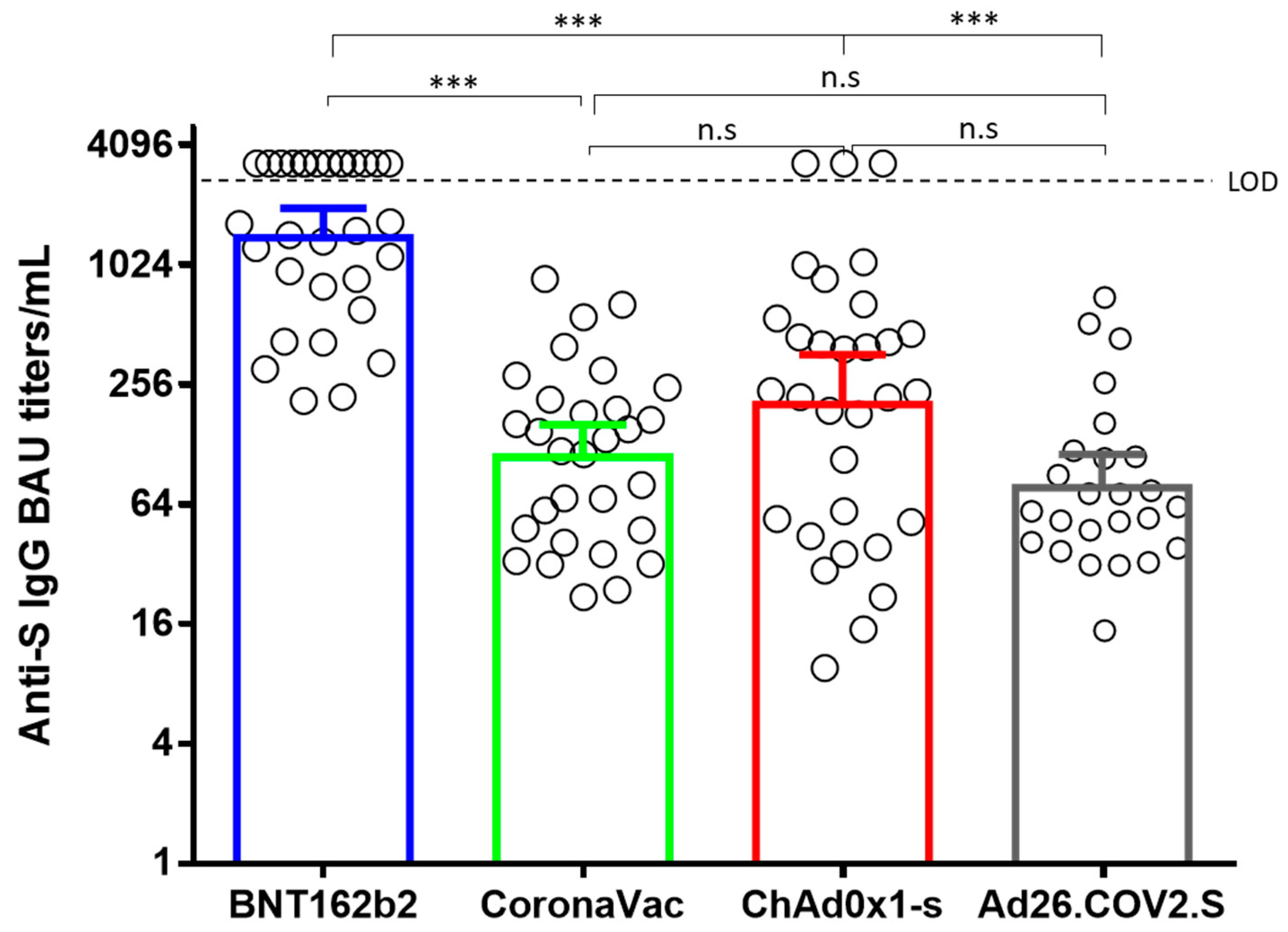
3.1. Vaccine Elicited Anti-S IgG Titers Correlate with Neutralizing Antibody Titers
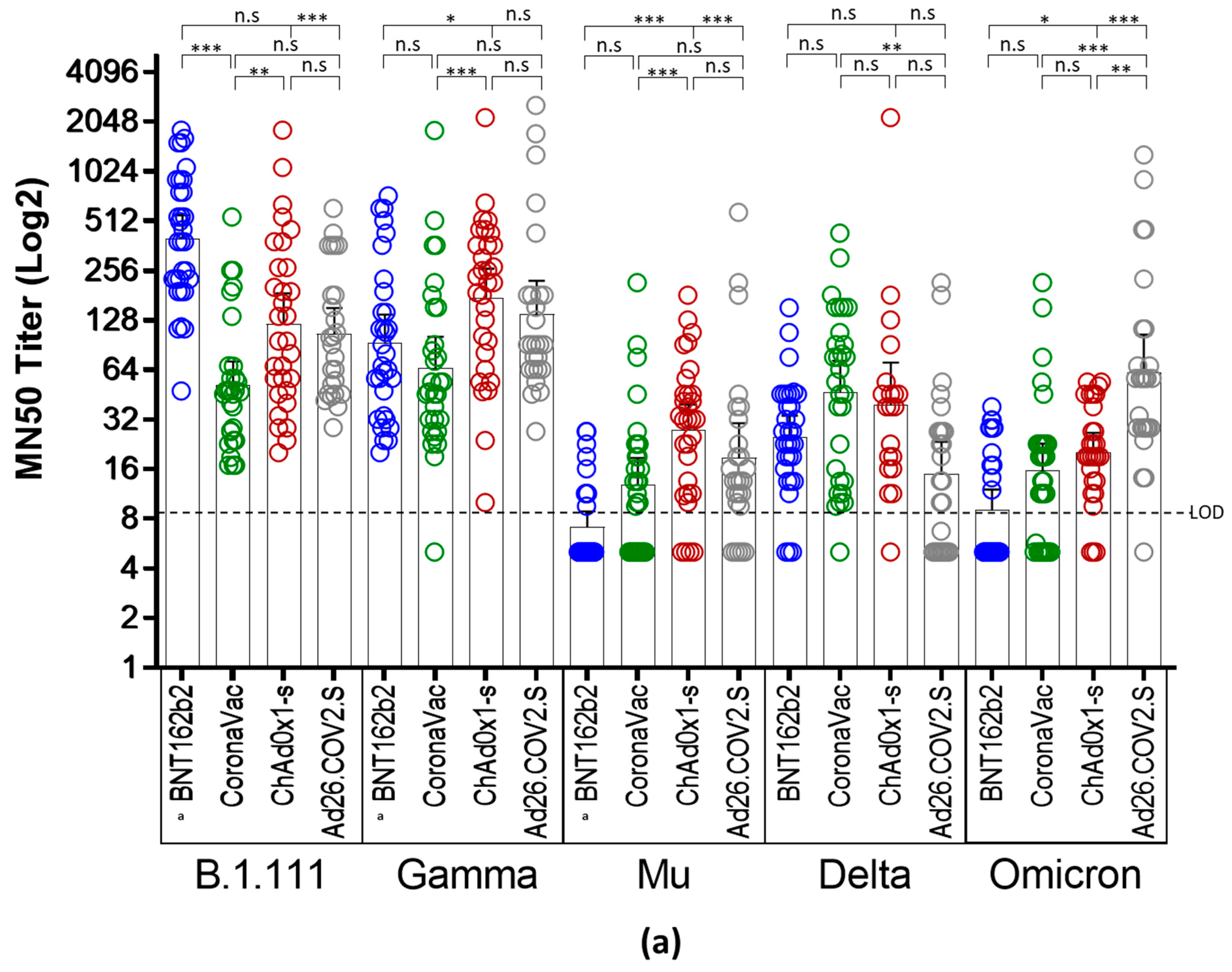
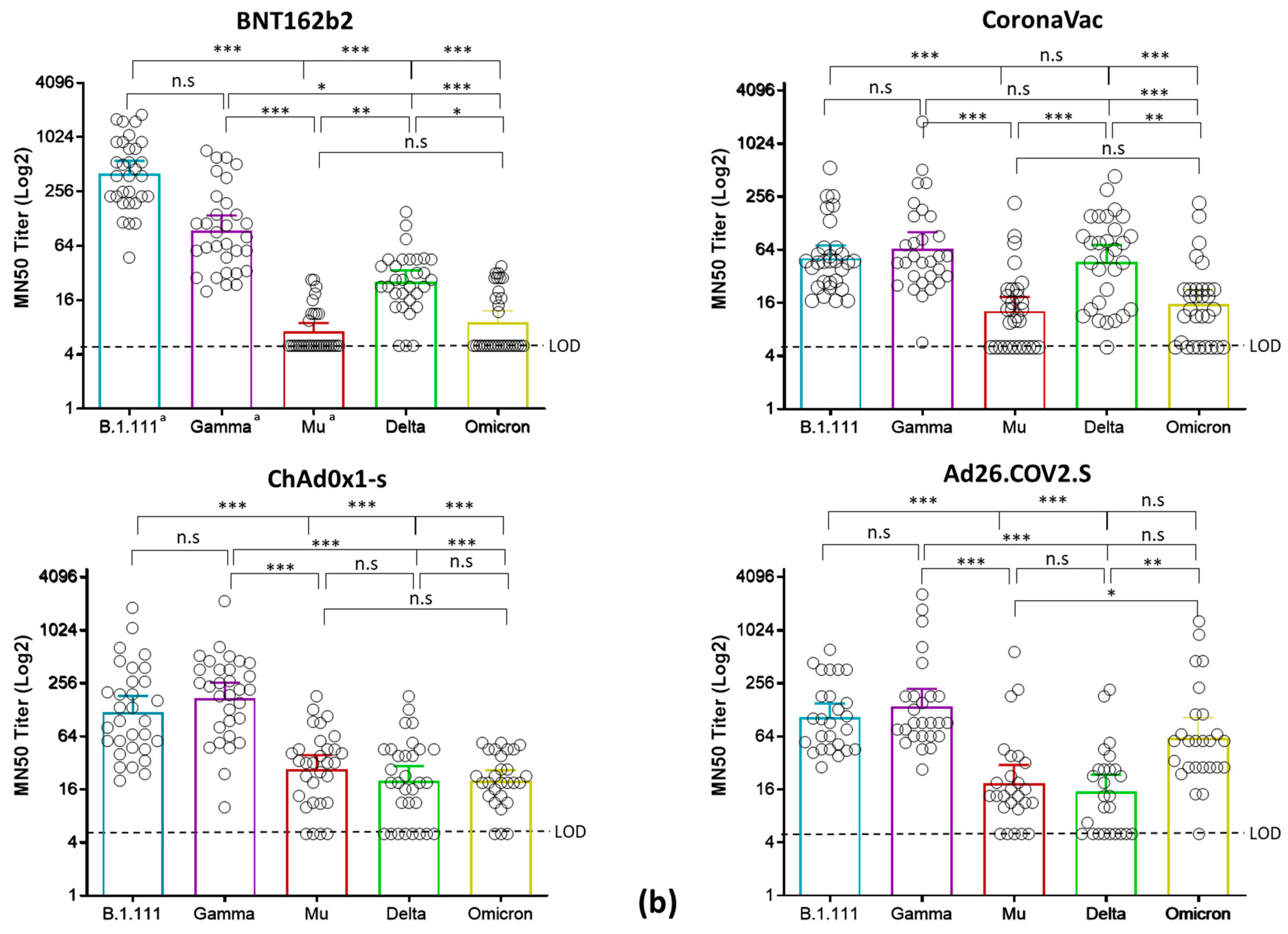
3.2. Reduced nAb Responses against Mu, Delta, and Omicron Variants in Vaccinated Individuals with BNT162b2, CoronaVac, ChAdOx1, and Ad26.COV2.S
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Minsalud. Dosis Aplicadas Contra COVID-19. Available online: https://app.powerbi.com/view?r=eyJrIjoiNThmZTJmZWYtOWFhMy00OGE1LWFiNDAtMTJmYjM0NDA5NGY2IiwidCI6ImJmYjdlMTNhLTdmYjctNDAxNi04MzBjLWQzNzE2ZThkZDhiOCJ9 (accessed on 14 July 2022).
- Minsalud. Vacunación Contra COVID-19. Available online: https://www.minsalud.gov.co/salud/publica/Vacunacion/Paginas/Vacunacion-covid-19.aspx (accessed on 11 March 2022).
- Fiolet, T.; Kherabi, Y.; MacDonald, C.J.; Ghosn, J.; Peiffer-Smadja, N. Comparing COVID-19 vaccines for their characteristics, efficacy and effectiveness against SARS-CoV-2 and variants of concern: A narrative review. Clin. Microbiol. Infect. 2022, 28, 202–221. [Google Scholar] [CrossRef] [PubMed]
- Alvarez-Diaz, D.A.; Munoz, A.L.; Tavera-Rodriguez, P.; Herrera-Sepulveda, M.T.; Ruiz-Moreno, H.A.; Laiton-Donato, K.; Franco-Munoz, C.; Pelaez-Carvajal, D.; Cuellar, D.; Munoz-Suarez, A.M.; et al. Low Neutralizing Antibody Titers against the Mu Variant of SARS-CoV-2 in 31 BNT162b2 Vaccinated Individuals in Colombia. Vaccines 2022, 10, 180. [Google Scholar] [CrossRef] [PubMed]
- Alvarez-Diaz, D.A.; Laiton-Donato, K.; Torres-Garcia, O.A.; Ruiz-Moreno, H.A.; Franco-Munoz, C.; Beltran, M.A.; Mercado-Reyes, M.; Rueda, M.G.; Munoz, A.L. Reduced levels of convalescent neutralizing antibodies against SARS-CoV-2 B.1+L249S+E484K lineage. Virus Res. 2022, 308, 198629. [Google Scholar] [CrossRef]
- INS. Coronavirus (COVID-2019) en Colombia: Genoma. Available online: https://www.ins.gov.co/Noticias/Paginas/coronavirus-genoma.aspx (accessed on 18 April 2022).
- Faria, N.R.; Mellan, T.A.; Whittaker, C.; Claro, I.M.; Candido, D.D.S.; Mishra, S.; Crispim, M.A.E.; Sales, F.C.S.; Hawryluk, I.; McCrone, J.T.; et al. Genomics and epidemiology of the P.1 SARS-CoV-2 lineage in Manaus, Brazil. Science 2021, 372, 815–821. [Google Scholar] [CrossRef]
- Dejnirattisai, W.; Zhou, D.; Supasa, P.; Liu, C.; Mentzer, A.J.; Ginn, H.M.; Zhao, Y.; Duyvesteyn, H.M.E.; Tuekprakhon, A.; Nutalai, R.; et al. Antibody evasion by the P.1 strain of SARS-CoV-2. Cell 2021, 184, 2939–2954.e9. [Google Scholar] [CrossRef] [PubMed]
- Souza, W.; Amorim, M.; Sesti-Costa, R.; Coimbra, L.D.; Brunetti, N.; Toledo-Teixeira, D.A.; Souza, G.; Muraro, S.; Lorencini Parise, P.; Barbosa, P.; et al. Neutralisation of SARS-CoV-2 lineage P.1 by antibodies elicited through natural SARS-CoV-2 infection or vaccination with an inactivated SARS-CoV-2 vaccine: An immunological study. Lancet Microbe 2021, 2, e527–e535. [Google Scholar] [CrossRef] [PubMed]
- Laiton-Donato, K.; Franco-Munoz, C.; Alvarez-Diaz, D.A.; Ruiz-Moreno, H.A.; Usme-Ciro, J.A.; Prada, D.A.; Reales-Gonzalez, J.; Corchuelo, S.; Herrera-Sepulveda, M.T.; Naizaque, J.; et al. Characterization of the emerging B.1.621 variant of interest of SARS-CoV-2. Infect. Genet. Evol. 2021, 95, 105038. [Google Scholar] [CrossRef]
- Uriu, K.; Kimura, I.; Shirakawa, K.; Takaori-Kondo, A.; Nakada, T.A.; Kaneda, A.; Nakagawa, S.; Sato, K. Neutralization of the SARS-CoV-2 Mu Variant by Convalescent and Vaccine Serum. N. Engl. J. Med. 2021, 385, 2397–2399. [Google Scholar] [CrossRef]
- Planas, D.; Veyer, D.; Baidaliuk, A.; Staropoli, I.; Guivel-Benhassine, F.; Rajah, M.M.; Planchais, C.; Porrot, F.; Robillard, N.; Puech, J.; et al. Reduced sensitivity of SARS-CoV-2 variant Delta to antibody neutralization. Nature 2021, 596, 276–280. [Google Scholar] [CrossRef]
- Edara, V.V.; Pinsky, B.A.; Suthar, M.S.; Lai, L.; Davis-Gardner, M.E.; Floyd, K.; Flowers, M.W.; Wrammert, J.; Hussaini, L.; Ciric, C.R.; et al. Infection and Vaccine-Induced Neutralizing-Antibody Responses to the SARS-CoV-2 B.1.617 Variants. N. Engl. J. Med. 2021, 385, 664–666. [Google Scholar] [CrossRef]
- Wang, B.; Goh, Y.S.; Fong, S.W.; Young, B.E.; Ngoh, E.Z.X.; Chavatte, J.M.; Salleh, S.N.M.; Yeo, N.K.; Amrun, S.N.; Hor, P.X.; et al. Resistance of SARS-CoV-2 Delta variant to neutralization by BNT162b2-elicited antibodies in Asians. Lancet Reg. Health West. Pac. 2021, 15, 100276. [Google Scholar] [CrossRef] [PubMed]
- Gangavarapu, K.; Latif, A.; Mullen, J.; Alkuzweny, M.; Hufbauer, E.; Tsueng, G.; Haag, E.; Zeller, M.; Aceves, C.; Zaiets, K.; et al. Lineage Comparison. Available online: https://outbreak.info/compare-lineages?pango=Gamma&pango=BA.1&pango=Mu&pango=Delta&pango=BA.2%2a%20%5BOmicron%20%28BA.2.X%29%5D&pango=BA.4&gene=S&threshold=75&nthresh=1&sub=false&dark=false (accessed on 22 March 2022).
- Messali, S.; Bertelli, A.; Campisi, G.; Zani, A.; Ciccozzi, M.; Caruso, A.; Caccuri, F. A cluster of the new SARS-CoV-2 B.1.621 lineage in Italy and sensitivity of the viral isolate to the BNT162b2 vaccine. J. Med. Virol. 2021, 93, 6468–6470. [Google Scholar] [CrossRef] [PubMed]
- WHO. Interim Statement on Booster Doses for COVID-19 Vaccination. Available online: https://www.who.int/news/item/22-12-2021-interim-statement-on-booster-doses-for-covid-19-vaccination---update-22-december-2021 (accessed on 11 March 2022).
- Lu, L.; Mok, B.W.; Chen, L.L.; Chan, J.M.; Tsang, O.T.; Lam, B.H.; Chuang, V.W.; Chu, A.W.; Chan, W.M.; Ip, J.D.; et al. Neutralization of SARS-CoV-2 Omicron variant by sera from BNT162b2 or Coronavac vaccine recipients. Clin. Infect. Dis. 2021, 75, e822–e826. [Google Scholar] [CrossRef] [PubMed]
- Malagon-Rojas, J.; Mercado-Reyes, M.; Toloza-Perez, Y.G.; Galindo, M.; Palma, R.M.; Catama, J.; Bedoya, J.F.; Parra-Barrera, E.L.; Meneses, X.; Barbosa, J.; et al. Comparison of Anti-SARS-CoV-2 IgG Antibody Responses Generated by the Administration of Ad26.COV2.S, AZD1222, BNT162b2, or CoronaVac: Longitudinal Prospective Cohort Study in the Colombian Population, 2021/2022. Vaccines 2022, 10, 1609. [Google Scholar] [CrossRef] [PubMed]
- Reed, L.J.; Muench, H. A simple method of estimating fifty per cent endpoints. Am. J. Epidemiol. 1938, 27, 493–497. [Google Scholar] [CrossRef]
- Saker, K.; Escuret, V.; Pitiot, V.; Massardier-Pilonchery, A.; Paul, S.; Mokdad, B.; Langlois-Jacques, C.; Rabilloud, M.; Goncalves, D.; Fabien, N.; et al. Evaluation of Commercial Anti-SARS-CoV-2 Antibody Assays and Comparison of Standardized Titers in Vaccinated Health Care Workers. J. Clin. Microbiol. 2022, 60, e0174621. [Google Scholar] [CrossRef]
- Fukutani, K.F.; Barreto, M.L.; Andrade, B.B.; Queiroz, A.T.L. Correlation between SARS-Cov-2 Vaccination, COVID-19 Incidence and Mortality: Tracking the Effect of Vaccination on Population Protection in Real Time. Front. Genet. 2021, 12, 679485. [Google Scholar] [CrossRef] [PubMed]
- Heinz, F.X.; Stiasny, K. Distinguishing features of current COVID-19 vaccines: Knowns and unknowns of antigen presentation and modes of action. NPJ Vaccines 2021, 6, 104. [Google Scholar] [CrossRef] [PubMed]
- Bian, L.; Liu, J.; Gao, F.; Gao, Q.; He, Q.; Mao, Q.; Wu, X.; Xu, M.; Liang, Z. Research progress on vaccine efficacy against SARS-CoV-2 variants of concern. Hum. Vaccines Immunother. 2022, 18, 2057161. [Google Scholar] [CrossRef]
- Lopera, T.J.; Chvatal-Medina, M.; Florez-Alvarez, L.; Zapata-Cardona, M.I.; Taborda, N.A.; Rugeles, M.T.; Hernandez, J.C. Humoral Response to BNT162b2 Vaccine against SARS-CoV-2 Variants Decays after Six Months. Front. Immunol. 2022, 13, 879036. [Google Scholar] [CrossRef]
- Yu, J.; Collier, A.Y.; Rowe, M.; Mardas, F.; Ventura, J.D.; Wan, H.; Miller, J.; Powers, O.; Chung, B.; Siamatu, M.; et al. Neutralization of the SARS-CoV-2 Omicron BA.1 and BA.2 Variants. N. Engl. J. Med. 2022, 386, 1579–1580. [Google Scholar] [CrossRef] [PubMed]
- Ou, J.; Lan, W.; Wu, X.; Zhao, T.; Duan, B.; Yang, P.; Ren, Y.; Quan, L.; Zhao, W.; Seto, D.; et al. Tracking SARS-CoV-2 Omicron diverse spike gene mutations identifies multiple inter-variant recombination events. Signal Transduct. Target. Ther. 2022, 7, 138. [Google Scholar] [CrossRef] [PubMed]
- Magazine, N.; Zhang, T.; Wu, Y.; McGee, M.C.; Veggiani, G.; Huang, W. Mutations and Evolution of the SARS-CoV-2 Spike Protein. Viruses 2022, 14, 640. [Google Scholar] [CrossRef] [PubMed]
- Fratev, F. R346K Mutation in the Mu Variant of SARS-CoV-2 Alters the Interactions with Monoclonal Antibodies from Class 2: A Free Energy Perturbation Study. J. Chem. Inf. Model. 2022, 62, 627–631. [Google Scholar] [CrossRef] [PubMed]
- Koyama, T.; Miyakawa, K.; Tokumasu, R.; Jeremiah, S.S.; Kudo, M.; Ryo, A. Evasion of vaccine-induced humoral immunity by emerging sub-variants of SARS-CoV-2. Future Microbiol. 2022, 17, 417–424. [Google Scholar] [CrossRef] [PubMed]
- McLean, G.; Kamil, J.; Lee, B.; Moore, P.; Schulz, T.F.; Muik, A.; Sahin, U.; Tureci, O.; Pather, S. The Impact of Evolving SARS-CoV-2 Mutations and Variants on COVID-19 Vaccines. mBio 2022, 13, e0297921. [Google Scholar] [CrossRef]
- Muttineni, R.; Binitha, R.N.; Putty, K.; Marapakala, K.; Sandra, K.P.; Panyam, J.; Vemula, A.; Singh, S.M.; Balachandran, S.; Viroji Rao, S.T.; et al. SARS-CoV-2 variants and spike mutations involved in second wave of COVID-19 pandemic in India. Transbound. Emerg. Dis. 2022, 69, e1721–e1733. [Google Scholar] [CrossRef]
- Di Giacomo, S.; Mercatelli, D.; Rakhimov, A.; Giorgi, F.M. Preliminary report on severe acute respiratory syndrome coronavirus 2 (SARS-CoV-2) Spike mutation T478K. J. Med. Virol. 2021, 93, 5638–5643. [Google Scholar] [CrossRef]
- Andrews, N.; Stowe, J.; Kirsebom, F.; Toffa, S.; Rickeard, T.; Gallagher, E.; Gower, C.; Kall, M.; Groves, N.; O’Connell, A.M.; et al. Covid-19 Vaccine Effectiveness against the Omicron (B.1.1.529) Variant. N. Engl. J. Med. 2022, 386, 1532–1546. [Google Scholar] [CrossRef]
- Speletas, M.; Voulgaridi, I.; Sarrou, S.; Dadouli, A.; Mouchtouri, V.A.; Nikoulis, D.J.; Tsakona, M.; Kyritsi, M.A.; Peristeri, A.M.; Avakian, I.; et al. Intensity and Dynamics of Anti-SARS-CoV-2 Immune Responses after BNT162b2 mRNA Vaccination: Implications for Public Health Vaccination Strategies. Vaccines 2022, 10, 316. [Google Scholar] [CrossRef]
- Lau, E.H.Y.; Tsang, O.T.Y.; Hui, D.S.C.; Kwan, M.Y.W.; Chan, W.H.; Chiu, S.S.; Ko, R.L.W.; Chan, K.H.; Cheng, S.M.S.; Perera, R.; et al. Neutralizing antibody titres in SARS-CoV-2 infections. Nat. Commun. 2021, 12, 63. [Google Scholar] [CrossRef] [PubMed]
- Gharbharan, A.; Jordans, C.C.E.; GeurtsvanKessel, C.; den Hollander, J.G.; Karim, F.; Mollema, F.P.N.; Stalenhoef-Schukken, J.E.; Dofferhoff, A.; Ludwig, I.; Koster, A.; et al. Effects of potent neutralizing antibodies from convalescent plasma in patients hospitalized for severe SARS-CoV-2 infection. Nat. Commun. 2021, 12, 3189. [Google Scholar] [CrossRef] [PubMed]
- Moss, P. The T cell immune response against SARS-CoV-2. Nat. Immunol. 2022, 23, 186–193. [Google Scholar] [CrossRef] [PubMed]
- Bertoletti, A.; Le Bert, N.; Tan, A.T. SARS-CoV-2-specific T cells in the changing landscape of the COVID-19 pandemic. Immunity 2022, 55, 1764–1778. [Google Scholar] [CrossRef] [PubMed]


{kind=link}
{kind=link}
{kind=link}
| Vaccine | GMBA Titers (BAU/mL)/(95% CI) | Comparison | Spearman r * | p-Value | |
|---|---|---|---|---|---|
| BNT16b2 | 1353 (959.9–1907) | a MN50 B.1.111 | a Anti-S IgG titer BAU/mL | 0.674 | <0.0001 |
| a MN50 Gamma (P.1) | a Anti-S IgG titer BAU/mL | 0.533 | 0.002 | ||
| a MN50 Mu (B.1.621) | a Anti-S IgG titer BAU/mL | 0.511 | <0.003 | ||
| MN50 Delta (AY.25.1) | Anti-S IgG titer BAU/mL | 0.258 | 0.161 | ||
| MN50 Omicron (BA.1) | Anti-S IgG titer BAU/mL | −0.089 | 0.6302 | ||
| CoronaVac | 111.3 (76.31–162.4) | MN50 B.1.111 | Anti-S IgG titer BAU/mL | 0.709 | <0.0001 |
| MN50 Gamma (P.1) | Anti-S IgG titer BAU/mL | 0.453 | 0.0118 | ||
| MN50 Mu (B.1.621) | Anti-S IgG titer BAU/mL | 0.627 | <0.0002 | ||
| MN50 Delta (AY.25.1) | Anti-S IgG titer BAU/mL | 0.580 | 0.0008 | ||
| MN50 Omicron (BA.1) | Anti-S IgG titer BAU/mL | 0.478 | 0.0075 | ||
| ChAd0x1-s | 198 (113.8–345) | MN50 B.1.111 | Anti-S IgG titer BAU/mL | 0.656 | <0.0001 |
| MN50 Gamma (P.1) | Anti-S IgG titer BAU/mL | 0.773 | <0.0001 | ||
| MN50 Mu (B.1.621) | Anti-S IgG titer BAU/mL | 0.723 | <0.0001 | ||
| MN50 Delta (AY.25.1) | Anti-S IgG titer BAU/mL | 0.8 | <0.0001 | ||
| MN50 Omicron (BA.1) | Anti-S IgG titer BAU/mL | 0.575 | 0.0013 | ||
| Ad26.COV2.S | 78.36 (53.2–115.4) | MN50 B.1.111 | Anti-S IgG titer BAU/mL | 0.625 | 0.0008 |
| MN50 Gamma (P.1) | Anti-S IgG titer BAU/mL | 0.725 | <0.0001 | ||
| MN50 Mu (B.1.621) | Anti-S IgG titer BAU/mL | 0.597 | 0.0016 | ||
| MN50 Delta (AY.25.1) | Anti-S IgG titer BAU/mL | 0.779 | <0.0001 | ||
| MN50 Omicron (BA.1) | Anti-S IgG titer BAU/mL | 0.389 | 0.054 | ||
| Vaccine | B.1.111 (Ancestral Virus) | P.1 (Gamma) | B.1.621 (Mu) | B.1.617.2 (Delta AY.25.1) | BA.1 (Omicron) | |
|---|---|---|---|---|---|---|
| Seropositive rate * % (no. positive/total) | BNT162b2 | a 100% (31/31) | a 100% (31/31) | a 32.2% (10/31) | 90.3% (28/31) | 41.2% (13/31) |
| CoronaVac | 100% (30/30) | 100% (30/30) | 63.3% (19/30) | 96.6% (29/30) | 73.3% (22/30) | |
| ChAd0x1-s | 100% (31/31) | 100% (31/31) | 87% (27/31) | 74.2% (23/31) | 89,3% (25/28) | |
| Ad26.COV2.S | 100% (26/26) | 100% (26/26) | 80.7% (21/26) | 65.4% (17/26) | 96.2% (25/26) | |
| Geometric mean TCID50 titer (95% CI) | BNT162b2 | a 401.3 (288.0–559.2) | a 94.02 (63.61–139) | a 7.2 (5.8–8.9) | 25.3 (19–34) | 25.4 (18.9–33.9) |
| CoronaVac | 51.7 (42.6–124.5) | 65.7 (42.2–102.1) | 12.9 (8.9–18.7) | 47.1 (30.4–72.9) | 15.6 (10.7–22.9) | |
| ChAd0x1-s | 121.6 (79.2–186.8) | 176.7 (118–264.6) | 27.6 (19.2–39.5) | 39.2 (21.6–71.4) | 20.3 (15.4–26.7) | |
| Ad26.COV2.S | 106.1 73.9–152.2) | 139.8 (87.4–223.6) | 18.8 (11.6–30.5) | 15.8 (9.6–23.6) | 62.1 (36.7–105.1) | |
| GMT Fold decrease relative to B.1.111 | BNT162b2 | - | 4.26 | 55.71 | 15.8 | 15.8 |
| CoronaVac | - | 0.78 | 4.0 | 1.1 | 3.3 | |
| ChAd0x1-s | - | 0.91 | 4.4 | 3.1 | 5.9 | |
| Ad26.COV2.S | - | 0.75 | 5.6 | 6.7 | 1.7 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Álvarez-Díaz, D.A.; Muñoz, A.L.; Herrera-Sepúlveda, M.T.; Tavera-Rodríguez, P.; Laiton-Donato, K.; Franco-Muñoz, C.; Ruiz-Moreno, H.A.; Galindo, M.; Catama, J.D.; Bermudez-Forero, A.; et al. Neutralizing Antibody Responses Elicited by Inactivated Whole Virus and Genetic Vaccines against Dominant SARS-CoV-2 Variants during the Four Epidemic Peaks of COVID-19 in Colombia. Vaccines 2022, 10, 2144. https://doi.org/10.3390/vaccines10122144
Álvarez-Díaz DA, Muñoz AL, Herrera-Sepúlveda MT, Tavera-Rodríguez P, Laiton-Donato K, Franco-Muñoz C, Ruiz-Moreno HA, Galindo M, Catama JD, Bermudez-Forero A, et al. Neutralizing Antibody Responses Elicited by Inactivated Whole Virus and Genetic Vaccines against Dominant SARS-CoV-2 Variants during the Four Epidemic Peaks of COVID-19 in Colombia. Vaccines. 2022; 10(12):2144. https://doi.org/10.3390/vaccines10122144
Chicago/Turabian StyleÁlvarez-Díaz, Diego A., Ana Luisa Muñoz, María T. Herrera-Sepúlveda, Pilar Tavera-Rodríguez, Katherine Laiton-Donato, Carlos Franco-Muñoz, Héctor Alejandro Ruiz-Moreno, Marisol Galindo, Jenssy D. Catama, Andrea Bermudez-Forero, and et al. 2022. "Neutralizing Antibody Responses Elicited by Inactivated Whole Virus and Genetic Vaccines against Dominant SARS-CoV-2 Variants during the Four Epidemic Peaks of COVID-19 in Colombia" Vaccines 10, no. 12: 2144. https://doi.org/10.3390/vaccines10122144
APA StyleÁlvarez-Díaz, D. A., Muñoz, A. L., Herrera-Sepúlveda, M. T., Tavera-Rodríguez, P., Laiton-Donato, K., Franco-Muñoz, C., Ruiz-Moreno, H. A., Galindo, M., Catama, J. D., Bermudez-Forero, A., & Mercado-Reyes, M. (2022). Neutralizing Antibody Responses Elicited by Inactivated Whole Virus and Genetic Vaccines against Dominant SARS-CoV-2 Variants during the Four Epidemic Peaks of COVID-19 in Colombia. Vaccines, 10(12), 2144. https://doi.org/10.3390/vaccines10122144