
Effect of Dietary Supplementation of Chestnut and Quebracho Tannin Supplementation on Neonatal Diarrhoea in Preweaning Calves

 ,
,  , , ,
, , , 
Abstract
1. Introduction
2. Materials and Methods
2.1. Animal Housing and Experimental Design
2.2. Zootechnical Performance, Diarrhoea Frequency and Sample Collection
2.3. Tannin Solubilization and Evaluation of Antioxidant Activity of Tannins Premixture
2.4. Nutrient Composition of the Whole Milk
2.5. Faecal Parameters and Apparent Total Tract Digestibility of Dietary Protein
2.6. Enzyme-Linked Immunosorbent Assay of Glucagon-like Peptide 2 and Diamine Oxidase in Serum Samples
2.7. Statistical Analysis
3. Results
3.1. Antioxidant Activity of Tannin Extract Premixture and Whole Milk Composition
3.2. Zootechnical Performance and Diarrhoea Occurrence
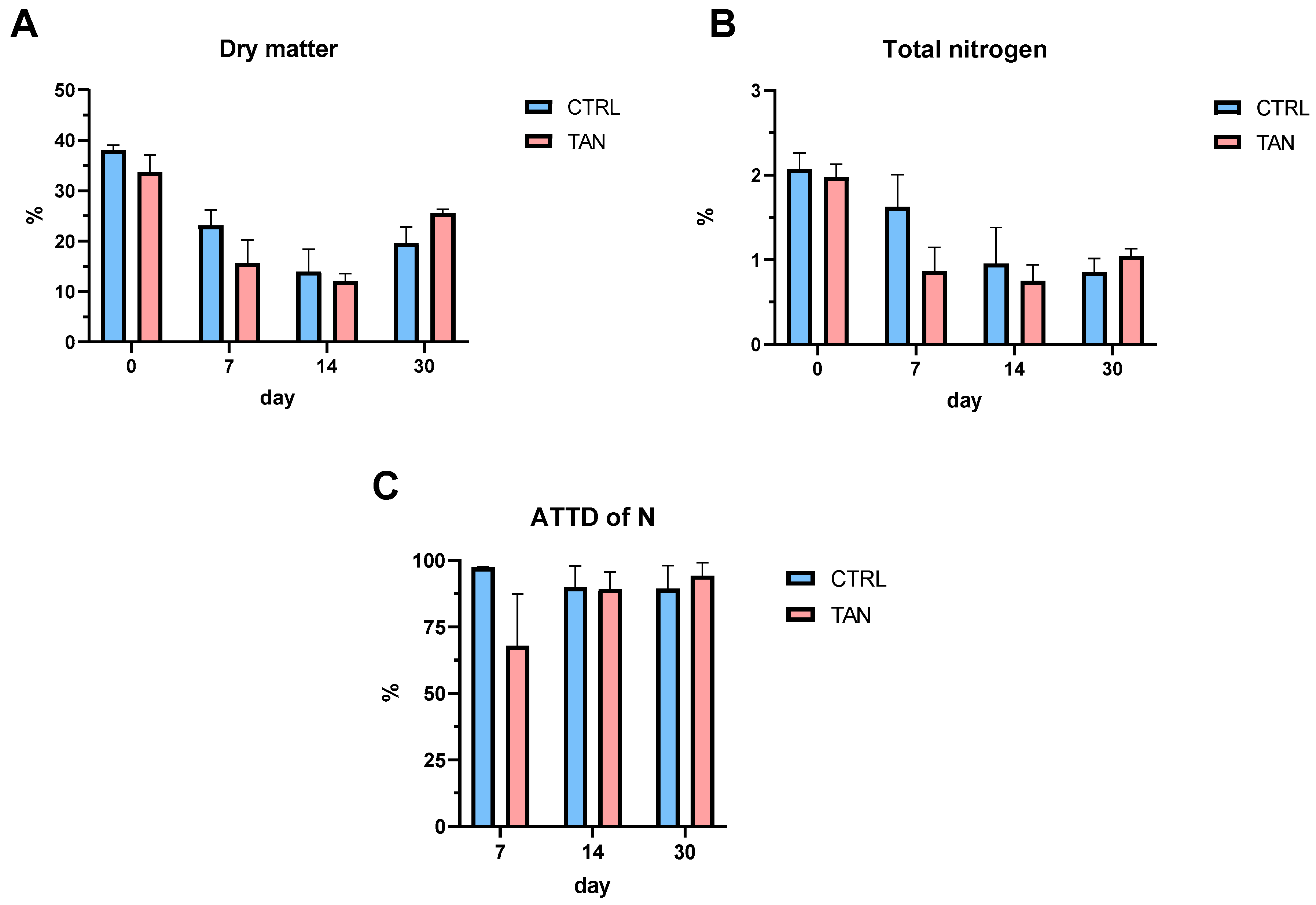
3.3. Faecal Parameters and Apparent Total Tract Digestibility of Dietary Protein
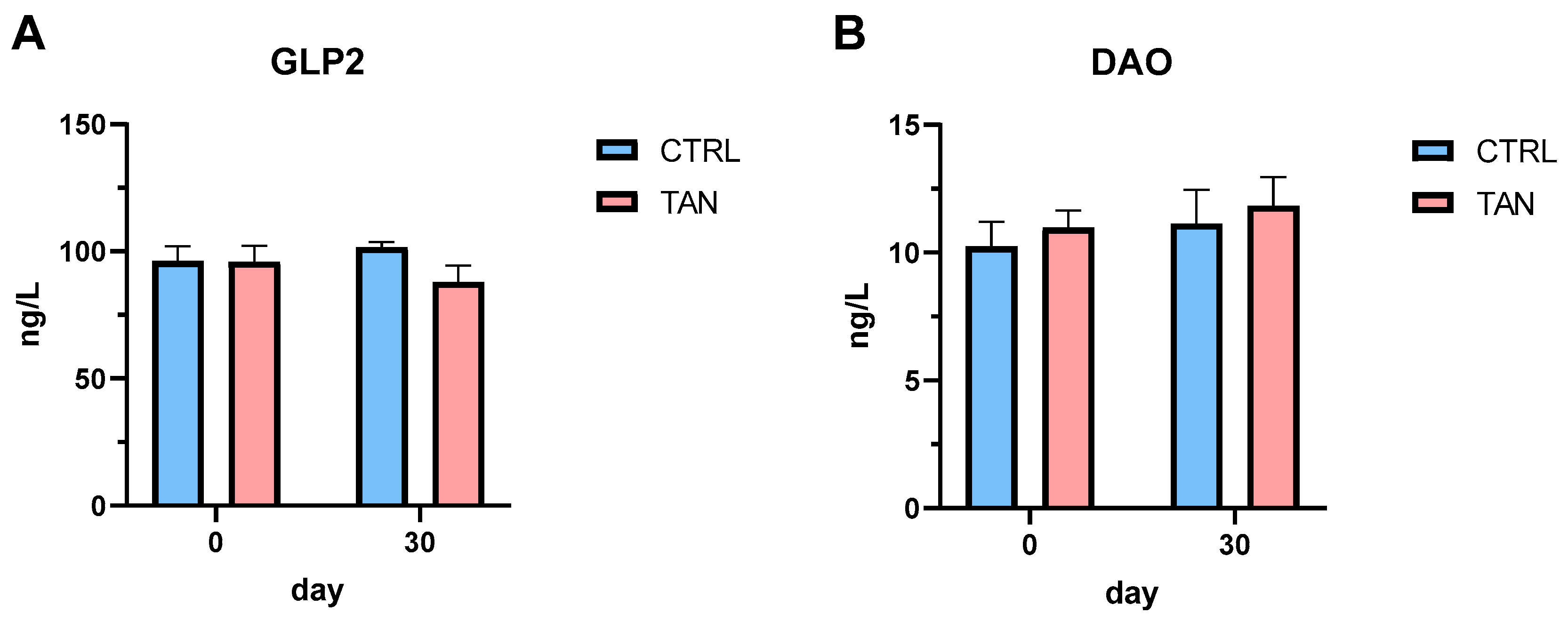
3.4. Serum Concentration of GLP2 and DAO
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Schinwald, M.; Creutzinger, K.; Keunen, A.; Winder, C.; Haley, D.; Renaud, D. Predictors of diarrhea, mortality, and weight gain in male dairy calves. J. Dairy Sci. 2022, 105, 5296–5309. [Google Scholar] [CrossRef]
- Heinrichs, A.J.; Heinrichs, B. A prospective study of calf factors affecting first-lactation and lifetime milk production and age of cows when removed from the herd. J. Dairy Sci. 2011, 94, 336–341. [Google Scholar] [CrossRef] [PubMed]
- Cho, Y.-i.; Yoon, K.-J. An overview of calf diarrhea-infectious etiology, diagnosis, and intervention. J. Vet. Sci. 2014, 15, 1–17. [Google Scholar] [CrossRef]
- Brunauer, M.; Roch, F.-F.; Conrady, B. Prevalence of worldwide neonatal calf diarrhoea caused by bovine rotavirus in combination with Bovine Coronavirus, Escherichia coli K99 and Cryptosporidium spp.: A meta-analysis. Animals 2021, 11, 1014. [Google Scholar] [CrossRef] [PubMed]
- Santín, M.; Trout, J.M.; Fayer, R. A longitudinal study of cryptosporidiosis in dairy cattle from birth to 2 years of age. Vet. Parasitol. 2008, 155, 15–23. [Google Scholar] [CrossRef]
- Lassen, B.; Viltrop, A.; Raaperi, K.; Järvis, T. Eimeria and Cryptosporidium in Estonian dairy farms in regard to age, species, and diarrhoea. Vet. Parasitol. 2009, 166, 212–219. [Google Scholar] [CrossRef] [PubMed]
- Zahedi, A.; Ryan, U. Cryptosporidium–an update with an emphasis on foodborne and waterborne transmission. Res. Vet. Sci. 2020, 132, 500–512. [Google Scholar] [CrossRef]
- Garcia, L.S.; Arrowood, M.; Kokoskin, E.; Paltridge, G.P.; Pillai, D.R.; Procop, G.W.; Ryan, N.; Shimizu, R.Y.; Visvesvara, G. Practical guidance for clinical microbiology laboratories: Laboratory diagnosis of parasites from the gastrointestinal tract. Clin. Microbiol. Rev. 2018, 31, e00025-17. [Google Scholar] [CrossRef]
- Quilez, J.; Sanchez-Acedo, C.; Del Cacho, E.; Clavel, A.; Causape, A. Prevalence of Cryptosporidium and Giardia infections in cattle in Aragon (northeastern Spain). Vet. Parasitol. 1996, 66, 139–146. [Google Scholar] [CrossRef]
- Weyl-Feinstein, S.; Markovics, A.; Eitam, H.; Orlov, A.; Yishay, M.; Agmon, R.; Miron, J.; Izhaki, I.; Shabtay, A. effect of pomegranate-residue supplement on Cryptosporidium parvum oocyst shedding in neonatal calves. J. Dairy Sci. 2014, 97, 5800–5805. [Google Scholar] [CrossRef]
- Al Mawly, J. Epidemiological Studies of Enteropathogens of Newborn Calves in New Zealand Dairy Farms. Ph.D. Thesis, Massey University, Palmerston North, New Zealand.
- Regulation (EU) 2019/6 of the European Parliament and of the Council of 11 December 2018 on Veterinary Medicinal Products and Repealing Directive 2001/82/EC (Text with EEA Relevance); European Parliament: Strasbourg, France, 2018.
- Bobeck, E.A. Nutrition and health: Companion animal applications: Functional nutrition in Livestock and Companion Animals to Modulate the Immune Response. J. Anim. Sci. 2020, 98, skaa035. [Google Scholar] [CrossRef]
- Jouany, J.-P.; Morgavi, D. Use of ‘natural’ products as alternatives to antibiotic feed additives in ruminant production. Animal 2007, 1, 1443–1466. [Google Scholar] [CrossRef]
- Silva, V.; Falco, V.; Dias, M.I.; Barros, L.; Silva, A.; Capita, R.; Alonso-Calleja, C.; Amaral, J.S.; Igrejas, G.; CFR Ferreira, I. Evaluation of the phenolic profile of Castanea sativa Mill. by-products and their antioxidant and antimicrobial activity against multiresistant bacteria. Antioxidants 2020, 9, 87. [Google Scholar] [CrossRef]
- Caprarulo, V.; Giromini, C.; Rossi, L. Chestnut and quebracho tannins in pig nutrition: The effects on performance and intestinal health. Animal 2020, 15, 100064. [Google Scholar] [CrossRef]
- Khanbabaee, K.; Van Ree, T. Tannins: Classification and definition. Nat. Prod. Rep. 2001, 18, 641–649. [Google Scholar]
- Ebrahimnejad, H.; Burkholz, T.; Jacob, C. Flavanols and proanthocyanidins. In Recent Advances in Redox Active Plant and Microbial Products; Springer: Berlin/Heidelberg, Germany, 2014; pp. 211–232. [Google Scholar]
- Wu, M.; Hou, J.; Yang, Z.; Wang, A.; Wang, Q.; Wu, J.; Miao, L. Enhanced degradation of methionine by eco-friendly and low-cost tannin extract modified Fenton oxidation: Promotion of the Fe (III)/Fe (II) cycle. Chem. Eng. J. 2023, 476, 146656. [Google Scholar] [CrossRef]
- Peng, K.; Lv, X.; Zhao, H.; Chen, B.; Chen, X.; Huang, W. Antioxidant and intestinal recovery function of condensed tannins in Lateolabrax maculatus responded to in vivo and in vitro oxidative stress. Aquaculture 2022, 547, 737399. [Google Scholar] [CrossRef]
- Gourlay, G.; Constabel, C.P. Condensed tannins are inducible antioxidants and protect hybrid poplar against oxidative stress. Tree Physiol. 2019, 39, 345–355. [Google Scholar] [CrossRef] [PubMed]
- de Veras, B.O.; da Silva, M.V.; Ribeiro, P.P.C. Tannic acid is a gastroprotective that regulates inflammation and oxidative stress. Food Chem. Toxicol. 2021, 156, 112482. [Google Scholar] [CrossRef] [PubMed]
- Mhlanga, P.; Perumal, P.O.; Somboro, A.M.; Amoako, D.G.; Khumalo, H.M.; Khan, R.B. Mechanistic insights into oxidative stress and apoptosis mediated by tannic acid in human liver hepatocellular carcinoma cells. Int. J. Mol. Sci. 2019, 20, 6145. [Google Scholar] [CrossRef]
- Lourenço, S.C.; Moldão-Martins, M.; Alves, V.D. Antioxidants of natural plant origins: From sources to food industry applications. Molecules 2019, 24, 4132. [Google Scholar] [CrossRef]
- Rahaman, M.M.; Hossain, R.; Herrera--Bravo, J.; Islam, M.T.; Atolani, O.; Adeyemi, O.S.; Owolodun, O.A.; Kambizi, L.; Daştan, S.D.; Calina, D. Natural antioxidants from some fruits, seeds, foods, natural products, and associated health benefits: An update. Food Sci. Nutr. 2023, 11, 1657–1670. [Google Scholar] [CrossRef] [PubMed]
- Dong, G.; Liu, H.; Yu, X.; Zhang, X.; Lu, H.; Zhou, T.; Cao, J. Antimicrobial and anti-biofilm activity of tannic acid against Staphylococcus aureus. Nat. Prod. Res. 2018, 32, 2225–2228. [Google Scholar] [CrossRef]
- Alshaibani, D.; Zhang, R.; Wu, V.C. Antibacterial characteristics and activity of Vaccinium macrocarpon proanthocyanidins against diarrheagenic Escherichia coli. J. Funct. Foods 2017, 39, 133–138. [Google Scholar] [CrossRef]
- Fraga-Corral, M.; Otero, P.; Echave, J.; Garcia-Oliveira, P.; Carpena, M.; Jarboui, A.; Nuñez-Estevez, B.; Simal-Gandara, J.; Prieto, M.A. By-Products of Agri-Food Industry as Tannin-Rich Sources: A Review of Tannins’ Biological Activities and Their Potential for Valorization. Foods 2021, 10, 137. [Google Scholar] [CrossRef]
- Bonelli, F.; Turini, L.; Sarri, G.; Serra, A.; Buccioni, A.; Mele, M. Oral administration of chestnut tannins to reduce the duration of neonatal calf diarrhea. BMC Vet. Res. 2018, 14, 227. [Google Scholar] [CrossRef] [PubMed]
- Rivera-Méndez, C.; Plascencia, A.; Torrentera, N.; Zinn, R. Effect of level and source of supplemental tannin on growth performance of steers during the late finishing phase. J. Appl. Anim. Res. 2017, 45, 199–203. [Google Scholar] [CrossRef]
- Krueger, W.; Gutierrez-Bañuelos, H.; Carstens, G.E.; Min, B.; Pinchak, W.; Gomez, R.; Anderson, R.; Krueger, N.; Forbes, T. Effects of dietary tannin source on performance, feed efficiency, ruminal fermentation, and carcass and non-carcass traits in steers fed a high-grain diet. Anim. Feed Sci. Technol. 2010, 159, 1–9. [Google Scholar] [CrossRef]
- Demarco, C.F.; Paisley, S.; Goodall, R.; Brauner, C.C.; Lake, S. Effects of bacterial DFM and tannins on measures of immunity and growth performance of newly weaned beef calves. Livest. Sci. 2021, 250, 104571. [Google Scholar] [CrossRef]
- Serri, M.; Mahdavi, A.; Riasi, A.; Hashemzadeh, F.; Kowsar, R. The addition of hydrolyzable tannin extract to milk affects calves’ performance, health, blood metabolites, and pathogen shedding. Anim. Feed Sci. Technol. 2022, 292, 115451. [Google Scholar] [CrossRef]
- Soleiman, P.; Kheiri, F. The effect of different levels of tannic acid on some performance traits in holstein dairy calves. Iran. J. Appl. Anim. Sci. 2018, 8, 19–23. [Google Scholar]
- Hassan, Z.M.; Manyelo, T.G.; Selaledi, L.; Mabelebele, M. The effects of tannins in monogastric animals with special reference to alternative feed ingredients. Molecules 2020, 25, 4680. [Google Scholar] [CrossRef]
- Aboagye, I.A.; Beauchemin, K.A. Potential of molecular weight and structure of tannins to reduce methane emissions from ruminants: A review. Animals 2019, 9, 856. [Google Scholar] [CrossRef] [PubMed]
- Caprarulo, V.; Hejna, M.; Giromini, C.; Liu, Y.; Dell’Anno, M.; Sotira, S.; Reggi, S.; Sgoifo-Rossi, C.A.; Callegari, M.L.; Rossi, L. Evaluation of Dietary Administration of Chestnut and Quebracho Tannins on Growth, Serum Metabolites and Fecal Parameters of Weaned Piglets. Animals 2020, 10, 1945. [Google Scholar] [CrossRef] [PubMed]
- Directive 2010/63/EU of the European Parliament and of the Council of 22 September 2010 on the Protection of Animals Used for Scientific Purposes. Text with EEA Relevance; European Parliament: Strasbourg, France, 2010.
- Directive 2008/119/EC of 18 December 2008 Laying Down Minimum Standards for the Protection of Calves; European Parliament: Strasbourg, France, 2008.
- Adejoro, F.A.; Hassen, A.; Akanmu, A.M. Effect of lipid-encapsulated acacia tannin extract on feed intake, nutrient digestibility and methane emission in sheep. Animals 2019, 9, 863. [Google Scholar] [CrossRef]
- Amaral-Phillips, D.M.; Scharko, P.B.; Johns, J.T.; Franklin, S. Feeding and Managing Baby Calves from Birth to 3 Months of Age. UK Cooperative Extension Service, University of Kentucky, ASC-161. 2006. Available online: https://www2.ca.uky.edu/agcomm/pubs/asc/asc161/ASC161.PDF (accessed on 18 January 2024).
- Santos, F.; De Paula, M.; Lezier, D.; Silva, J.; Santos, G.; Bittar, C. Essential oils for dairy calves: Effects on performance, scours, rumen fermentation and intestinal fauna. Animal 2015, 9, 958–965. [Google Scholar] [CrossRef]
- Gomez, D.; Arroyo, L.; Costa, M.; Viel, L.; Weese, J. Characterization of the fecal bacterial microbiota of healthy and diarrheic dairy calves. J. Vet. Intern. Med. 2017, 31, 928–939. [Google Scholar] [CrossRef]
- Frazzini, S.; Scaglia, E.; Dell’Anno, M.; Reggi, S.; Panseri, S.; Giromini, C.; Lanzoni, D.; Sgoifo Rossi, C.A.; Rossi, L. Antioxidant and Antimicrobial Activity of Algal and Cyanobacterial Extracts: An In Vitro Study. Antioxidants 2022, 11, 992. [Google Scholar] [CrossRef]
- AOAC. Official Methods of Analysis, 21st ed.; Association of Official Analytical Chemists: Washington, DC, USA, 2019. [Google Scholar]
- Kelln, B.; Penner, G.B.; Acharya, S.N.; McAllister, T.A.; Lardner, H.A. Impact of condensed tannin-containing legumes on ruminal fermentation, nutrition, and performance in ruminants: A review. Can. J. Anim. Sci. 2020, 101, 210–223. [Google Scholar] [CrossRef]
- Addisu, S. Effect of dietary tannin source feeds on ruminal fermentation and production of cattle; a review. Online J. Anim. Feed Res. 2016, 6, 45–56. [Google Scholar]
- Orzuna-Orzuna, J.F.; Dorantes-Iturbide, G.; Lara-Bueno, A.; Mendoza-Martínez, G.D.; Miranda-Romero, L.A.; Hernández-García, P.A. Effects of dietary tannins’ supplementation on growth performance, rumen fermentation, and enteric methane emissions in beef cattle: A meta-analysis. Sustainability 2021, 13, 7410. [Google Scholar] [CrossRef]
- Reggi, S.; Giromini, C.; Dell’Anno, M.; Baldi, A.; Rebucci, R.; Rossi, L. In Vitro Digestion of Chestnut and Quebracho Tannin Extracts: Antimicrobial Effect, Antioxidant Capacity and Cytomodulatory Activity in Swine Intestinal IPEC-J2 Cells. Animals 2020, 10, 195. [Google Scholar] [CrossRef] [PubMed]
- Hoffmann, A.; Görlich, S.; Steingass, H.; Terry, H.; Schollenberger, M.; Hartung, K.; Mosenthin, R. Milk production and milk fatty acids in dairy cows fed crushed rapeseed or rapeseed oil. Livest. Sci. 2016, 190, 31–34. [Google Scholar] [CrossRef]
- Franceschi, P.; Malacarne, M.; Bortolazzo, E.; Coloretti, F.; Formaggioni, P.; Garavaldi, A.; Musi, V.; Summer, A. Automatic milking systems in the production of Parmigiano Reggiano cheese: Effects on the milk quality and on cheese characteristics. Agriculture 2022, 12, 104. [Google Scholar] [CrossRef]
- Malacarne, M.; Summer, A.; Fossa, E.; Formaggioni, P.; Franceschi, P.; Pecorari, M.; Mariani, P. Composition, coagulation properties and Parmigiano-Reggiano cheese yield of Italian Brown and Italian Friesian herd milks. J. Dairy Res. 2006, 73, 171–177. [Google Scholar] [CrossRef]
- Bonelli, F.; Turini, L.; Paganelli, A.; Conte, G.; Meucci, V.; Sgorbini, M.; Buccioni, A.; Mele, M. Evaluation of oral administration of chestnut tannins in preventing calf diarrhoea. Ital. J. Anim. Sci. 2021, 20, 640–649. [Google Scholar] [CrossRef]
- Oliveira, R.; Narciso, C.; Bisinotto, R.; Perdomo, M.; Ballou, M.; Dreher, M.; Santos, J. Effects of feeding polyphenols from pomegranate extract on health, growth, nutrient digestion, and immunocompetence of calves. J. Dairy Sci. 2010, 93, 4280–4291. [Google Scholar] [CrossRef]
- Smith, D.R. Field disease diagnostic investigation of neonatal calf diarrhea. Vet. Clin. Food Anim. Pract. 2012, 28, 465–481. [Google Scholar] [CrossRef]
- Muktar, Y.; Mamo, G.; Tesfaye, B.; Belina, D. A review on major bacterial causes of calf diarrhea and its diagnostic method. J. Vet. Med. Anim. Health 2015, 7, 173–185. [Google Scholar]
- Nydam, D.V.; Wade, S.E.; Schaaf, S.L.; Mohammed, H.O. Number of Cryptosporidium parvum oocysts or Giardia spp cysts shed by dairy calves after natural infection. Am. J. Vet. Res. 2001, 62, 1612–1615. [Google Scholar] [CrossRef] [PubMed]
- Zambriski, J.A.; Nydam, D.V.; Bowman, D.D.; Bellosa, M.L.; Burton, A.J.; Linden, T.C.; Liotta, J.L.; Ollivett, T.L.; Tondello-Martins, L.; Mohammed, H.O. Description of fecal shedding of Cryptosporidium parvum oocysts in experimentally challenged dairy calves. Parasitol. Res. 2013, 112, 1247–1254. [Google Scholar] [CrossRef]
- Delafosse, A.; Chartier, C.; Dupuy, M.-C.; Dumoulin, M.; Pors, I.; Paraud, C. Cryptosporidium parvum infection and associated risk factors in dairy calves in western France. Prev. Vet. Med. 2015, 118, 406–412. [Google Scholar] [CrossRef]
- Álvarez, P.; Alvarado, C.; Puerto, M.; Schlumberger, A.; Jiménez, L.; De la Fuente, M. Improvement of leukocyte functions in prematurely aging mice after five weeks of diet supplementation with polyphenol-rich cereals. Nutrition 2006, 22, 913–921. [Google Scholar] [CrossRef]
- Verhelst, R.; Schroyen, M.; Buys, N.; Niewold, T. Dietary polyphenols reduce diarrhea in enterotoxigenic Escherichia coli (ETEC) infected post-weaning piglets. Livest. Sci. 2014, 160, 138–140. [Google Scholar] [CrossRef]
- Sung, S.H.; Kim, K.H.; Jeon, B.T.; Cheong, S.H.; Park, J.H.; Kim, D.H.; Kweon, H.J.; Moon, S.H. Antibacterial and antioxidant activities of tannins extracted from agricultural by-products. J. Med. Plants Res. 2012, 6, 3072–3079. [Google Scholar] [CrossRef]
- Fu, Z.; Yang, Y.; Ma, L.; Malmuthuge, N.; Guan, L.; Bu, D. Dynamics of oxidative stress and immune responses in neonatal calves during diarrhea. J. Dairy Sci. 2023, 107, 1286–1298. [Google Scholar] [CrossRef]
- Yanar, K.E.; Eren, E.; Aktaş, M.S.; Eroğlu, M.S.; Kandemir, Ö.; Aydın, G. Prognostic potential of inflammatory markers, oxidative status, thrombocyte indices, and renal biochemical markers in neonatal calf diarrhoea-induced systemic inflammatory response syndrome. Vet. Immunol. Immunopathol. 2023, 265, 110680. [Google Scholar] [CrossRef]
- Kumar, S.; Jakhar, K. Haematological, biochemical and oxidative stress parameter as prognostic indicators in calf diarrhoea. Pharma Innov. J. 2020, 9, 10–13. [Google Scholar]
- Farha, A.K.; Yang, Q.-Q.; Kim, G.; Li, H.-B.; Zhu, F.; Liu, H.-Y.; Gan, R.-Y.; Corke, H. Tannins as an alternative to antibiotics. Food Biosci. 2020, 38, 100751. [Google Scholar] [CrossRef]
- Kaczmarek, B. Tannic acid with antiviral and antibacterial activity as a promising component of biomaterials—A minireview. Materials 2020, 13, 3224. [Google Scholar] [CrossRef] [PubMed]
- Ekambaram, S.P.; Perumal, S.S.; Balakrishnan, A. Scope of hydrolysable tannins as possible antimicrobial agent. Phytother. Res. 2016, 30, 1035–1045. [Google Scholar] [CrossRef] [PubMed]
- Turini, L.; Mantino, A.; Tozzi, B.; Bonelli, F.; Silvi, A.; Mele, M.; Sgorbini, M.; Meucci, V.; Minieri, S. Effect of a Phytogenic Feed Additive in Preventing Calves’ Diarrhea. Front. Vet. Sci. 2022, 9, 873194. [Google Scholar] [CrossRef] [PubMed]
- Gupta, N.; Rajora, V.; Gupta, K.K.; Arora, N. Evaluation of novel polyherbal formulation for the clinico-therapeutic management of diarrhea in calves. J. Entomol. Zool. Stud. 2020, 8, 193–199. [Google Scholar]
- Landau, S.; Azaizeh, H.; Muklada, H.; Glasser, T.; Ungar, E.; Baram, H.; Abbas, N.; Markovics, A. Anthelmintic activity of Pistacia lentiscus foliage in two Middle Eastern breeds of goats differing in their propensity to consume tannin-rich browse. Vet. Parasitol. 2010, 173, 280–286. [Google Scholar] [CrossRef]
- Markovics, A.; Cohen, I.; Muklada, H.; Glasser, T.; Dvash, L.; Ungar, E.; Azaizeh, H.; Landau, S. Consumption of Pistacia lentiscus foliage alleviates coccidiosis in young goats. Vet. Parasitol. 2012, 186, 165–169. [Google Scholar] [CrossRef]
- Burke, J.; Miller, J.; Terrill, T.; Orlik, S.; Acharya, M.; Garza, J.; Mosjidis, J. Sericea lespdeza as an aid in the control of Emeria spp. in lambs. Vet. Parasitol. 2013, 193, 39–46. [Google Scholar] [CrossRef]
- Bhatta, R.; Uyeno, Y.; Tajima, K.; Takenaka, A.; Yabumoto, Y.; Nonaka, I.; Enishi, O.; Kurihara, M. Difference in the nature of tannins on in vitro ruminal methane and volatile fatty acid production and on methanogenic archaea and protozoal populations. J. Dairy Sci. 2009, 92, 5512–5522. [Google Scholar] [CrossRef]
- Benchaar, C.; McAllister, T.; Chouinard, P. Digestion, ruminal fermentation, ciliate protozoal populations, and milk production from dairy cows fed cinnamaldehyde, quebracho condensed tannin, or Yucca schidigera saponin extracts. J. Dairy Sci. 2008, 91, 4765–4777. [Google Scholar] [CrossRef]
- Vasta, V.; Yáñez-Ruiz, D.R.; Mele, M.; Serra, A.; Luciano, G.; Lanza, M.; Biondi, L.; Priolo, A. Bacterial and protozoal communities and fatty acid profile in the rumen of sheep fed a diet containing added tannins. Appl. Environ. Microbiol. 2010, 76, 2549–2555. [Google Scholar] [CrossRef]
- Redondo, L.M.; Chacana, P.A.; Dominguez, J.E.; Fernandez Miyakawa, M.E. Perspectives in the use of tannins as alternative to antimicrobial growth promoter factors in poultry. Front. Microbiol. 2014, 5, 118. [Google Scholar] [CrossRef]
- Dell’Anno, M.; Reggi, S.; Caprarulo, V.; Hejna, M.; Sgoifo Rossi, C.A.; Callegari, M.L.; Baldi, A.; Rossi, L. Evaluation of Tannin Extracts, Leonardite and Tributyrin Supplementation on Diarrhoea Incidence and Gut Microbiota of Weaned Piglets. Animals 2021, 11, 1693. [Google Scholar] [CrossRef]
- Khan, D.; Vasu, S.; Moffett, R.C.; Irwin, N.; Flatt, P.R. Differential expression of glucagon-like peptide-2 (GLP-2) is involved in pancreatic islet cell adaptations to stress and beta-cell survival. Peptides 2017, 95, 68–75. [Google Scholar] [CrossRef] [PubMed]
- Hatew, B.; Inabu, Y.; Sugino, T.; Steele, M. Effects of pulse-dose ruminal infusion of butyrate on plasma glucagon-like peptide 1 and 2 concentrations in dairy calves. J. Dairy Sci. 2019, 102, 2254–2265. [Google Scholar] [CrossRef]
- García-Martín, E.; Ayuso, P.; Martínez, C.; Agúndez, J.A. Improved analytical sensitivity reveals the occurrence of gender-related variability in diamine oxidase enzyme activity in healthy individuals. Clin. Biochem. 2007, 40, 1339–1341. [Google Scholar] [CrossRef] [PubMed]
- Dell’Anno, M.; Scaglia, E.; Reggi, S.; Grossi, S.; Rossi, C.A.S.; Frazzini, S.; Caprarulo, V.; Rossi, L. Evaluation of tributyrin supplementation in milk replacer on diarrhoea occurrence in preweaning Holstein calves. Animal 2023, 17, 100791. [Google Scholar] [CrossRef] [PubMed]
- Thomas, V.L.; Chavez, M.; Browne, E.N.; Minnis, A.M. Instagram as a tool for study engagement and community building among adolescents: A social media pilot study. Digit. Health 2020, 6, 2055207620904548. [Google Scholar] [CrossRef]
- Locke, S. Instagram as a Tool of Diffusion for the Livestock Industry. J. Appl. Commun. 2023, 107, 1. [Google Scholar] [CrossRef]
- Muca, E.; Buonaiuto, G.; Lamanna, M.; Silvestrelli, S.; Ghiaccio, F.; Federiconi, A.; De Matos Vettori, J.; Colleluori, R.; Fusaro, I.; Raspa, F. Reaching a wider audience: Instagram’s role in dairy cow nutrition education and engagement. Animals 2023, 13, 3503. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Group | p-Values | ||||
|---|---|---|---|---|---|
| CTRL | TAN | Treatment | Time | Time × Treatment | |
| BW (kg) | 0.0417 | <0.0001 | 0.9115 | ||
| d0 | 45.30 ± 1.39 | 43.20 ± 1.39 | |||
| d14 | 45.80 ± 1.39 | 42.20 ± 1.39 | |||
| d30 | 60.80 ± 1.39 | 58.00 ± 1.39 | |||
| ADG (g/day) | 0.7979 | <0.0001 | 0.9423 | ||
| d0–14 | 35.72 ± 79.90 | 35.71 ± 79.90 | |||
| d15–30 | 937.50 ± 79.90 | 893.75 ± 79.90 | |||
| d0–30 | 516.67 ± 63.25 | 493.33 ± 63.25 | 0.7971 | ||
| CTRL | TAN | p-Value | ||
|---|---|---|---|---|
| Frequency of diarrhoea (%) | d0 | 10.00 | 0.00 | 0.1360 |
| d3 | 30.00 a | 5.00 b | 0.0191 | |
| d7 | 36.00 | 25.00 | 0.3613 | |
| d14 | 40.00 | 30.00 | 0.3291 | |
| d30 | 20.00 | 15.00 | 0.6392 | |
| d0–30 | 27.00 a | 15.00 b | 0.0150 | |
| Moderate diarrhoea (%) | d0 | 5.00 | 0.00 | 0.3049 |
| d3 | 30.00 a | 5.00 b | 0.0191 | |
| d7 | 25.00 | 30.00 | 0.3613 | |
| d14 | 35.00 | 25.00 | 0.3613 | |
| d30 | 15.00 | 10.00 | 0.6056 | |
| d0–30 | 22.00 a | 11.00 b | 0.0193 | |
| Severe diarrhoea (%) | d0 | 5.00 | 0.00 | 0.3049 |
| d3 | 0.00 | 0.00 | - | |
| d7 | 10.00 | 10.00 | 1.0000 | |
| d14 | 5.00 | 5.00 | 1.0000 | |
| d30 | 5.00 | 5.00 | 1.0000 | |
| d0–30 | 5.00 | 4.00 | 0.7268 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Dell’Anno, M.; Frazzini, S.; Ferri, I.; Tuberti, S.; Bonaldo, E.; Botti, B.; Grossi, S.; Sgoifo Rossi, C.A.; Rossi, L. Effect of Dietary Supplementation of Chestnut and Quebracho Tannin Supplementation on Neonatal Diarrhoea in Preweaning Calves. Antioxidants 2024, 13, 237. https://doi.org/10.3390/antiox13020237
Dell’Anno M, Frazzini S, Ferri I, Tuberti S, Bonaldo E, Botti B, Grossi S, Sgoifo Rossi CA, Rossi L. Effect of Dietary Supplementation of Chestnut and Quebracho Tannin Supplementation on Neonatal Diarrhoea in Preweaning Calves. Antioxidants. 2024; 13(2):237. https://doi.org/10.3390/antiox13020237
Chicago/Turabian StyleDell’Anno, Matteo, Sara Frazzini, Irene Ferri, Susanna Tuberti, Elisa Bonaldo, Benedetta Botti, Silvia Grossi, Carlo Angelo Sgoifo Rossi, and Luciana Rossi. 2024. "Effect of Dietary Supplementation of Chestnut and Quebracho Tannin Supplementation on Neonatal Diarrhoea in Preweaning Calves" Antioxidants 13, no. 2: 237. https://doi.org/10.3390/antiox13020237
APA StyleDell’Anno, M., Frazzini, S., Ferri, I., Tuberti, S., Bonaldo, E., Botti, B., Grossi, S., Sgoifo Rossi, C. A., & Rossi, L. (2024). Effect of Dietary Supplementation of Chestnut and Quebracho Tannin Supplementation on Neonatal Diarrhoea in Preweaning Calves. Antioxidants, 13(2), 237. https://doi.org/10.3390/antiox13020237