

Optimization, Metabolomic Analysis, Antioxidant Potential and Depigmenting Activity of Polyphenolic Compounds from Unmature Ajwa Date Seeds (Phoenix dactylifera L.) Using Ultrasonic-Assisted Extraction



Abstract
1. Introduction
2. Materials and Methods
2.1. Sample Collection and Preparation
2.2. Measurement of Total Phenolic (TPC) and Total Flavonoid Content (TFC)
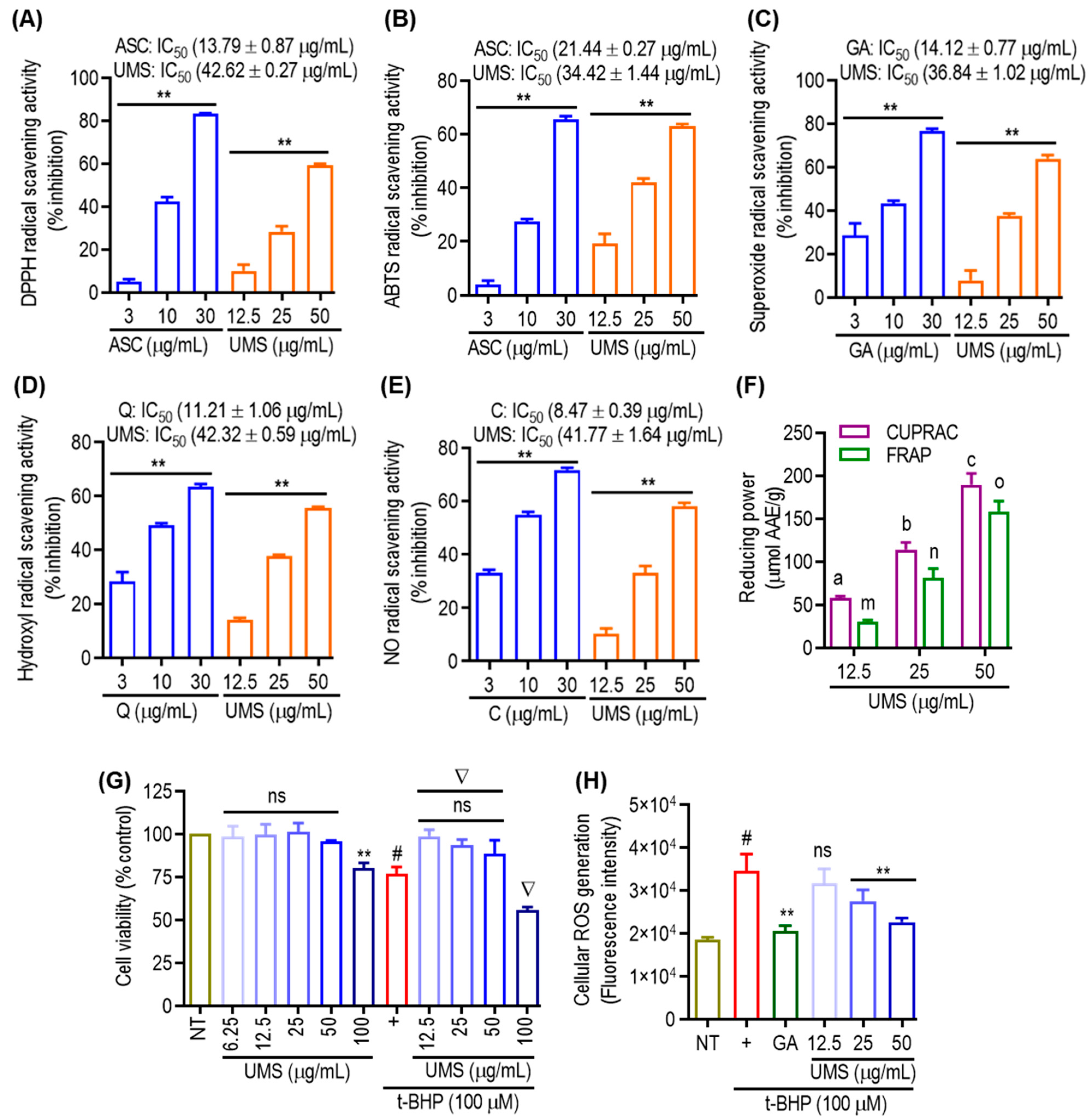
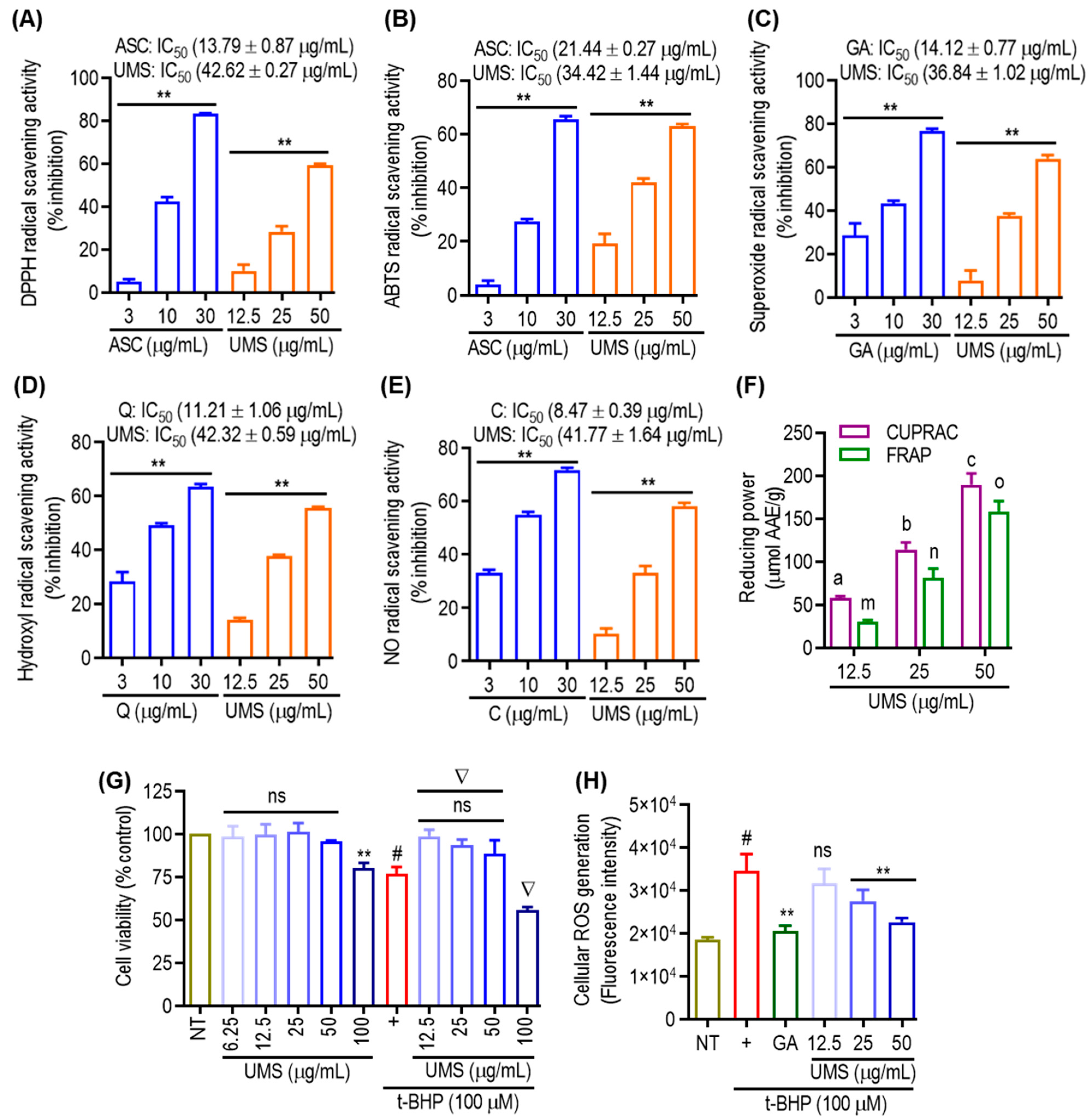
2.3. Cell-Free Antioxidant Assays
2.4. Cell Culture and Intracellular ROS Generation Assay
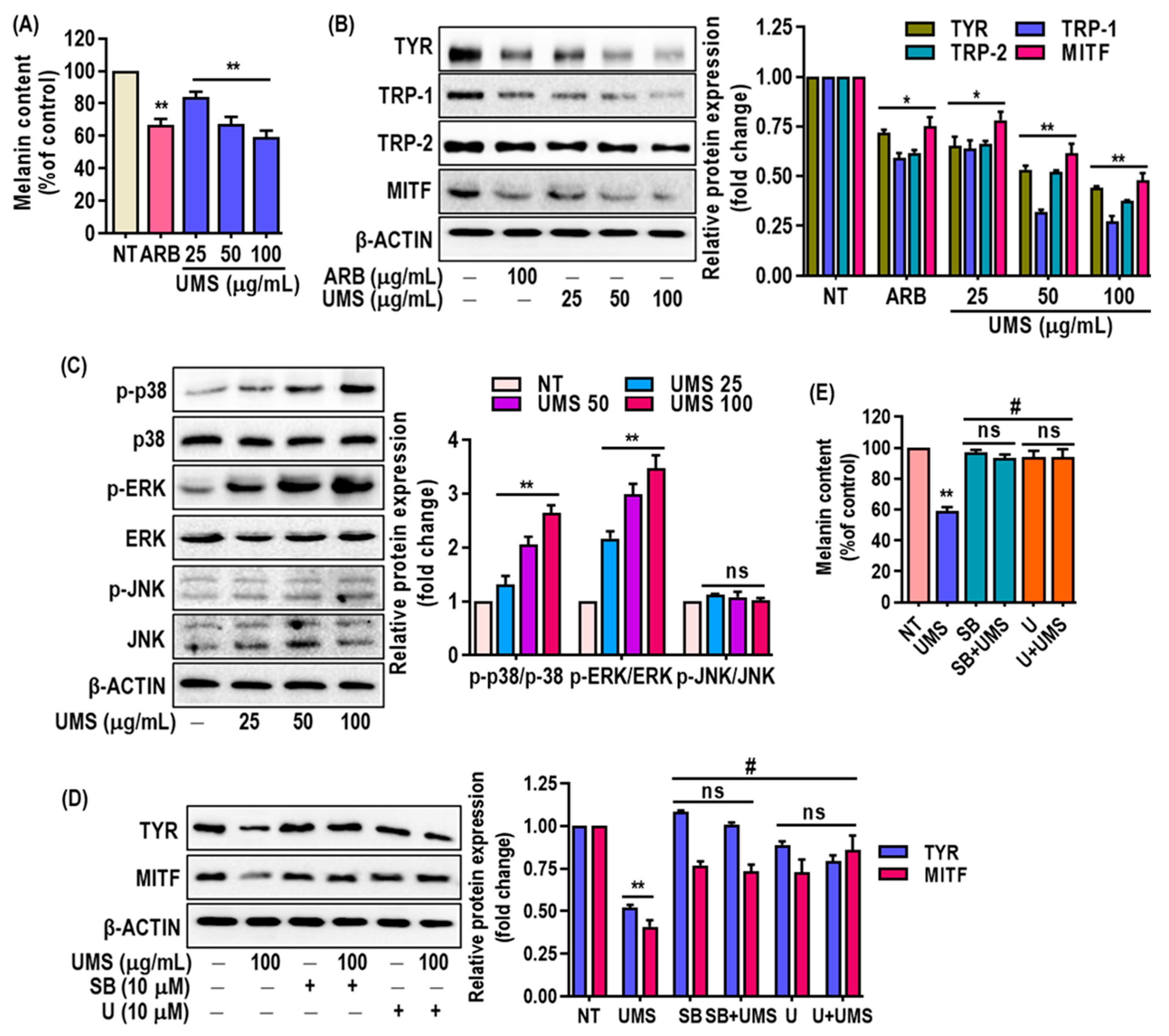
2.5. Effect of UMS on Melanin Content
2.6. Preparation of Cell Lysates and Western Blotting
2.7. Single-Factor Experiment
2.8. Experimental Design of RSM for the Extraction Process
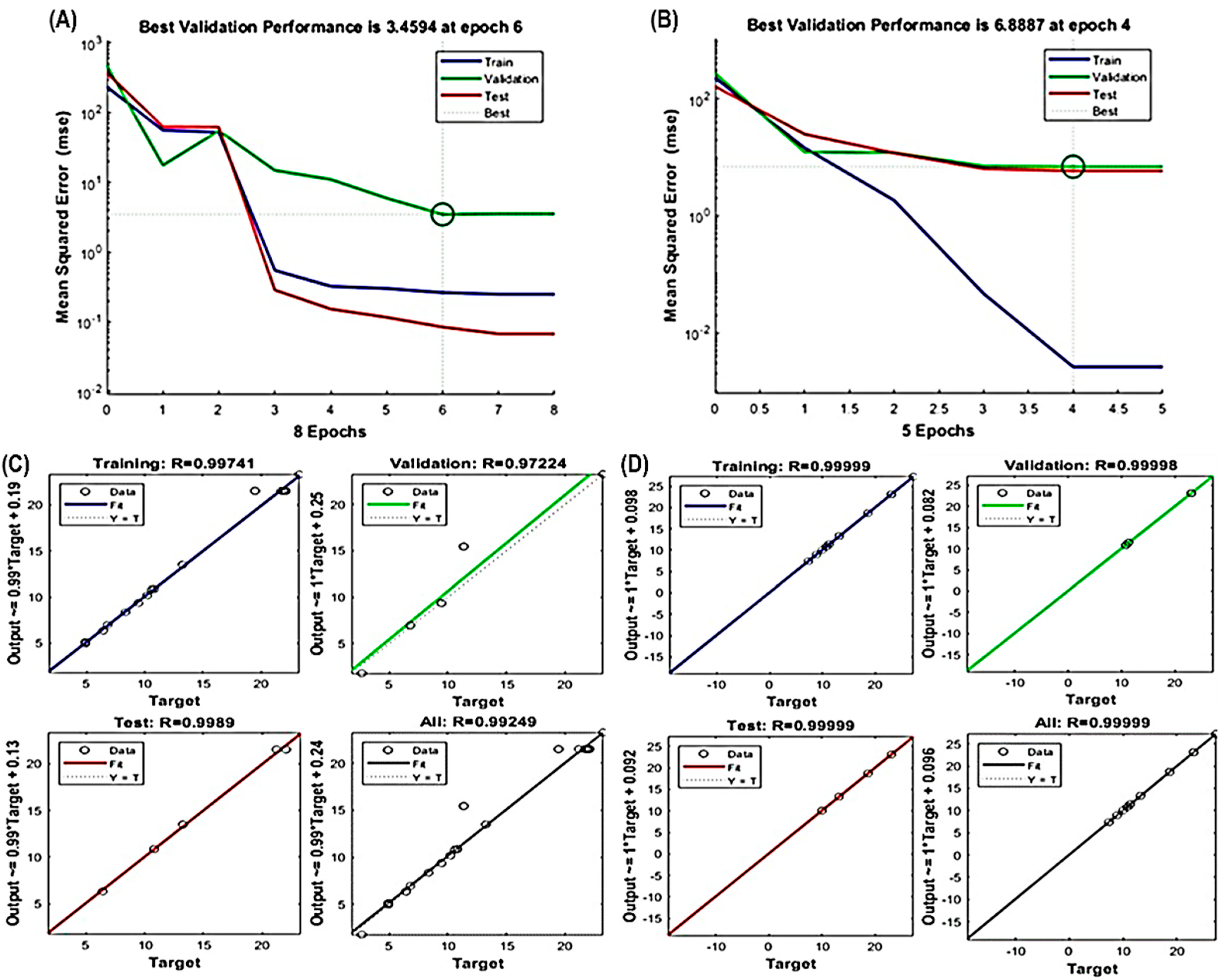
2.9. Artificial Neural Network (ANN) Modeling
2.10. Validation of the Model
2.11. Analysis of Chemical Compounds by ESI–MS/MS
2.12. Statistical Analysis
3. Results and Discussion
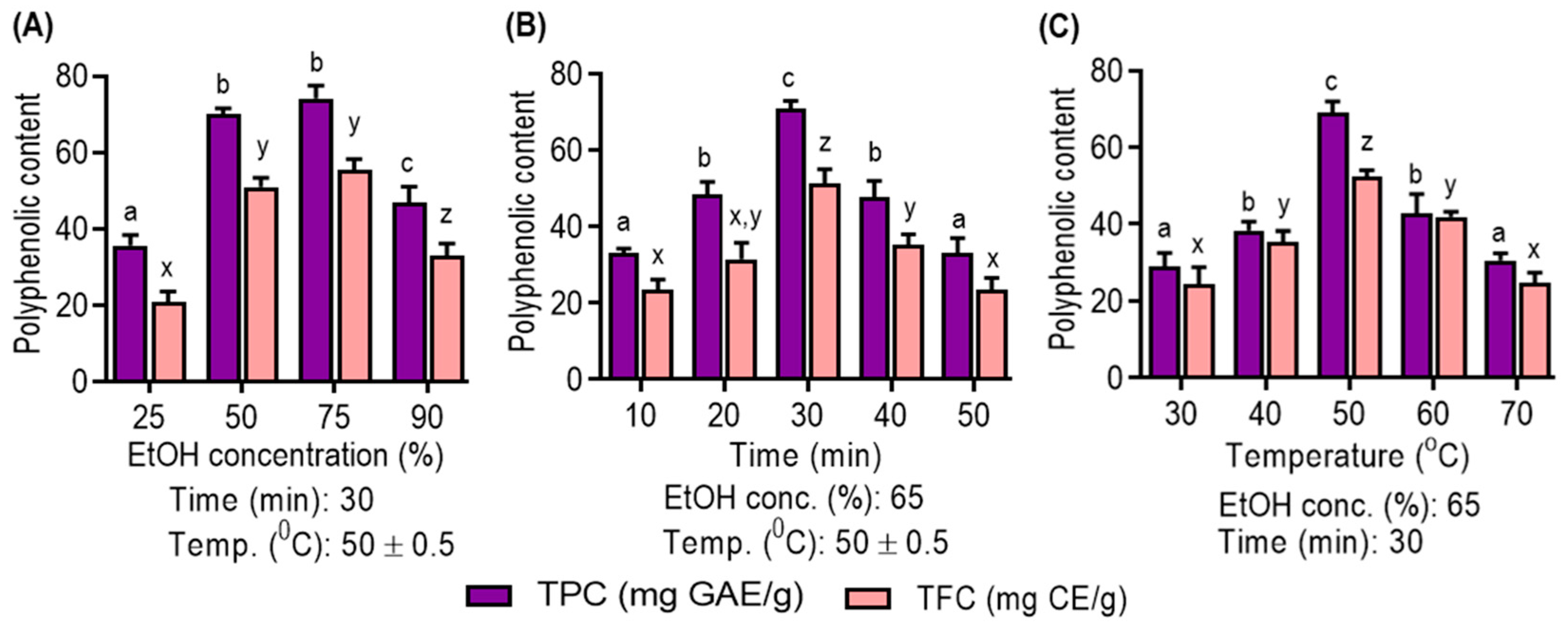
3.1. Single Factor Analysis
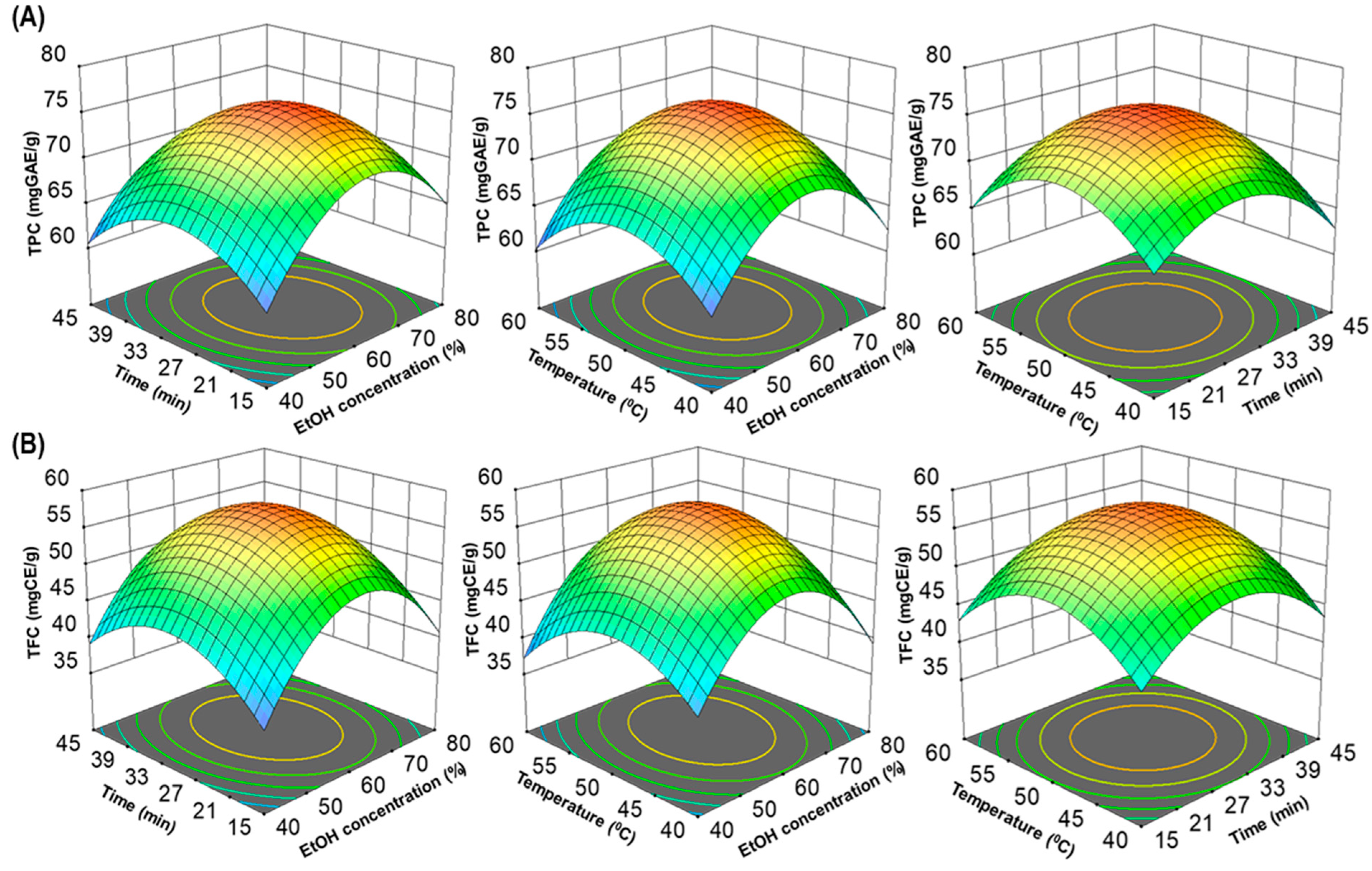
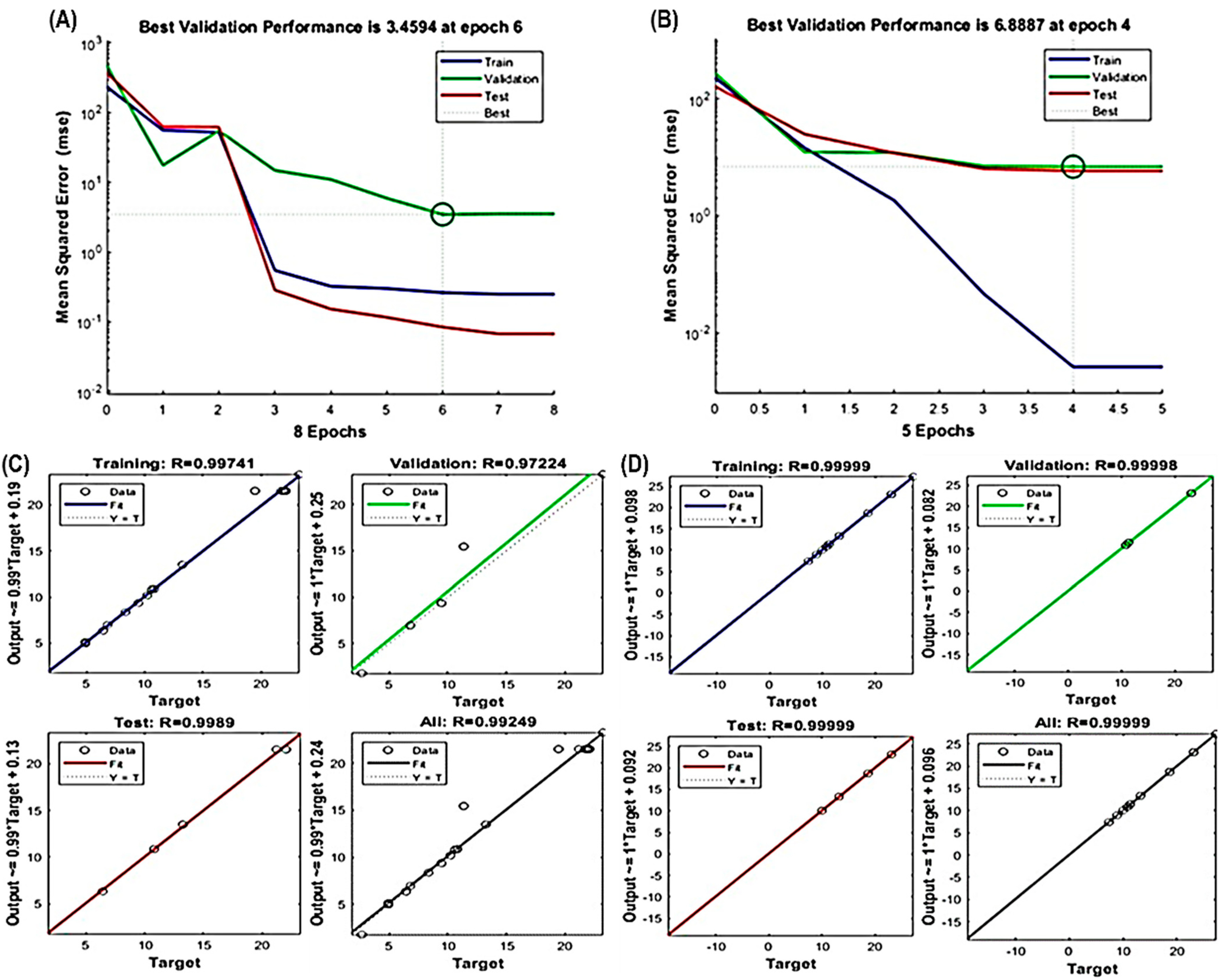
3.2. Fitting of the RSM and ANN Models
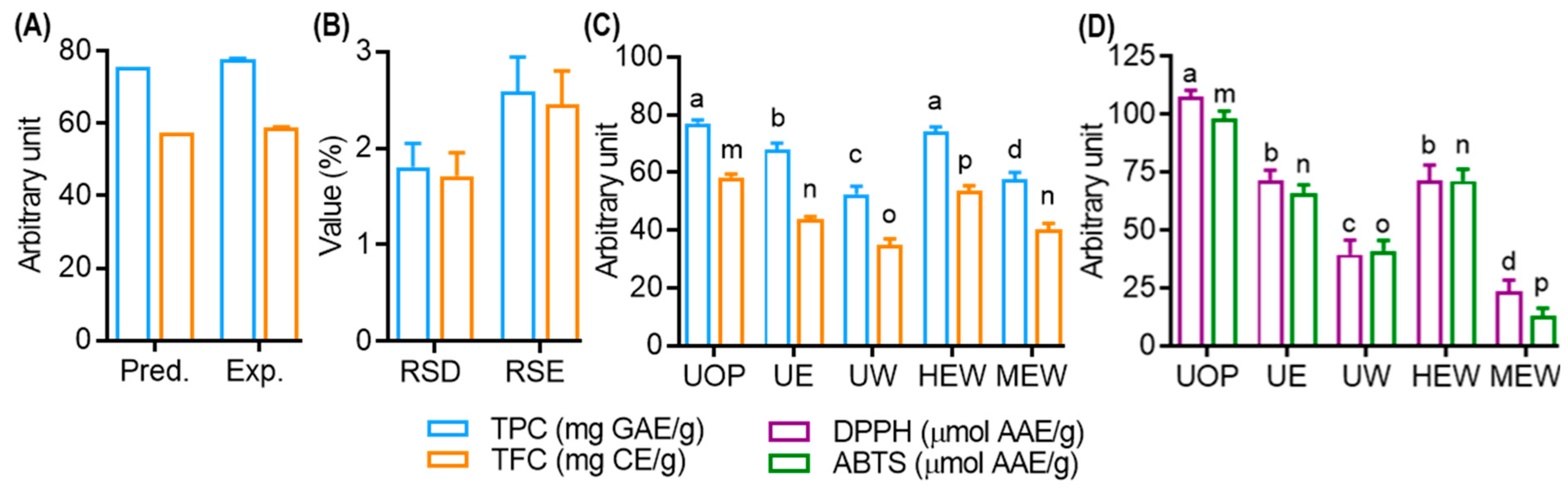
3.3. Comparison of the Prediction Abilities of the RSM and ANN Models
3.4. Model Validation and Comparison with Other Extraction Methods
3.5. Identification of Secondary Metabolites in UMS by High-Resolution Mass Spectroscopy
3.6. Antioxidant Effect of UMS
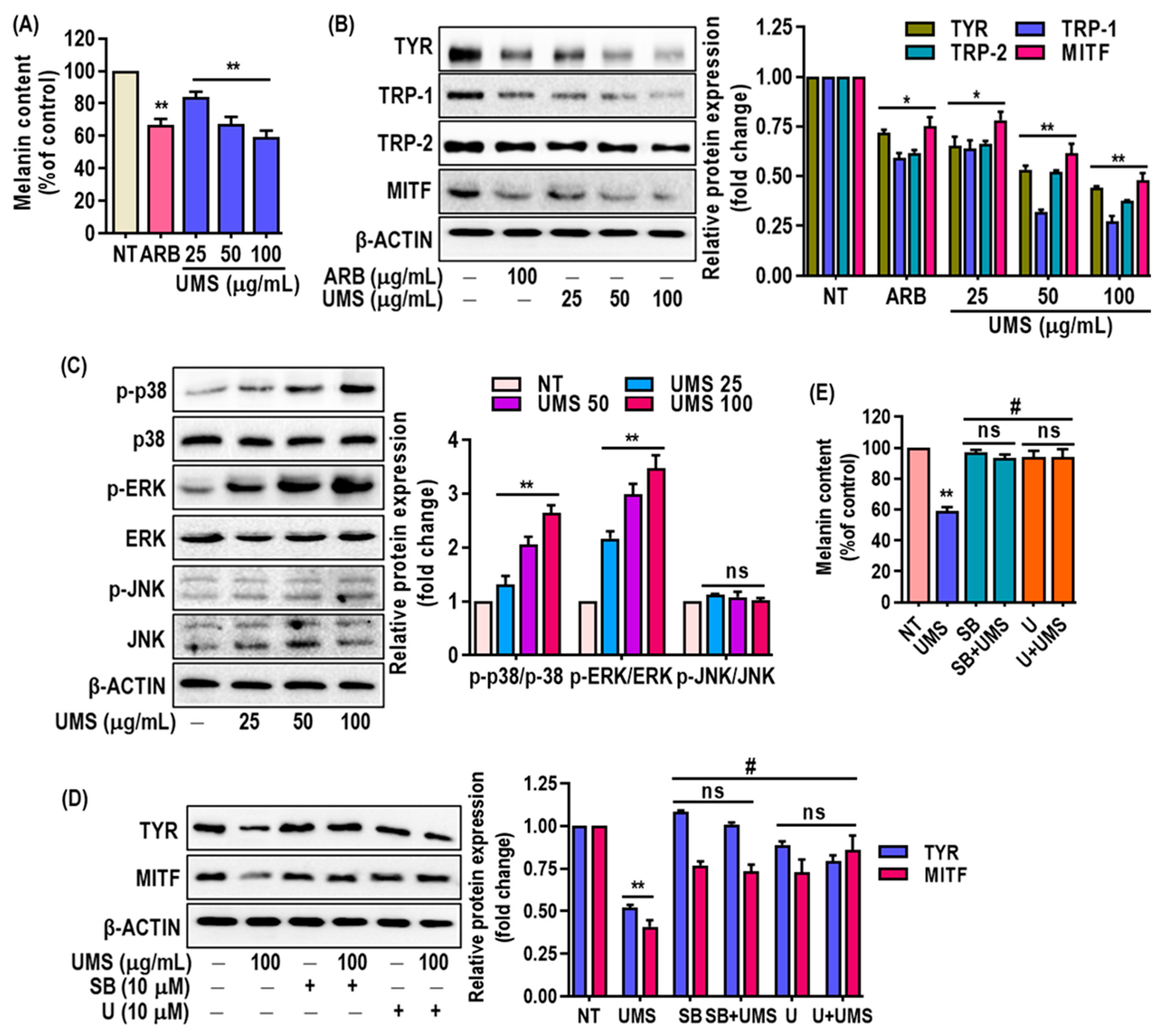
3.7. Depigmenting Effect of UMS on Hyperpigmented Melanocyte (MNT-1) Cells
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Sedraoui, S.; Badr, A.; Barba, M.G.M.; Doyen, A.; Tabka, Z.; Desjardins, Y. Optimization of the Ultrahigh-Pressure–Assisted Extraction of Phenolic Compounds and Antioxidant Activity from Palm Dates (Phoenix dactylifera L.). Food Anal. Methods 2020, 13, 1556–1569. [Google Scholar] [CrossRef]
- Galanakis, C.M. Emerging technologies for the production of nutraceuticals from agricultural by-products: A viewpoint of opportunities and challenges. Food Bioprod. Process. 2013, 91, 575–579. [Google Scholar] [CrossRef]
- Chen, M.; Zhao, Y.; Yu, S. Optimisation of ultrasonic-assisted extraction of phenolic compounds, antioxidants, and anthocyanins from sugar beet molasses. Food Chem. 2015, 172, 543–550. [Google Scholar] [CrossRef]
- Koraqi, H.; Petkoska, A.T.; Khalid, W.; Sehrish, A.; Ambreen, S.; Lorenzo, J.M. Optimization of the Extraction Conditions of Antioxidant Phenolic Compounds from Strawberry Fruits (Fragaria x ananassa Duch.) Using Response Surface Methodology. Food Anal. Methods 2023, 16, 1030–1042. [Google Scholar] [CrossRef] [PubMed]
- Bezerra, M.A.; Santelli, R.E.; Oliveira, E.P.; Villar, L.S.; Escaleira, L.A. Response surface methodology (RSM) as a tool for optimization in analytical chemistry. Talanta 2008, 76, 965–977. [Google Scholar] [CrossRef]
- Desai, K.M.; Survase, S.A.; Saudagar, P.S.; Lele, S.S.; Singhal, R.S. Comparison of artificial neural network (ANN) and response surface methodology (RSM) in fermentation media optimization: Case study of fermentative production of scleroglucan. Biochem. Eng. J. 2008, 41, 266–273. [Google Scholar] [CrossRef]
- Zobel, C.W.; Cook, D.F. Evaluation of neural network variable influence measures for process control. Eng. Appl. Artif. Intell. 2011, 24, 803–812. [Google Scholar] [CrossRef]
- Alam, M.B.; Ju, M.-K.; Lee, S.-H. DNA Protecting Activities of Nymphaea nouchali (Burm. f) Flower Extract Attenuate t-BHP-Induced Oxidative Stress Cell Death through Nrf2-Mediated Induction of Heme Oxygenase-1 Expression by Activating MAP-Kinases. Int. J. Mol. Sci. 2017, 18, 2069. [Google Scholar] [CrossRef]
- Alam, M.B.; Naznin, M.; Islam, S.; Alshammari, F.H.; Choi, H.J.; Song, B.R.; Kim, S.; Lee, S.H. High Resolution Mass Spec-troscopy-Based Secondary Metabolite Profiling of Nymphaea nouchali (Burm. f) Stem Attenuates Oxidative Stress via Regula-tion of MAPK/Nrf2/HO-1/ROS Pathway. Antioxidants 2021, 10, 719. [Google Scholar] [CrossRef]
- D’Mello, S.A.; Finlay, G.J.; Baguley, B.C.; Askarian-Amiri, M.E. Signaling Pathways in Melanogenesis. Int. J. Mol. Sci. 2016, 17, 1144. [Google Scholar] [CrossRef]
- Alam, M.B.; Ahmed, A.; Motin, M.A.; Kim, S.; Lee, S.-H. Attenuation of melanogenesis by Nymphaea nouchali (Burm. f) flower extract through the regulation of cAMP/CREB/MAPKs/MITF and proteasomal degradation of tyrosinase. Sci. Rep. 2018, 8, 13928. [Google Scholar] [CrossRef]
- Kondo, T.; Hearing, V.J. Update on the regulation of mammalian melanocyte function and skin pigmentation. Expert Rev. Dermatol. 2011, 6, 97–108. [Google Scholar] [CrossRef]
- Yasin, B.R.; El-Fawal, H.A.N.; Mousa, S.A. Date (Phoenix dactylifera) Polyphenolics and Other Bioactive Compounds: A Traditional Islamic Remedy’s Potential in Prevention of Cell Damage, Cancer Therapeutics and Beyond. Int. J. Mol. Sci. 2015, 16, 30075–30090. [Google Scholar] [CrossRef] [PubMed]
- Zhang, C.-R.; Aldosari, S.A.; Vidyasagar, P.S.P.V.; Shukla, P.; Nair, M.G. Health-benefits of date fruits produced in Saudi Arabia based on in vitro antioxidant, anti-inflammatory and human tumor cell proliferation inhibitory assays. J. Saudi Soc. Agric. Sci. 2017, 16, 287–293. [Google Scholar] [CrossRef]
- Samad, M.A.; Hashim, S.H.; Simarani, K.; Yaacob, J.S. Antibacterial Properties and Effects of Fruit Chilling and Extract Storage on Antioxidant Activity, Total Phenolic and Anthocyanin Content of Four Date Palm (Phoenix dactylifera) Cultivars. Molecules 2016, 21, 419. [Google Scholar] [CrossRef]
- Hasan, M.; Mohieldein, A. In Vivo Evaluation of Anti Diabetic, Hypolipidemic, Antioxidative Activities of Saudi Date Seed Extract on Streptozotocin Induced Diabetic Rats. J. Clin. Diagn. Res. JCDR 2016, 10, Ff06–Ff12. [Google Scholar] [CrossRef]
- Al-Yahya, M.; Raish, M.; AlSaid, M.S.; Ahmad, A.; Mothana, R.A.; Al-Sohaibani, M.; Al-Dosari, M.S.; Parvez, M.K.; Rafatullah, S. ‘Ajwa’ dates (Phoenix dactylifera L.) extract ameliorates isoproterenol-induced cardiomyopathy through downregulation of oxidative, inflammatory and apoptotic molecules in rodent model. Phytomedicine 2016, 23, 1240–1248. [Google Scholar] [CrossRef]
- Sheikh, B.Y.; Elsaed, W.M.; Samman, A.H.; Sheikh, B.Y.; Ladin, A. Ajwa dates as a protective agent against liver toxicity in rat. Eur. Sci. J. 2014, 10, 358–368. [Google Scholar]
- Khalid, S.; Khalid, N.; Khan, R.S.; Ahmed, H.; Ahmad, A. A review on chemistry and pharmacology of Ajwa date fruit and pit. Trends Food Sci. Technol. 2017, 63, 60–69. [Google Scholar] [CrossRef]
- Nematallah, K.A.; Ayoub, N.A.; Abdelsattar, E.; Meselhy, M.R.; Elmazar, M.M.; El-Khatib, A.H.; Linscheid, M.W.; Hathout, R.M.; Godugu, K.; Adel, A.; et al. Polyphenols LC-MS2 profile of Ajwa date fruit (Phoenix dactylifera L.) and their microemulsion: Potential impact on hepatic fibrosis. J. Funct. Foods 2018, 49, 401–411. [Google Scholar] [CrossRef]
- Ben-Youssef, S.; Fakhfakh, J.; Breil, C.; Abert-Vian, M.; Chemat, F.; Allouche, N. Green extraction procedures of lipids from Tunisian date palm seeds. Ind. Crops Prod. 2017, 108, 520–525. [Google Scholar] [CrossRef]
- Alshammari, F.; Alam, M.B.; Song, B.R.; Lee, S.H. Antioxidant, Tyrosinase, α-Glucosidase, and Elastase Enzyme Inhibition Activities of Optimized Unripe Ajwa Date Pulp (Phoenix dactylifera) Extracts by Response Surface Methodology. Int. J. Mol. Sci. 2023, 24, 3396. [Google Scholar] [CrossRef]
- Valipour, M.; Banihabib, M.E.; Behbahani, S.M.R. Comparison of the ARMA, ARIMA, and the autoregressive artificial neural network models in forecasting the monthly inflow of Dez dam reservoir. J. Hydrol. 2013, 476, 433–441. [Google Scholar] [CrossRef]
- Alshammari, F.; Alam, M.B.; Naznin, M.; Javed, A.; Kim, S.; Lee, S.-H. Profiling of Secondary Metabolites of Optimized Ripe Ajwa Date Pulp (Phoenix dactylifera L.) Using Response Surface Methodology and Artificial Neural Network. Pharmaceuticals 2023, 16, 319. [Google Scholar] [CrossRef]
- Zakaria, F.; Tan, J.-K.; Mohd Faudzi, S.M.; Abdul Rahman, M.B.; Ashari, S.E. Ultrasound-assisted extraction conditions optimisation using response surface methodology from Mitragyna speciosa (Korth.) Havil leaves. Ultrason. Sonochemistry 2021, 81, 105851. [Google Scholar] [CrossRef]
- Naveja, J.J.; Rico-Hidalgo, M.P.; Medina-Franco, J.L. Analysis of a large food chemical database: Chemical space, diversity, and complexity. F1000Research 2018, 77, 993. [Google Scholar] [CrossRef]
- Guijas, C.; Montenegro-Burke, J.R.; Domingo-Almenara, X.; Palermo, A.; Warth, B.; Hermann, G.; Koellensperger, G.; Huan, T.; Uritboonthai, W.; Aisporna, A.E.; et al. METLIN: A Technology Platform for Identifying Knowns and Unknowns. Anal. Chem. 2018, 90, 3156–3164. [Google Scholar] [CrossRef] [PubMed]
- Wang, F.; Liigand, J.; Tian, S.; Arndt, D.; Greiner, R.; Wishart, D.S. CFM-ID 4.0: More Accurate ESI-MS/MS Spectral Prediction and Compound Identification. Anal. Chem. 2021, 93, 11692–11700. [Google Scholar] [CrossRef]
- Ciric, A.; Krajnc, B.; Heath, D.; Ogrinc, N. Response surface methodology and artificial neural network approach for the optimization of ultrasound-assisted extraction of polyphenols from garlic. Food Chem. Toxicol. 2020, 135, 110976. [Google Scholar] [CrossRef] [PubMed]
- Xu, D.-P.; Zheng, J.; Zhou, Y.; Li, Y.; Li, S.; Li, H.-B. Ultrasound-assisted extraction of natural antioxidants from the flower of Limonium sinuatum: Optimization and comparison with conventional methods. Food Chem. 2017, 217, 552–559. [Google Scholar] [CrossRef]
- Nipornram, S.; Tochampa, W.; Rattanatraiwong, P.; Singanusong, R. Optimization of low power ultrasound-assisted extraction of phenolic compounds from mandarin (Citrus reticulata Blanco cv. Sainampueng) peel. Food Chem. 2018, 241, 338–345. [Google Scholar] [CrossRef] [PubMed]
- Huang, S.-M.; Kuo, C.-H.; Chen, C.-A.; Liu, Y.-C.; Shieh, C.-J. RSM and ANN modeling-based optimization approach for the development of ultrasound-assisted liposome encapsulation of piceid. Ultrason. Sonochemistry 2017, 36, 112–122. [Google Scholar] [CrossRef]
- Kuo, C.-H.; Liu, T.-A.; Chen, J.-H.; Chang, C.-M.J.; Shieh, C.-J. Response surface methodology and artificial neural network optimized synthesis of enzymatic 2-phenylethyl acetate in a solvent-free system. Biocatal. Agric. Biotechnol. 2014, 3, 1–6. [Google Scholar] [CrossRef]
- Derrien, M.; Badr, A.; Gosselin, A.; Desjardins, Y.; Angers, P. Optimization of a green process for the extraction of lutein and chlorophyll from spinach by-products using response surface methodology (RSM). LWT-Food Sci. Technol. 2017, 79, 170–177. [Google Scholar] [CrossRef]
- Schymanski, E.L.; Jeon, J.; Gulde, R.; Fenner, K.; Ruff, M.; Singer, H.P.; Hollender, J. Identifying Small Molecules via High Resolution Mass Spectrometry: Communicating Confidence. Environ. Sci. Technol. 2014, 48, 2097–2098. [Google Scholar] [CrossRef]
- Shen, N.; Wang, T.; Gan, Q.; Liu, S.; Wang, L.; Jin, B. Plant flavonoids: Classification, distribution, biosynthesis, and antioxidant activity. Food Chem. 2022, 383, 132531. [Google Scholar] [CrossRef]
- Liu, H.; Orjala, J.; Sticher, O.; Rali, T. Acylated Flavonol Glycosides from Leaves of Stenochlaena palustris. J. Nat. Prod. 1999, 62, 70–75. [Google Scholar] [CrossRef]
- Murakami, T.; Kohno, K.; Ninomiya, K.; Matsuda, H.; Yoshikawa, M. Medicinal foodstuffs. XXV. Hepatoprotective principle and structures of ionone glucoside, phenethyl glycoside, and flavonol oligoglycosides from young seedpods of garden peas, Pisum sativum L. Chem. Pharm. Bull. 2001, 49, 1003–1008. [Google Scholar] [CrossRef]
- Biswal, R.P.; Dandamudi, R.B.; Patnana, D.P.; Pandey, M.; Vutukuri, V. Metabolic fingerprinting of Ganoderma spp. using UHPLC-ESI-QTOF-MS and its chemometric analysis. Phytochemistry 2022, 199, 113169. [Google Scholar] [CrossRef] [PubMed]
- Wu, S.; Wu, Z.; Fu, C.; Wu, C.; Yuan, J.; Xian, X.; Gao, H. Simultaneous identification and analysis of cassane diterpenoids in Caesalpinia minax Hance by high-performance liquid chromatography with quadrupole time-of-flight mass spectrometry. J. Sep. Sci. 2015, 38, 4000–4013. [Google Scholar] [CrossRef] [PubMed]
- Tian, Q.; Schwartz, S.J. Mass spectrometry and tandem mass spectrometry of citrus limonoids. Anal. Chem. 2003, 75, 5451–5460. [Google Scholar] [CrossRef]
- Girardi, C.; Jullian, V.; Haddad, M.; Vansteelandt, M.; Cabanillas, B.J.; Kapanda, C.N.; Herent, M.F.; Quetin-Leclercq, J.; Fabre, N. Analysis and fragmentation mechanisms of hirsutinolide-type sesquiterpene lactones by ultra-high-performance liquid chromatography/electrospray ionization linear ion trap Orbitrap mass spectrometry. Rapid Commun. Mass Spectrom. RCM 2016, 30, 569–580. [Google Scholar] [CrossRef]
- Tugizimana, F.; Steenkamp, P.A.; Piater, L.A.; Dubery, I.A. Ergosterol-induced sesquiterpenoid synthesis in tobacco cells. Molecules 2012, 17, 1698–1715. [Google Scholar] [CrossRef]
- Aisa, H.A.; Xin, X.L.; Tang, D. Chemical constituents and their pharmacological activities of plants from Cichorium genus. Chin. Herb. Med. 2020, 12, 224–236. [Google Scholar] [CrossRef]
- Zhu, H.; Tu, P. A New Triterpenoid Saponin and Two Neolignans from Ligularia veitchiana. Z. Für Naturforschung B 2004, 59, 1063–1066. [Google Scholar] [CrossRef]
- Jia, X.; Yang, D.; Xie, H.; Jiang, Y.; Wei, X. Non-flavonoid phenolics from Averrhoa carambola fresh fruit. J. Funct. Foods 2017, 32, 419–425. [Google Scholar] [CrossRef]
- Alam, M.B.; Park, N.H.; Song, B.R.; Lee, S.H. Antioxidant Potential-Rich Betel Leaves (Piper betle L.) Exert Depigmenting Action by Triggering Autophagy and Downregulating MITF/Tyrosinase In Vitro and In Vivo. Antioxidants 2023, 12, 374. [Google Scholar] [CrossRef]
- Islam, S.; Alam, M.B.; Ahmed, A.; Lee, S.; Lee, S.-H.; Kim, S. Identification of secondary metabolites in Averrhoa carambola L. bark by high-resolution mass spectrometry and evaluation for α-glucosidase, tyrosinase, elastase, and antioxidant potential. Food Chem. 2020, 332, 127377. [Google Scholar] [CrossRef] [PubMed]
- Najm, O.A.; Addnan, F.H.; Mohd-Manzor, N.F.; Elkadi, M.A.; Abdullah, W.O.; Ismail, A.; Mansur, F.A.F. Identification of Phytochemicals of Phoenix dactylifera L. Cv Ajwa with UHPLC-ESI-QTOF-MS/MS. Int. J. Fruit Sci. 2021, 21, 848–867. [Google Scholar] [CrossRef]
- Hassanen, E.I.; Tohamy, A.F.; Issa, M.Y.; Ibrahim, M.A.; Farroh, K.Y.; Hassan, A.M. Pomegranate Juice Diminishes The Mitochondria-Dependent Cell Death And NF-kB Signaling Pathway Induced By Copper Oxide Nanoparticles On Liver And Kidneys Of Rats. Int. J. Nanomed. 2019, 14, 8905–8922. [Google Scholar] [CrossRef] [PubMed]
- Liu, W.Y.; Zou, C.M.; Hu, J.H.; Xu, Z.J.; Si, L.Q.; Liu, J.J.; Huang, J.G. Kinetic Characterization of Tyrosinase-catalyzed Oxidation of Four Polyphenols. Curr. Med. Sci. 2020, 40, 239–248. [Google Scholar] [CrossRef]
- Feng, D.; Fang, Z.; Zhang, P. The melanin inhibitory effect of plants and phytochemicals: A systematic review. Phytomedicine 2022, 107, 154449. [Google Scholar] [CrossRef] [PubMed]
- Chai, W.-M.; Huang, Q.; Lin, M.-Z.; Ou-Yang, C.; Huang, W.-Y.; Wang, Y.-X.; Xu, K.-L.; Feng, H.-L. Condensed Tannins from Longan Bark as Inhibitor of Tyrosinase: Structure, Activity, and Mechanism. J. Agric. Food Chem. 2018, 66, 908–917. [Google Scholar] [CrossRef] [PubMed]
- Monmai, C.; Kim, J.-S.; Chin, J.H.; Lee, S.; Baek, S.-H. Inhibitory Effects of Polyphenol- and Flavonoid-Enriched Rice Seed Extract on Melanogenesis in Melan-a Cells via MAPK Signaling-Mediated MITF Downregulation. Int. J. Mol. Sci. 2023, 24, 11841. [Google Scholar] [CrossRef] [PubMed]

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Run | Independent Variables | Response | |||||||
|---|---|---|---|---|---|---|---|---|---|
| EC (%) (X1) | Time (min) (X2) | Temp (°C) (X3) | TPC (mg GAE/g) (Y1) | TFC (mg CE/g) (Y2) | |||||
| RSM (prd.) | ANN (prd.) | Exp. | RSM (prd.) | ANN (prd.) | Exp. | ||||
| 1 | 80 | 15 | 50 | 64.99 | 65.59 | 65.15 ± 1.15 | 40.71 | 41.66 | 39.25 ± 1.05 |
| 2 | 80 | 30 | 40 | 62.30 | 62.60 | 61.56 ± 0.52 | 39.28 | 39.25 | 40.25 ± 0.98 |
| 3 | 60 | 45 | 60 | 64.13 | 64.89 | 63.55 ± 1.15 | 45.29 | 44.57 | 44.80 ± 0.56 |
| 4 | 60 | 30 | 50 | 75.70 | 76.88 | 75.26 ± 1.01 | 57.41 | 57.95 | 58.32 ± 0.28 |
| 5 | 60 | 30 | 50 | 75.77 | 76.85 | 75.56 ± 0.89 | 57.41 | 57.20 | 57.01 ± 1.15 |
| 6 | 60 | 30 | 50 | 75.87 | 75.32 | 76.15 ± 0.69 | 57.41 | 57.69 | 59.40 ± 0.89 |
| 7 | 80 | 45 | 50 | 61.57 | 61.60 | 61.95 ± 1.00 | 41.65 | 41.25 | 41.53 ± 0.79 |
| 8 | 60 | 15 | 60 | 65.62 | 66.29 | 64.55 ± 1.15 | 42.98 | 42.77 | 43.83 ± 0.69 |
| 9 | 60 | 45 | 40 | 62.97 | 62.57 | 63.35 ± 0.69 | 43.52 | 44.09 | 42.67 ± 1.09 |
| 10 | 40 | 15 | 50 | 61.21 | 61.55 | 60.92 ± 0.59 | 37.50 | 38.83 | 37.62 ± 1.10 |
| 11 | 60 | 15 | 40 | 65.80 | 66.33 | 66.39 ± 1.01 | 43.28 | 42.62 | 43.77 ± 0.99 |
| 12 | 40 | 30 | 60 | 60.34 | 60.95 | 61.09 ± 1.15 | 37.14 | 36.99 | 36.17 ± 1.09 |
| 13 | 40 | 45 | 50 | 60.38 | 61.75 | 60.23 ± 0.79 | 39.10 | 39.56 | 40.56 ± 1.00 |
| 14 | 60 | 30 | 50 | 75.17 | 76.60 | 76.65 ± 0.49 | 57.41 | 58.25 | 56.32 ± 1.02 |
| 15 | 60 | 30 | 50 | 75.77 | 76.05 | 75.25 ± 0.89 | 57.41 | 57.85 | 56.01 ± 0.58 |
| 16 | 40 | 30 | 40 | 61.10 | 61.59 | 60.89 ± 0.92 | 39.41 | 40.66 | 38.79 ± 0.65 |
| 17 | 80 | 30 | 60 | 64.03 | 65.60 | 64.15 ± 1.09 | 43.03 | 44.25 | 43.64 ± 0.45 |
| ANOVA for Quadratic Model for TPC | |||||||
|---|---|---|---|---|---|---|---|
| Source | RC | SS | DF | MS | F-Value | p-Value | |
| Model | 632.77 | 9 | 70.31 | 107.80 | <0.0001 | Significant | |
| Intercept | 75.77 | ||||||
| Linear terms | |||||||
| X1 | 1.21 | 11.71 | 1 | 11.71 | 17.96 | 0.0039 | Significant |
| X2 | −0.9912 | 7.86 | 1 | 7.86 | 12.05 | 0.0104 | Significant |
| X3 | 0.1438 | 0.1653 | 1 | 0.1653 | 0.2535 | 0.6301 | Not Significant |
| Interaction terms | |||||||
| X1X2 | −0.6275 | 1.58 | 1 | 1.58 | 2.41 | 0.1641 | Not significant |
| X1X3 | 0.5975 | 1.43 | 1 | 1.43 | 2.19 | 0.1825 | Not significant |
| X2X3 | 0.5100 | 1.04 | 1 | 1.04 | 1.60 | 0.2470 | Not Significant |
| Quadratic terms | |||||||
| X12 | −8.12 | 277.93 | 1 | 277.93 | 426.15 | <0.0001 | Significant |
| X22 | −5.59 | 131.43 | 1 | 131.43 | 201.52 | <0.0001 | Significant |
| X32 | −5.73 | 138.10 | 1 | 138.10 | 211.75 | <0.0001 | Significant |
| Lack of Fit | 3.07 | 3 | 1.02 | 2.74 | 0.1772 | Not significant | |
| Pure error | 1.49 | 4 | 0.3733 | ||||
| R2 | 0.9928 | ||||||
| Adjusted R2 | 0.9836 | ||||||
| Pred. R2 | 0.9192 | ||||||
| Adeq Precision | 25.0494 | ||||||
| C.V. % | 1.21 | ||||||
| ANOVA for quadratic model for TFC | |||||||
| Source | RC | SS | DF | MS | F-value | p-value | |
| Model | 1016.65 | 9 | 112.96 | 46.54 | <0.0001 | Significant | |
| Intercept | 57.42 | ||||||
| Linear terms | |||||||
| X1 | 1.44 | 16.62 | 1 | 16.62 | 6.85 | 0.0346 | Significant |
| X2 | 0.6375 | 3.25 | 1 | 3.25 | 1.34 | 0.2851 | Not Significant |
| X3 | 0.3691 | 1.09 | 1 | 1.09 | 0.4489 | 0.5243 | Not Significant |
| Interaction terms | |||||||
| X1X2 | −0.1651 | 0.1090 | 1 | 0.1090 | 0.0449 | 0.8382 | Not significant |
| X1X3 | 1.50 | 9.04 | 1 | 9.04 | 3.72 | 0.0949 | Not significant |
| X2X3 | 0.5177 | 1.07 | 1 | 1.07 | 0.4416 | 0.5276 | Not Significant |
| Quadratic terms | |||||||
| X12 | −10.86 | 496.93 | 1 | 496.93 | 204.73 | <0.0001 | Significant |
| X22 | −6.81 | 195.12 | 1 | 195.12 | 80.39 | <0.0001 | Significant |
| X32 | −6.83 | 196.58 | 1 | 196.58 | 80.99 | <0.0001 | Significant |
| Lack of Fit | 8.88 | 3 | 2.96 | 1.46 | 0.3516 | Not significant | |
| Pure error | 8.11 | 4 | 2.03 | ||||
| R2 | 0.9836 | ||||||
| Adjusted R2 | 0.9624 | ||||||
| Pred. R2 | 0.8503 | ||||||
| Adeq Precision | 16.9654 | ||||||
| C.V. % | 3.40 | ||||||
| Parameters | TPC | TFC | ||
|---|---|---|---|---|
| RSM | ANN | RSM | ANN | |
| R2 | 0.9928 | 0.9963 | 0.9836 | 0.9912 |
| RMSE | 6.7139 | 1.7760 | 3.8464 | 2.2384 |
| AAD (%) | 0.9237 | 0.1390 | 0.7828 | 0.2201 |
| SEP (%) | 0.0649 | 0.0171 | 0.0561 | 0.0326 |
| Groups | No. | Compound Name | EF | OM (m/z) | CM (m/z) | MS/MS | CL |
|---|---|---|---|---|---|---|---|
| Phenolic acids and derivatives | 1 | p-Coumaroyl aspartic acid | C13H13NO6 | 278.0669 | 278.0664 | 260.05, 234.07, 216.06 | 2 |
| 2 | 4-Hydroxybenzoyl glucose | C13H16O8 | 299.0773 | 299.0766 | 137.02, 163.02 | 1 | |
| 3 | Coumaroylshikimic acid | C16H16O7 | 319.0824 | 319.0817 | 173.04, 163.03, 145.02 | 2 | |
| 4 | Vanillic acid glucoside | C14H18O9 | 329.0879 | 329.0872 | 167.03, 152.02, 123.04 | 1 | |
| 5 | Caffeoyl shikimic acid | C16H16O8 | 335.0771 | 335.0772 | 179.01, 161.03, 155.03, 137.05 | 1 | |
| 6 | Glucosyringic acid | C15H20O10 | 359.0985 | 359.0978 | 197.04, 182.01, 153.05 | 2 | |
| 7 | Caffeic acid derivatives | C18H18O9 | 377.0853 | 377.0878 | 341.10, 215.03, 179.06, 161.04 | 2 | |
| 8 | Sinapic acid hexoside | C17H22O10 | 385.1154 | 385.1135 | 223.06, 205.05 | 1 | |
| 9 | Sinapoylspermine | C21H36N4O4 | 407.2649 | 407.2658 | 350.20, 279.13, 201.20 | 2 | |
| 10 | Methyl 4,6-di-O-galloyl-glucose | C21H22O14 | 497.0927 | 497.0931 | 345.05, 183.12, 169.05, 125.01 | 2 | |
| 11 | Caffeoyl shikimic acid hexoside | C22H26O13 | 497.1278 | 497.1295 | 335.01, 178.02, 135.02 | 2 | |
| 12 | Cinnamoyl-1,2-digalloyl glucose | C29H26O15 | 613.1126 | 613.1193 | 483.07, 443.09, 169.01, 147.04 | 2 | |
| 13 | 3-O-feruloyl-7-O-acyl-feruloyl-4-O-caffeoyl-quinic acid | C38H36O16 | 747.1895 | 747.1931 | 729.05, 687.15, 571.02, 529.05, 409.12, 381.05, 357.06 | 2 | |
| Flavonoids and derivatives | 14 | Apigenin | C15H10O5 | 269.0454 | 269.045 | 241.01, 151.01, 149.03 | 1 |
| 15 | Luteolin | C15H10O6 | 285.0405 | 285.0399 | 267.05, 241.03, 151.00, 133.02 | 1 | |
| 16 | Catechin/Epicatechin | C15H14O6 | 289.0718 | 289.0712 | 245.04, 205.05, 179, 151.04, 137.02 | 1 | |
| 17 | Chrysoeriol | C13H16O8 | 299.0561 | 299.0555 | 285.03, 255.02, 153.01, 135.03, 125.03 | 2 | |
| 18 | Quercetin | C15H10O7 | 301.0352 | 301.0348 | 273.04, 257.04, 179.00, 151.00 | 1 | |
| 19 | Taxifolin | C15H12O7 | 303.0511 | 303.0504 | 285.04, 275.02, 241.05, 151.04, 125.02 | 2 | |
| 20 | Epigallocatechin | C15H14O7 | 305.0637 | 305.0661 | 287.05, 137.02, 125.02 | 1 | |
| 21 | Methoxysinensetin | C21H22O8 | 401.1299 | 401.1236 | 371.11, 339.08, 191.71 | 2 | |
| 22 | Epicatechin hydroxybenzoate | C22H18O8 | 409.0924 | 409.0923 | 289.07, 271.06, 137.02, 119.01 | 2 | |
| 23 | Naringenin rhamnoside | C21H22O9 | 417.1249 | 417.1186 | 271.06, 187.03, 151.00, 119.05 | 2 | |
| 24 | Epiafzelechin gallate | C22H18O9 | 425.0877 | 425.0872 | 287.05, 273.07, 169.01, 151.00 | 2 | |
| 25 | Apigenin hexoside | C21H20O10 | 431.0989 | 431.0978 | 269.04, 241.01, 151.01, 149.03 | 1 | |
| 26 | Naringin | C21H22O10 | 433.1137 | 433.1134 | 271.06, 187.03, 151.00, 119.05 | 1 | |
| 27 | Epicatechin gallate | C22H18O10 | 441.0810 | 441.0821 | 135, 169, 273, 371, 399, 413, 427 | 2 | |
| 28 | Biochanin A glucoside | C22H22O10 | 445.1199 | 445.1135 | 283.06, 268.03, 239.03, 211.04, 132.02 | 2 | |
| 29 | Kaempferol hexoside | C21H20O11 | 447.0929 | 447.0927 | 285.04, 241.03, 151.00, 133.02 | 1 | |
| 30 | Taxifolin rhamnoside | C21H22O11 | 449.1089 | 449.1089 | 303.05, 285.04, 275.02, 151.04, 125.02 | 2 | |
| 31 | Catechin glucoside | C21H24O11 | 451.1356 | 451.1240 | 289.15, 151.10, 137.08, 123.10 | 2 | |
| 32 | Epicatechin 3-(3-methylgallate) | C23H20O10 | 455.1018 | 455.0978 | 289.02, 183.05, 124.01 | 2 | |
| 33 | Afrormosin glucoside | C23H24O10 | 459.1354 | 459.1291 | 297.07, 281.04, 267.06 | 2 | |
| 34 | Chrysoeriol hexoside | C22H22O11 | 461.1085 | 461.1083 | 299.05, 285.03, 153.01, 135.03, 125.03 | 2 | |
| 35 | Isoquercitrin | C21H20O12 | 463.0884 | 463.0876 | 301.05, 268.01, 179.02, 151.01 | 1 | |
| 36 | Epicatechin glucuronide | C21H22O12 | 465.1036 | 465.1033 | 289.15, 151.10, 137.08, 123.10 | 2 | |
| 37 | Epigallocatechin caffeate | C24H20O10 | 467.0980 | 467.0978 | 305.06, 287.05, 179.03, 137.02, 125.02 | 2 | |
| 38 | Isorhamnetin glucoside | C22H22O12 | 477.1035 | 477.1033 | 315.05, 300.01, 255.05, 179.05, 151.02 | 2 | |
| 39 | Luteone glucoside | C26H28O11 | 515.1615 | 515.1553 | 353.10, 311.05, 297.04 | 2 | |
| 40 | Isorhamnetin malonyl hexoside | C24H24O13 | 519.1141 | 519.1138 | 315.05, 300.02, 227.01, 204.04, 177.01 | 2 | |
| 41 | Luteolin hexosyl sulfate | C21H20O14S | 527.0502 | 527.0495 | 447.05, 285.01, 241.06 | 2 | |
| 42 | Chrysoeriol hexosyl sulfate | C22H22O14S | 541.0658 | 541.0652 | 299.05, 284.05, 241.02 | 2 | |
| 43 | Isoquercitrin sulfate | C21H20O15S | 543.0448 | 543.0444 | 463.05, 301.01, 268.01, 179.02, 151.01 | 2 | |
| 44 | Procyanidin A2 | C30H24O12 | 575.1195 | 575.1189 | 539.09, 449.08, 423.07, 289.07, 285.04, 269.04, 125.02 | 1 | |
| 45 | Procyanidin B2 | C30H26O12 | 577.1352 | 577.1346 | 451.10, 425.08, 407.07, 289.07, 287.05, 269.04, 125.02 | 1 | |
| 46 | Luteolin rhamnosyl hexoside | C27H30O15 | 593.1509 | 593.1506 | 447.09, 285.03, 153.01, 135.04 | 2 | |
| 47 | Chrysoeriol rhamnosyl hexoside | C28H32O15 | 607.1672 | 607.1663 | 461.10, 299.05, 284.03, 153.01, 149.05 | 2 | |
| 48 | Isorhamnetin rhamnosyl hexoside | C28H32O16 | 623.1609 | 623.1612 | 477.10, 315.05, 299.05, 165.05 | 2 | |
| 49 | Isorhamnetin dihexoside | C28H32O17 | 639.1556 | 639.1561 | 447.01, 315.01 | 2 | |
| 50 | Procyanidin B2 gallate | C37H30O16 | 729.1473 | 729.1455 | 451.10, 425.08, 407.07, 289.07, 287.05, 169.01, 125.02 | 2 | |
| 51 | Kaempferol 3-(3″,6″-di-p-coumaroyl galactoside) (Stenopalustroside A) | C39H32O15 | 739.1679 | 739.1663 | 593.12, 575.11, 285.03, 163.03 | 2 | |
| 52 | Quercetin 3-O-xylosyl-rutinoside | C32H38O20 | 741.1846 | 741.1878 | 609.14, 301.03 | 2 | |
| 53 | Epicatechin-(4beta- >8)-epigallocatechin gallate | C37H30O17 | 745.1395 | 745.1404 | 441.08, 303.05, 169.01, 125.02 | 2 | |
| 54 | Luteolin rhamnosyl dihexoside | C33H40O20 | 755.1990 | 755.2034 | 709.16, 593.10, 575.05, 285.01 | 2 | |
| 55 | Quercetin rhamnosyl dihexoside | C33H40O21 | 771.1969 | 771.1983 | 609.14, 591.05, 301.03, 153.02, 125.00 | 2 | |
| 56 | Isorhamnetin rhamnosyl dihexoside | C34H42O21 | 785.2110 | 785.2140 | 623.16, 477.10, 315.05 | 2 | |
| 57 | Quercetin 3-sophorotrioside | C33H40O22 | 787.1909 | 787.1933 | 625.10, 463.09, 301.01 | 2 | |
| 58 | Quercetin 3-(6′′′′-p-coumaryl sophorotrioside) (Pisumflavonoside I) | C42H46O24 | 933.2302 | 933.2300 | 787.19, 625.10, 463.09, 301.01 | 2 | |
| 59 | Quercetin 3-(6″-caffeoyl sophorotrioside) | C42H46O25 | 949.2223 | 949.2250 | 787.19, 625.10, 463.09, 301.01 | 2 | |
| Terpenoids | 60 | 8-Hydroxy-(+)-δ-cadinene | C15H24O | 219.175 | 219.1748 | 203.14, 201.16, 179.14 | 2 |
| 61 | Valerenic acid | C15H22O2 | 233.1544 | 233.1541 | 219.13, 189.16, 161.13 | 2 | |
| 62 | β-Ionyl acetate | C15H24O2 | 235.1700 | 235.1698 | 193.15, 175.14, 149.13 | 2 | |
| 63 | Valerenolic acid | C15H22O3 | 249.1528 | 249.1490 | 231.13, 205.15, 187.14, 177.12 | 2 | |
| 64 | Phytuberol | C15H24O3 | 251.1651 | 251.1647 | 233.15, 221.15, 193.12 | 2 | |
| 65 | Curcolonol | C15H20O4 | 263.1288 | 263.1283 | 245.11, 227.10, 205.08 | 2 | |
| 66 | Absindiol | C15H22O4 | 265.1445 | 265.1439 | 247.13, 221.15, 209.11 | 2 | |
| 67 | Acoric acid | C15H24O4 | 267.1602 | 267.1596 | 249.14, 223.16, 181.12 | 2 | |
| 68 | Phytuberin | C17H26O4 | 293.1758 | 293.1752 | 251.16, 233.15, 221.15, 193.12 | 2 | |
| 69 | Trilobinol | C20H28O2 | 299.2016 | 299.2011 | 283.16, 265.15, 257.15 | 2 | |
| 70 | Abietadiene-diol | C20H32O2 | 303.2330 | 303.2324 | 287.20, 257.22, 241.19, 215.18 | 2 | |
| 71 | Piperochromenoic acid | C22H28O3 | 339.2000 | 339.1960 | 325.18, 295.20, 189.05, 137.02 | 2 | |
| 72 | Eucannabinolide | C22H28O8 | 419.1710 | 419.1705 | 389.16, 371.14, 359.14, 347.14 | 2 | |
| 73 | β-Amyrenone | C30H48O | 423.3624 | 423.3626 | 407.33, 391.30 | 2 | |
| 74 | Cichorioside M | C21H32O9 | 427.1974 | 427.1968 | 265.14, 247.13, 221.15, 209.11 | 2 | |
| 75 | Cynaroside A | C21H32O10 | 443.1921 | 443.1917 | 281.13, 263.12, 237.14, 193.12 | 2 | |
| 76 | Oleanonic acid | C30H46O3 | 453.3376 | 453.3368 | 241, 323, 341, 379 | 2 | |
| Lignans | 77 | 1,2-Di-(syringoyl)-hexoside | C24H28O14 | 539.1385 | 539.1401 | 359.09, 341.08, 197.04, 153.05 | 2 |
| 78 | Citrusin B | C27H36O13 | 567.2084 | 567.2077 | 405.15, 387.14, 358.14, 209.08, 197.08 | 3 | |
| 79 | Lyoniresinol glucoside | C28H37O13 | 581.2236 | 581.2234 | 419.17, 265.10, 247.09 | 3 | |
| Carboxylic acid, fatty acids and amino acids | 80 | Fumaric acid | C4H4O4 | 115.0026 | 115.0037 | 71.01 | 2 |
| 81 | Succinic acid | C4H6O4 | 117.0183 | 117.0187 | 99.00, 73.02 | 2 | |
| 82 | Malic acid | C4H6O5 | 133.0133 | 133.0142 | 115.00, 89.02, 71.01 | 2 | |
| 83 | Tartaric acid | C4H6O6 | 148.9235 | 149.0086 | 87.05 | 2 | |
| 84 | Ribonic acid | C5H10O6 | 165.0398 | 165.0418 | 149.04, 105.01, 87.00, 75.00 | 2 | |
| 85 | Citric acid | C6H8O7 | 191.0191 | 191.0197 | 173.00, 129.01, 111.00 | 2 | |
| 86 | Homocitric acid | C7H10O7 | 205.0349 | 205.0348 | 161.04. 143.04. 117.05 | 2 | |
| 87 | Lauric acid | C12H24O2 | 199.1698 | 199.1698 | 181.16, 165.13, 163.11, 139.11, 135.11 | 2 | |
| 88 | Myristic acid | C14H28O2 | 227.2014 | 227.2011 | 209.19, 183.21, 179.18 | 2 | |
| 89 | Methylmyristic acid | C15H30O2 | 241.2171 | 241.2167 | 227.20, 209.19, 183.21, 179.18 | 2 | |
| 90 | Palmitic acid | C16H32O2 | 255.2327 | 255.233 | 237.23, 211.24, 197.22 | 2 | |
| 91 | 16-Hydroxypalmitic acid | C16H32O3 | 271.2279 | 271.2273 | 253.12, 237.22. 225.25, 211.24. 195.21 | 2 | |
| 92 | α-Linoleic acid | C18H32O2 | 279.2328 | 279.233 | 261.22 | 2 | |
| 93 | Oleic acid | C18H34O2 | 281.2485 | 281.2486 | 263.25, 181.21, 127.25 | 2 | |
| 94 | Dihydroxy octadecadienoic acid | C18H32O4 | 311.2226 | 311.2239 | 293.22, 275.23 | 2 | |
| 95 | Dihydroxy octadecenoic acid | C18H34O4 | 313.2383 | 313.2378 | 295.23, 277.25, 183.32 | 2 | |
| 96 | Dihydroxy octadecanoic acid | C18H36O4 | 315.2538 | 315.2535 | 297.23, 279.25, | 2 | |
| 97 | Trihydroxy octadecadienoic acid | C18H32O5 | 327.2175 | 327.2171 | 309.23, 291.25, 273.23 | 2 | |
| 98 | Trihydroxy octadecenoic acid | C18H34O5 | 329.2332 | 329.2333 | 311.25, 293.26, 275.23 | 2 | |
| 99 | α-Hydroxybehenic acid | C22H44O3 | 355.3217 | 355.3212 | 337.31, 311.33, 293.32, 281.32 | 2 | |
| 100 | 26-Hydroxyhexacosanoic acid | C26H52O3 | 411.3842 | 411.3838 | 393.37, 381.37, 367.39 | 2 | |
| Others | 101 | Dihydrojasmonic acid | C12H20O3 | 211.1335 | 211.1334 | 167.14, 111.08, 59.10 | 2 |
| 102 | N-acetyl-α-neuraminic acid | C11H19NO9 | 308.0986 | 308.0987 | 290.09, 219.06, 200.05, 146.08, 128.07 | 2 | |
| 103 | 1-Deoxynojirimycin hexoside | C12H23NO9 | 324.1298 | 324.1295 | 161.04, 144.06, 143.03, 113.02 | 2 | |
| 104 | Icariside D1 | C19H28O10 | 415.1609 | 415.1604 | 398.15, 384.14, 250.12 | 2 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Alshammari, F.; Alam, M.B.; Naznin, M.; Kim, S.; Lee, S.-H. Optimization, Metabolomic Analysis, Antioxidant Potential and Depigmenting Activity of Polyphenolic Compounds from Unmature Ajwa Date Seeds (Phoenix dactylifera L.) Using Ultrasonic-Assisted Extraction. Antioxidants 2024, 13, 238. https://doi.org/10.3390/antiox13020238
Alshammari F, Alam MB, Naznin M, Kim S, Lee S-H. Optimization, Metabolomic Analysis, Antioxidant Potential and Depigmenting Activity of Polyphenolic Compounds from Unmature Ajwa Date Seeds (Phoenix dactylifera L.) Using Ultrasonic-Assisted Extraction. Antioxidants. 2024; 13(2):238. https://doi.org/10.3390/antiox13020238
Chicago/Turabian StyleAlshammari, Fanar, Md Badrul Alam, Marufa Naznin, Sunghwan Kim, and Sang-Han Lee. 2024. "Optimization, Metabolomic Analysis, Antioxidant Potential and Depigmenting Activity of Polyphenolic Compounds from Unmature Ajwa Date Seeds (Phoenix dactylifera L.) Using Ultrasonic-Assisted Extraction" Antioxidants 13, no. 2: 238. https://doi.org/10.3390/antiox13020238
APA StyleAlshammari, F., Alam, M. B., Naznin, M., Kim, S., & Lee, S.-H. (2024). Optimization, Metabolomic Analysis, Antioxidant Potential and Depigmenting Activity of Polyphenolic Compounds from Unmature Ajwa Date Seeds (Phoenix dactylifera L.) Using Ultrasonic-Assisted Extraction. Antioxidants, 13(2), 238. https://doi.org/10.3390/antiox13020238