

The Amazonian Camu-Camu Fruit Modulates the Development of Drosophila melanogaster and the Neural Function of Adult Flies under Oxidative Stress Conditions

 , , and
, , and
Abstract

1. Introduction
2. Materials and Methods
2.1. Camu-Camu Powder
2.2. D. melanogaster Stock
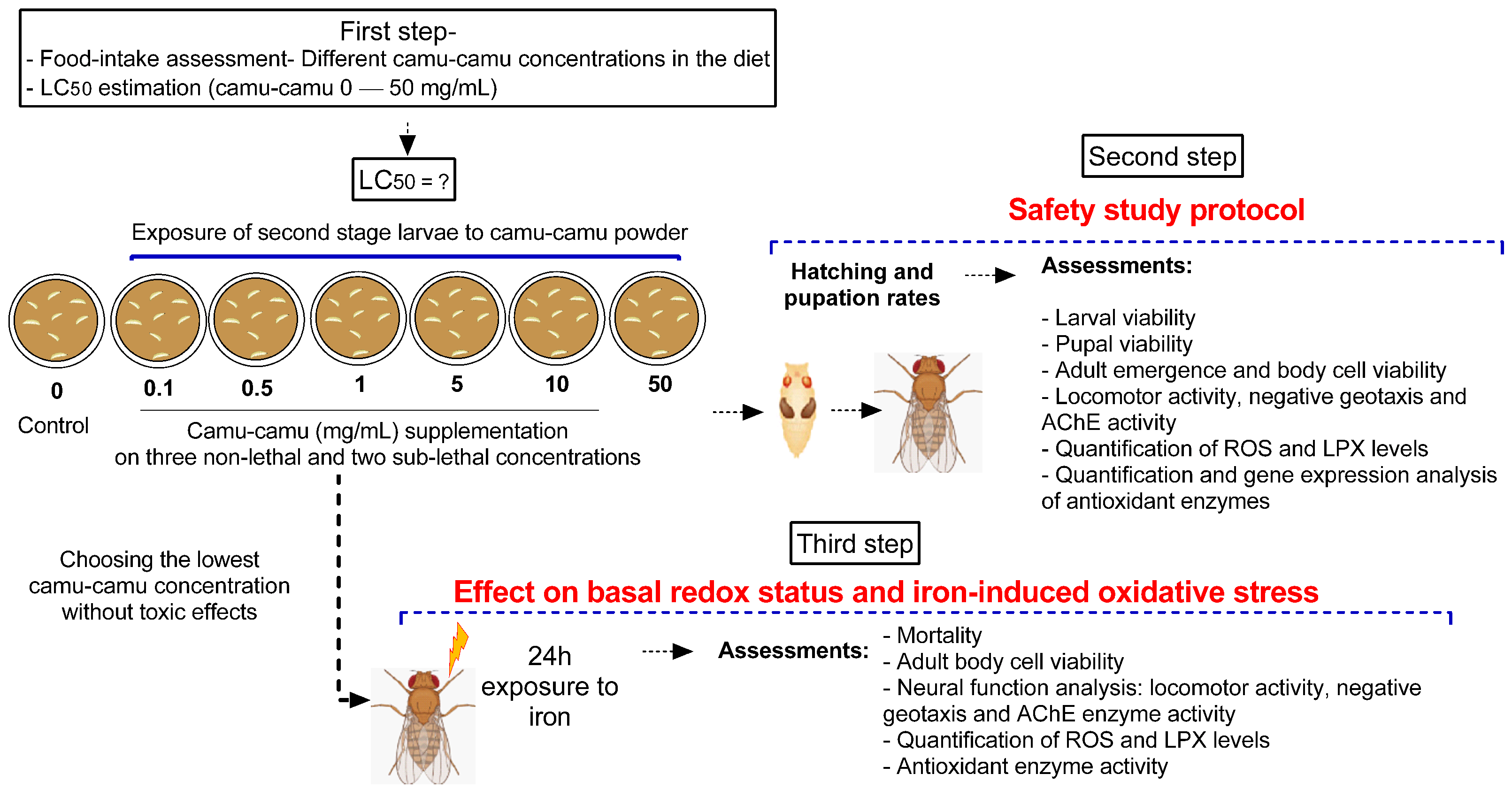
2.3. Experimental Design
2.4. Step One: Camu-Camu Powder Safety Assessment
2.4.1. Food Intake
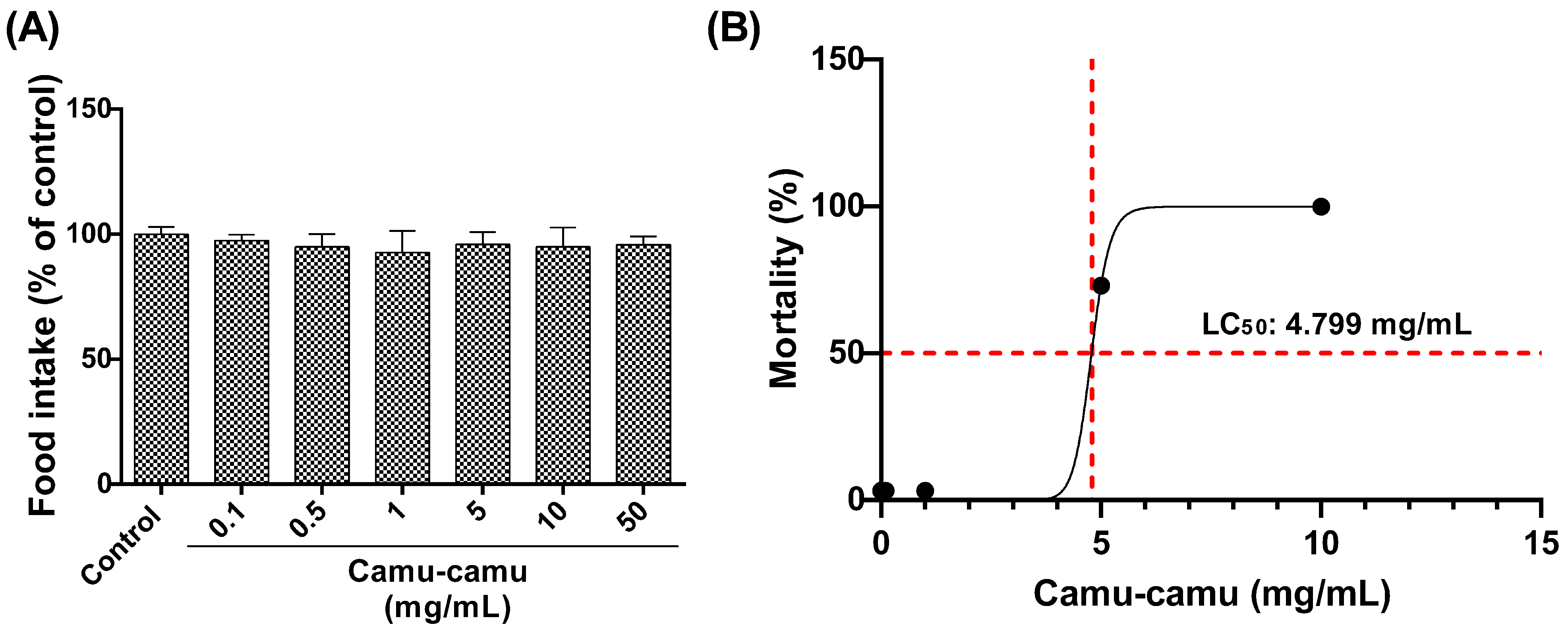
2.4.2. Lethal Concentrations
2.4.3. Determination of Safe Concentrations of Camu-Camu Powder
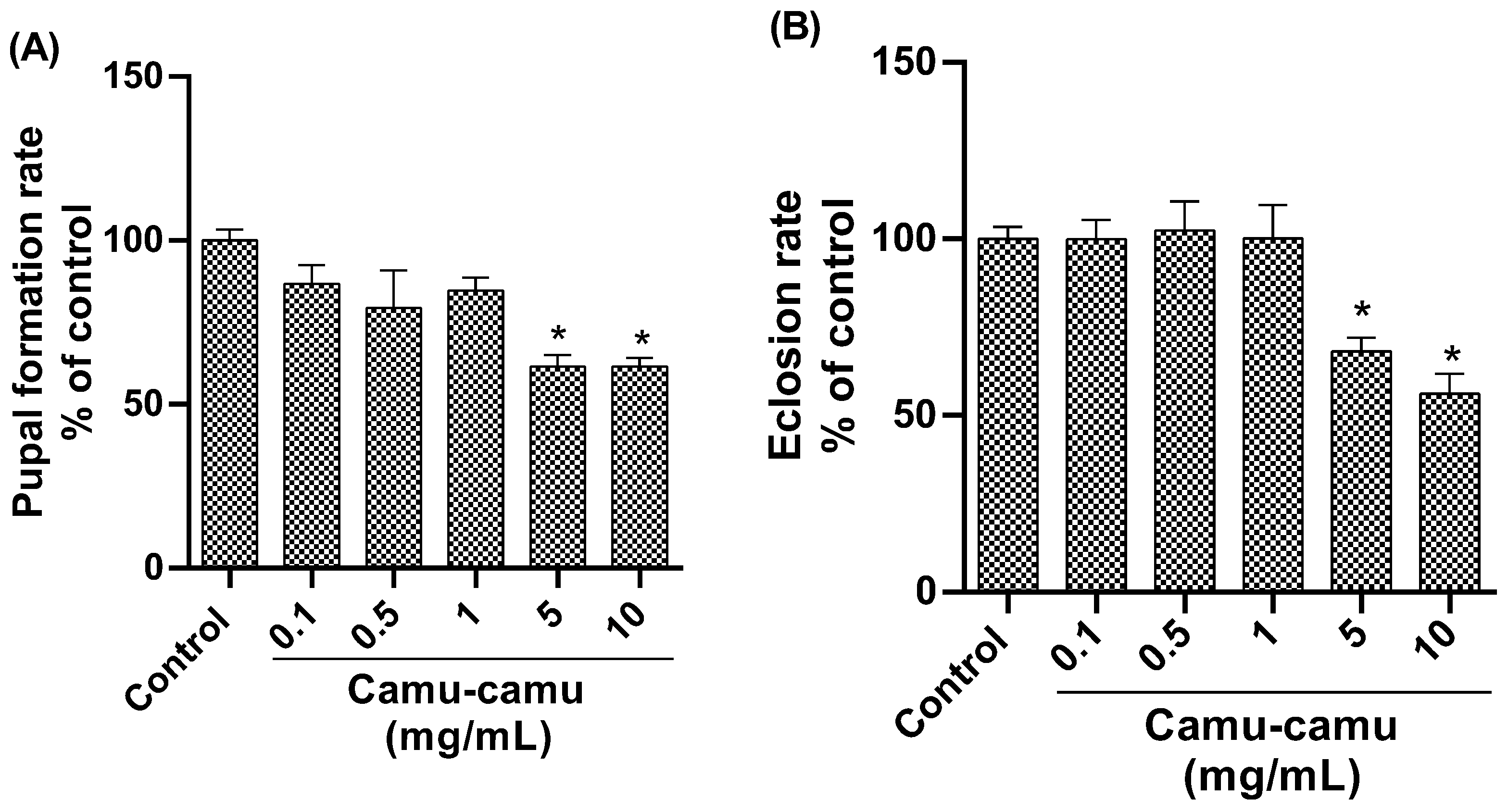
2.4.4. Pupation and Hatching Rate
2.5. Iron-Induced Oxidative Stress Model
2.6. Adult Neural Function Assays
2.6.1. Locomotor Activity Behavior during an Open-Field Test
2.6.2. Negative Geotaxis Behavior Assay
2.7. Adult D. melanogaster Biochemical and Molecular Assays
2.7.1. D. melanogaster (Body) Cell Viability Assay
2.7.2. Acetylcholinesterase Activity Assay
2.7.3. Reactive Species Quantification
2.7.4. Lipid Peroxidation Quantification
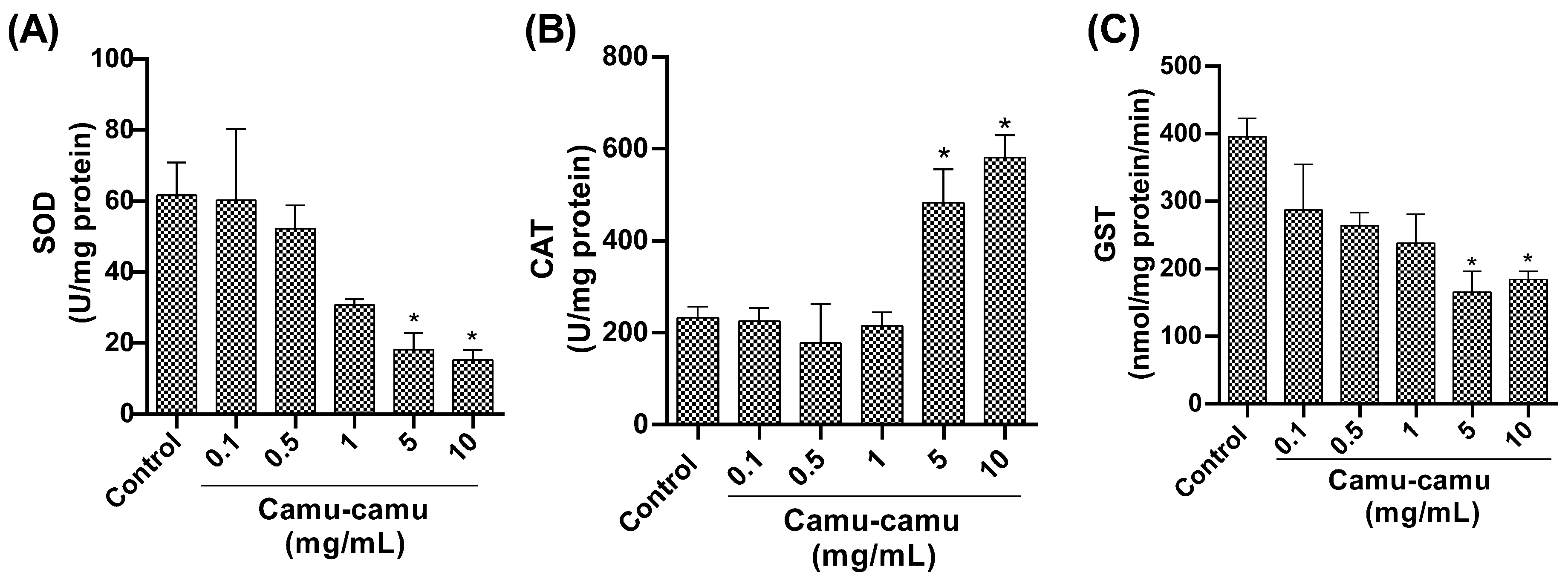
2.7.5. Superoxide Dismutase Activity Quantification
2.7.6. Catalase Activity Quantification
2.7.7. Glutathione S-Transferase Activity Quantification
2.7.8. Total Protein Quantification
2.8. Gene Expression by qRT-PCR Molecular Assay
2.9. Statistical Analysis
3. Results
3.1. Safety Evaluation of Different Concentrations of Camu-Camu Powder
3.1.1. Food Intake Test and LC50
3.1.2. Hatching and Pupation Rate
3.1.3. Behavioral Tests and AChE Activity
3.1.4. Quantification of Reactive Species, Lipid Peroxidation Levels, and Cell Viability
3.1.5. Antioxidant and Detoxification Enzyme Activity
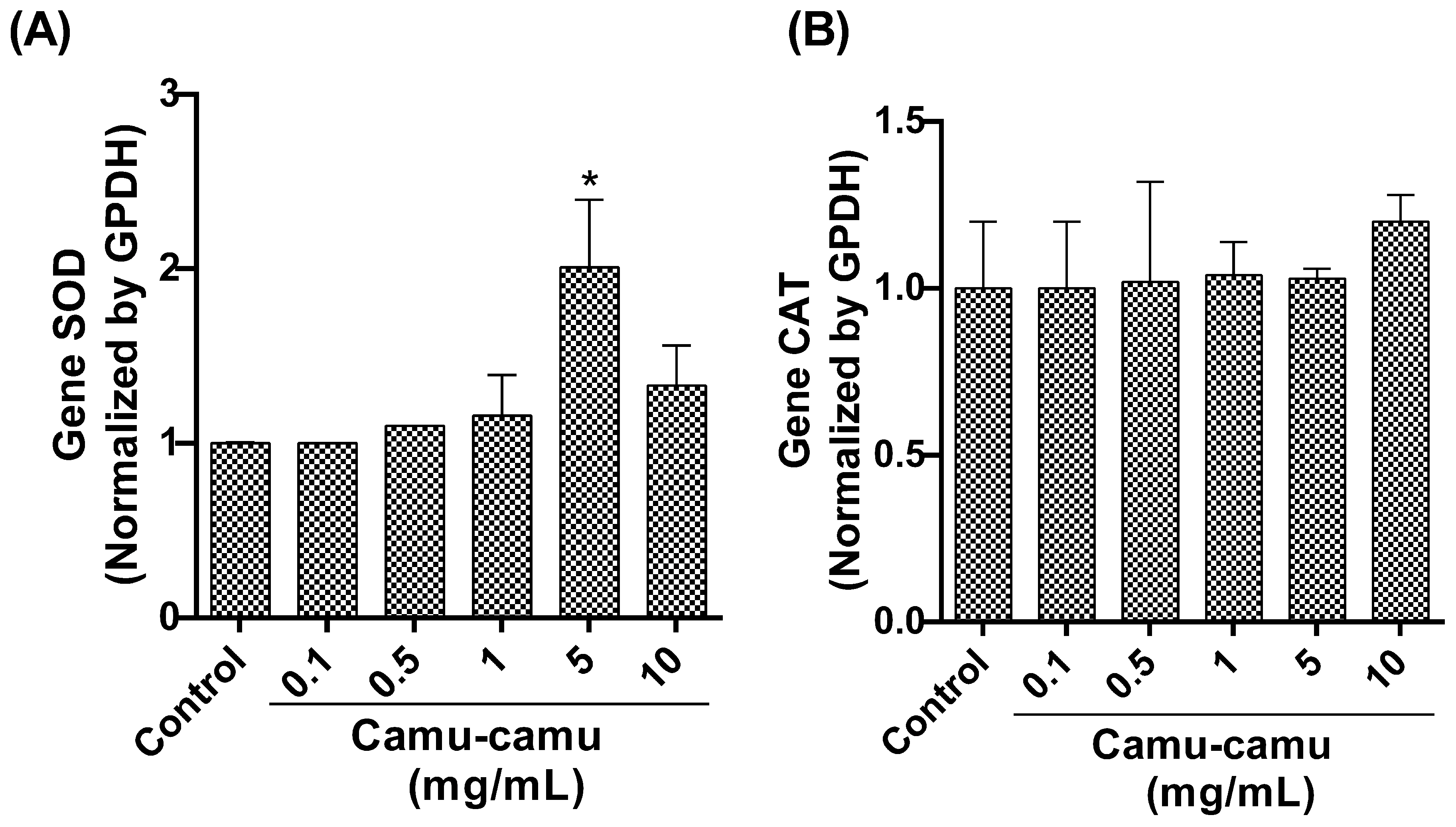
3.1.6. SOD and CAT Expression
3.2. The Protective Effect of Camu-Camu Powder against Fe-Induced Oxidative Stress
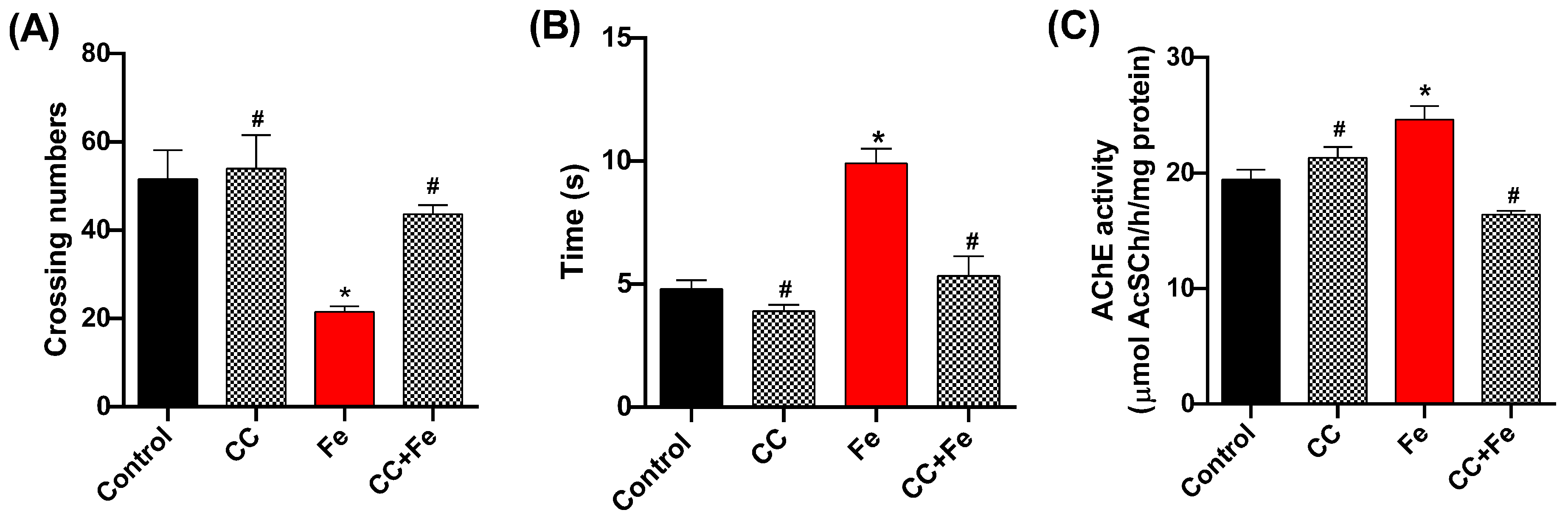
3.2.1. Locomotor and Climbing Capacity and AChE Activity in Relation to Fe Exposure
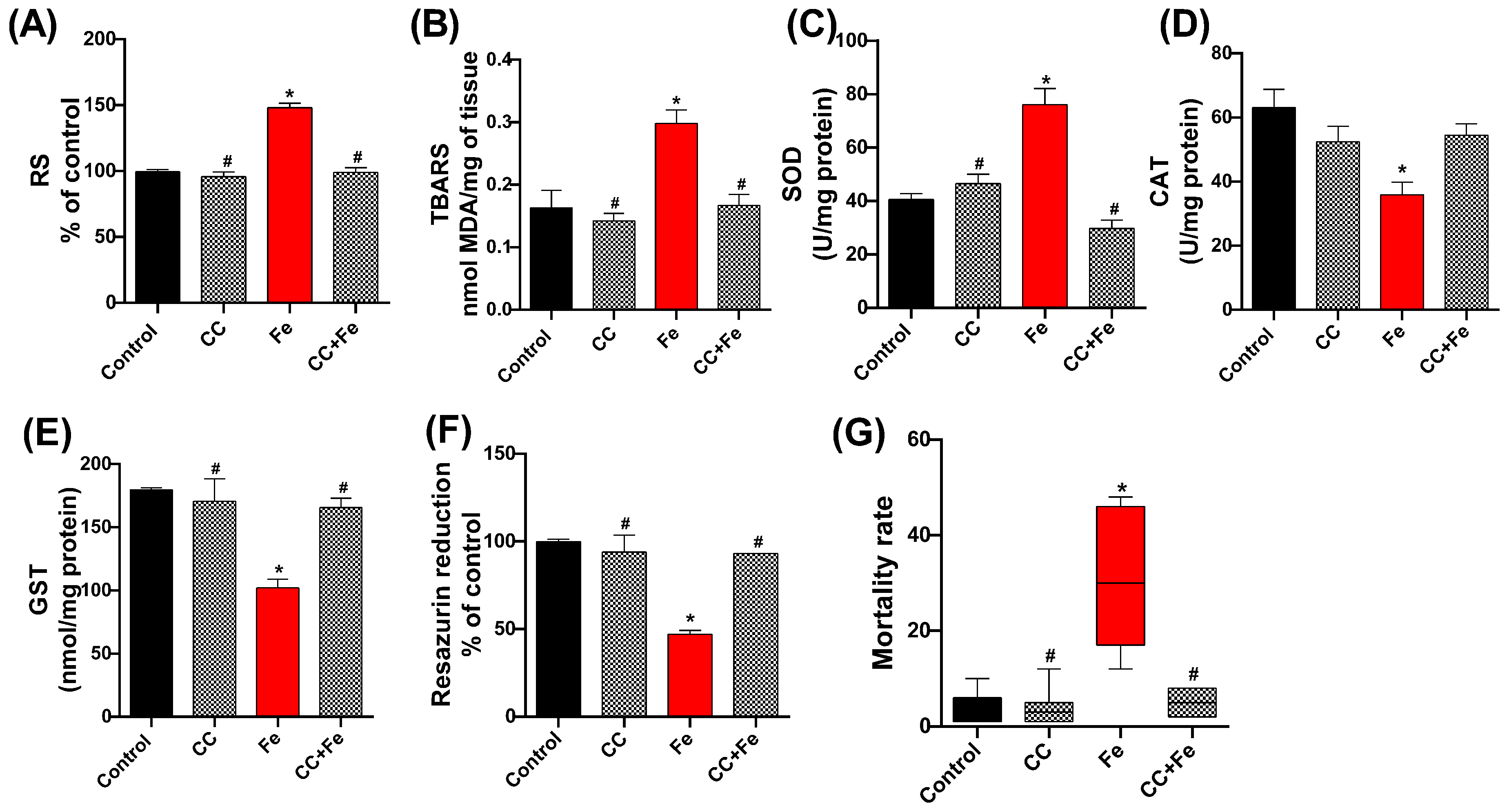
3.2.2. Oxidative Stress Markers, Cell Viability, and Mortality of Flies in Relation to Fe Exposure
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Serafini, M.; Peluso, I. Functional Foods for Health: The Interrelated Antioxidant and Anti-Inflammatory Role of Fruits, Vegetables, Herbs, Spices and Cocoa in Humans. Curr. Pharm. Des. 2016, 22, 6701–6715. [Google Scholar] [CrossRef] [PubMed]
- Santos, O.V.D.; Pinaffi Langley, A.C.d.C.; Mota de Lima, A.J.; Vale Moraes, V.S.; Dias Soares, S.; Teixeira-Costa, B.E. Nutraceutical potential of Amazonian oilseeds in modulating the immune system against COVID-19—A narrative review. J. Funct. Foods 2022, 94, 105123. [Google Scholar] [CrossRef]
- Lima, R.S.; de Carvalho, A.P.A.; Conte-Junior, C.A. Health from Brazilian Amazon food wastes: Bioactive compounds, antioxidants, antimicrobials, and potentials against cancer and oral diseases. Crit. Rev. Food Sci. Nutr. 2022, 63, 12453–12475. [Google Scholar] [CrossRef]
- de Carvalho, A.P.A.; Conte-Junior, C.A. Health benefits of phytochemicals from Brazilian native foods and plants: Antioxidant, antimicrobial, anti-cancer, and risk factors of metabolic/endocrine disorders control. Trends Food Sci. Technol. 2021, 111, 534–548. [Google Scholar] [CrossRef]
- Neves, L.C.; Campos, A.J.d.; Benedette, R.M.; Tosin, J.M.; Chagas, E.A. Characterization of the antioxidant capacity of natives fruits from the Brazilian Amazon Region. Rev. Bras. Frutic. 2012, 34, 1165–1173. [Google Scholar] [CrossRef]
- García-Chacón, J.M.; Marín-Loaiza, J.C.; Osorio, C. Camu Camu (Myrciaria dubia (Kunth) McVaugh): An Amazonian Fruit with Biofunctional Properties—A Review. ACS Omega 2023, 8, 5169–5183. [Google Scholar] [CrossRef] [PubMed]
- Fracassetti, D.; Costa, C.; Moulay, L.; Tomás-Barberán, F.A. Ellagic acid derivatives, ellagitannins, proanthocyanidins and other phenolics, vitamin C and antioxidant capacity of two powder products from camu-camu fruit (Myrciaria dubia). Food Chem. 2013, 139, 578–588. [Google Scholar] [CrossRef]
- Yi, Y.; Xu, W.; Fan, Y.; Wang, H.X. Drosophila as an emerging model organism for studies of food-derived antioxidants. Food Res. Int. 2021, 143, 110307. [Google Scholar] [CrossRef]
- Kawabata, T. Iron-Induced Oxidative Stress in Human Diseases. Cells 2022, 11, 2152. [Google Scholar] [CrossRef]
- Lushchak, O.V.; Rovenko, B.M.; Gospodaryov, D.V.; Lushchak, V.I. Drosophila melanogaster larvae fed by glucose and fructose demonstrate difference in oxidative stress markers and antioxidant enzymes of adult flies. Comp. Biochem. Physiol. A Mol. Integr. Physiol. 2011, 160, 27–34. [Google Scholar] [CrossRef]
- Mishra, M.; Panda, M. Reactive oxygen species: The root cause of nanoparticle-induced toxicity in Drosophila melanogaster. Free Radic. Res. 2021, 55, 671–687. [Google Scholar] [CrossRef] [PubMed]
- Musachio, E.A.S.; de Freitas Couto, S.; Poetini, M.R.; Bortolotto, V.C.; Dahleh, M.M.M.; Janner, D.E.; Araujo, S.M.; Ramborger, B.P.; Rohers, R.; Guerra, G.P.; et al. Bisphenol A exposure during the embryonic period: Insights into dopamine relationship and behavioral disorders in Drosophila melanogaster. Food Chem. Toxicol. 2021, 157, 112526. [Google Scholar] [CrossRef] [PubMed]
- Poetini, M.R.; Araujo, S.M.; Trindade de Paula, M.; Bortolotto, V.C.; Meichtry, L.B.; Polet de Almeida, F.; Jesse, C.R.; Kunz, S.N.; Prigol, M. Hesperidin attenuates iron-induced oxidative damage and dopamine depletion in Drosophila melanogaster model of Parkinson’s disease. Chem. Biol. Interact. 2018, 279, 177–186. [Google Scholar] [CrossRef] [PubMed]
- Connolly, K. Locomotor activity in drosophila. II. Selection for active and inactive strains. Anim. Behav. 1966, 14, 444–449. [Google Scholar] [CrossRef] [PubMed]
- Musachio, E.A.S.; Araujo, S.M.; Bortolotto, V.C.; de Freitas Couto, S.; Dahleh, M.M.M.; Poetini, M.R.; Jardim, E.F.; Meichtry, L.B.; Ramborger, B.P.; Roehrs, R.; et al. Bisphenol A exposure is involved in the development of Parkinson like disease in Drosophila melanogaster. Food Chem. Toxicol. 2020, 137, 111128. [Google Scholar] [CrossRef] [PubMed]
- Jimenez-Del-Rio, M.; Guzman-Martinez, C.; Velez-Pardo, C. The effects of polyphenols on survival and locomotor activity in Drosophila melanogaster exposed to iron and paraquat. Neurochem. Res. 2010, 35, 227–238. [Google Scholar] [CrossRef] [PubMed]
- Franco, J.L.; Posser, T.; Dunkley, P.R.; Dickson, P.W.; Mattos, J.J.; Martins, R.; Bainy, A.C.D.; Marques, M.R.; Dafre, A.L.; Farina, M. Methylmercury neurotoxicity is associated with inhibition of the antioxidant enzyme glutathione peroxidase. Free Radic. Biol. Med. 2009, 47, 449–457. [Google Scholar] [CrossRef]
- Gamage, R.; Wagnon, I.; Rossetti, I.; Childs, R.; Niedermayer, G.; Chesworth, R.; Gyengesi, E. Cholinergic modulation of glial function during aging and chronic neuroinflammation. Front. Cell. Neurosci. 2020, 14, 577912. [Google Scholar] [CrossRef]
- Adedara, A.O.; Otenaike, T.A.; Olabiyi, A.A.; Adedara, I.A.; Abolaji, A.O. Neurotoxic and behavioral deficit in Drosophila melanogaster co-exposed to rotenone and iron. Metab. Brain Dis. 2023, 38, 349–360. [Google Scholar] [CrossRef]
- Ellman, G.L.; Courtney, K.D.; Andres, V., Jr.; Feather-Stone, R.M. A new and rapid colorimetric determination of acetylcholinesterase activity. Biochem. Pharmacol. 1961, 7, 88–95. [Google Scholar] [CrossRef]
- Pérez-Severiano, F.; Santamaría, A.; Pedraza-Chaverri, J.; Medina-Campos, O.N.; Ríos, C.; Segovia, J. Increased formation of reactive oxygen species, but no changes in glutathione peroxidase activity, in striata of mice transgenic for the Huntington’s disease mutation. Neurochem. Res. 2004, 29, 729–733. [Google Scholar] [CrossRef] [PubMed]
- Ohkawa, H.; Ohishi, N.; Yagi, K. Assay for lipid peroxides in animal tissues by thiobarbituric acid reaction. Anal. Biochem. 1979, 95, 351–358. [Google Scholar] [CrossRef] [PubMed]
- Pinheiro, F.C.; Bortolotto, V.C.; Araujo, S.M.; Poetini, M.R.; Sehn, C.P.; Neto, J.S.S.; Zeni, G.; Prigol, M. Antimicrobial effect of 2-phenylethynyl-butyltellurium in Escherichia coli and its association with oxidative stress. J. Microbiol. Biotechnol. 2018, 28, 1209–1216. [Google Scholar] [CrossRef] [PubMed]
- Aebi, H. Catalase in vitro. In Methods in Enzymology; Elsevier: Amsterdam, The Netherlands, 1984; pp. 121–126. [Google Scholar]
- Santos Musachio, E.A.; Poetini, M.R.; Janner, D.E.; Fernandes, E.J.; Meichtry, L.B.; Mustafa Dahleh, M.M.; Guerra, G.P.; Prigol, M. Safer alternatives? Bisphenol F and Bisphenol S induce oxidative stress in Drosophila melanogaster larvae and trigger developmental damage. Food Chem. Toxicol. 2023, 175, 113701. [Google Scholar] [CrossRef]
- Bradford, M.M. A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein-dye binding. Anal. Biochem. 1976, 72, 248–254. [Google Scholar] [CrossRef] [PubMed]
- da Cruz, I.B.M.; de Afonso Bonotto, N.C.; Turra, B.O.; Teixeira, C.F.; Azzolin, V.F.; Ribeiro, E.A.M.; Piccoli, J.D.C.E.; Barbisan, F. Rotenone-exposure as cytofunctional aging model of human dermal fibroblast prior replicative senescence. Toxicol. In Vitro 2023, 91, 105637. [Google Scholar] [CrossRef] [PubMed]
- Jaszczak, J.S.; Halme, A. Arrested development: Coordinating regeneration with development and growth in Drosophila melanogaster. Curr. Opin. Genet. Dev. 2016, 40, 87–94. [Google Scholar] [CrossRef] [PubMed]
- Dhage, P.A.; Sharbidre, A.A. Bimodal Behavior of Ascorbic Acid in Musca domestica Larvae. Biointerface Res. Appl. Chem. 2021, 12, 5199–5216. [Google Scholar]
- de Freitas Couto, S.; Araujo, S.M.; Bortolotto, V.C.; Poetini, M.R.; Pinheiro, F.C.; Santos Musachio, E.A.; Meichtry, L.B.; do Sacramento, M.; Alves, D.; La Rosa Novo, D.; et al. 7-chloro-4-(phenylselanyl) quinoline prevents dopamine depletion in a Drosophila melanogaster model of Parkinson’s-like disease. J. Trace Elem. Med. Biol. 2019, 54, 232–243. [Google Scholar] [CrossRef]
- Pandey, U.B.; Nichols, C.D. Human disease models in Drosophila melanogaster and the role of the fly in therapeutic drug discovery. Pharmacol. Rev. 2011, 63, 411–436. [Google Scholar] [CrossRef]
- Jordan, A.C.; Perry, C.G.R.; Cheng, A.J. Promoting a pro-oxidant state in skeletal muscle: Potential dietary, environmental, and exercise interventions for enhancing endurance-training adaptations. Free Radic. Biol. Med. 2021, 176, 189–202. [Google Scholar] [CrossRef] [PubMed]
- Kim, J.H.; Kang, J.C. Detoxification effects of ascorbic acid on the oxidative stress, neurotoxicity, and metallothionein (MT) gene expression in juvenile rockfish, Sebastes schlegelii by the dietary chromium exposure. Fish Shellfish Immunol. 2023, 132, 108464. [Google Scholar] [CrossRef] [PubMed]
- Aversa, R.; Petrescu, R.V.V.; Apicella, A.; Petrescu, F.I.T. One Can Slow Down the Aging through Antioxidants. Am. J. Eng. Appl. Sci. 2016, 9, 1112–1126. [Google Scholar] [CrossRef]
- Scandalios, J. Oxidative stress: Molecular perception and transduction of signals triggering antioxidant gene defenses. Braz. J. Med. Biol. Res. 2005, 38, 995–1014. [Google Scholar] [CrossRef] [PubMed]
- Ou, M.; Jiang, Y.; Ji, Y.; Zhou, Q.; Du, Z.; Zhu, H.; Zhou, Z. Role and mechanism of ferroptosis in neurological diseases. Mol. Metab. 2022, 61, 101502. [Google Scholar] [CrossRef] [PubMed]
- Fidelis, M.; de Oliveira, S.M.; Sousa Santos, J.; Bragueto Escher, G.; Silva Rocha, R.; Gomes Cruz, A.; Araújo Vieira do Carmo, M.; Azevedo, L.; Kaneshima, T.; Oh, W.Y.; et al. From byproduct to a functional ingredient: Camu-camu (Myrciaria dubia) seed extract as an antioxidant agent in a yogurt model. J. Dairy Sci. 2020, 103, 1131–1140. [Google Scholar] [CrossRef]
- Portela, J.L.; Bianchini, M.C.; Boligon, A.A.; Carriço, M.R.S.; Roehrs, R.; Soares, F.A.A.; de Gomes, M.G.; Hassan, W.; Puntel, R.L. Ilex paraguariensis attenuates changes in mortality, behavioral and biochemical parameters associated to methyl malonate or malonate exposure in Drosophila melanogaster. Neurochem. Res. 2019, 44, 2202–2214. [Google Scholar] [CrossRef]
- Weber, V.; Coudert, P.; Rubat, C.; Duroux, E.; Leal, F.; Couquelet, J. Antioxidant properties of novel lipophilic ascorbic acid analogues. J. Pharm. Pharmacol. 2000, 52, 523–530. [Google Scholar] [CrossRef]
- Meščić Macan, A.; Gazivoda Kraljević, T.; Raić-Malić, S. Therapeutic perspective of vitamin C and its derivatives. Antioxidants 2019, 8, 247. [Google Scholar] [CrossRef]
- Kaźmierczak-Barańska, J.; Boguszewska, K.; Adamus-Grabicka, A.; Karwowski, B.T. Two faces of vitamin C-antioxidative and pro-oxidative agent. Nutrients 2020, 12, 1501. [Google Scholar] [CrossRef]
- Chirinos, R.; Galarza, J.; Betalleluz-Pallardel, I.; Pedreschi, R.; Campos, D. Antioxidant compounds and antioxidant capacity of Peruvian camu camu (Myrciaria dubia (H.B.K.) McVaugh) fruit at different maturity stages. Food Chem. 2010, 120, 1019–1024. [Google Scholar] [CrossRef]
- De Souza Schmidt Gonçalves, A.E.; Lajolo, F.M.; Genovese, M.I. Chemical composition and antioxidant/antidiabetic potential of Brazilian native fruits and commercial frozen pulps. J. Agric. Food Chem. 2010, 58, 4666–4674. [Google Scholar] [CrossRef] [PubMed]
- Rufino, M.d.S.M.; Alves, R.E.; de Brito, E.S.; Pérez-Jiménez, J.; Saura-Calixto, F.; Mancini-Filho, J. Bioactive compounds and antioxidant capacities of 18 non-traditional tropical fruits from Brazil. Food Chem. 2010, 121, 996–1002. [Google Scholar] [CrossRef]
- Akter, M.S.; Oh, S.; Eun, J.B.; Ahmed, M. Nutritional compositions and health promoting phytochemicals of camu-camu (Myrciaria dubia) fruit: A review. Food Res. Int. 2011, 44, 1728–1732. [Google Scholar] [CrossRef]
- da Silva, L.A.; Scapim, M.R.d.S.; da Silva, J.F.; Stafussa, A.P.; Aranha, A.C.R.; Jorge, L.M.d.M.; Defendi, R.O.; Júnior, O.d.O.S.; Madrona, G.S. Modeling the extraction of bioactive compounds of green and red camu-camu peel and identification using UPLC-MS/MS. Chem. Eng. Res. Des. 2023, 196, 1–12. [Google Scholar] [CrossRef]
- Grigio, M.L.; Moura, E.A.d.; Chagas, E.A.; Durigan, M.F.B.; Chagas, P.C.; Carvalho, G.F.d.; Zanchetta, J.J. Bioactive compounds in and antioxidant activity of camu-camu fruits harvested at different maturation stages during postharvest storage. Acta Sci. Agron. 2021, 43, e50997. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Macro and Micronutrients | Concentration at 5 g/Product |
|---|---|
| Total carbohydrate | 4 g |
| Vitamin C | 682 mg |
| Potassium | 39 mg |
| Sodium | 15 mg |
| Calcium | 4 mg |
| Total fat | 0 g |
| Protein | 0 g |
| Iron | 0 g |
| Genes | Primers | |
|---|---|---|
| Sense | Antisense | |
| GPDH Gene ID: 33824 | ATGGAGATGATTCGCTTCT | GCTCCTCAATGGTTTTTCCA |
| SOD Gene ID: 36878 | ACCGCACTTCAATCCGTAG | AGTCGGTGATGTTGACCTTG |
| CAT Gene ID: 40048 | ACCAGGGCATCAAGAATCG | AACTTCTTGGCCTGCTCGTA |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Musachio, E.A.S.; Pires, R.G.; Fernandes, E.J.; Andrade, S.; Meichtry, L.B.; Janner, D.E.; Meira, G.M.; Ribeiro, E.E.; Barbisan, F.; Cruz, I.B.M.d.; et al. The Amazonian Camu-Camu Fruit Modulates the Development of Drosophila melanogaster and the Neural Function of Adult Flies under Oxidative Stress Conditions. Antioxidants 2024, 13, 102. https://doi.org/10.3390/antiox13010102
Musachio EAS, Pires RG, Fernandes EJ, Andrade S, Meichtry LB, Janner DE, Meira GM, Ribeiro EE, Barbisan F, Cruz IBMd, et al. The Amazonian Camu-Camu Fruit Modulates the Development of Drosophila melanogaster and the Neural Function of Adult Flies under Oxidative Stress Conditions. Antioxidants. 2024; 13(1):102. https://doi.org/10.3390/antiox13010102
Chicago/Turabian StyleMusachio, Elize Aparecida Santos, Rafaela Garay Pires, Eliana Jardim Fernandes, Stefani Andrade, Luana Barreto Meichtry, Dieniffer Espinosa Janner, Graziela Moro Meira, Euler Esteves Ribeiro, Fernanda Barbisan, Ivana Beatrice Mânica da Cruz, and et al. 2024. "The Amazonian Camu-Camu Fruit Modulates the Development of Drosophila melanogaster and the Neural Function of Adult Flies under Oxidative Stress Conditions" Antioxidants 13, no. 1: 102. https://doi.org/10.3390/antiox13010102
APA StyleMusachio, E. A. S., Pires, R. G., Fernandes, E. J., Andrade, S., Meichtry, L. B., Janner, D. E., Meira, G. M., Ribeiro, E. E., Barbisan, F., Cruz, I. B. M. d., & Prigol, M. (2024). The Amazonian Camu-Camu Fruit Modulates the Development of Drosophila melanogaster and the Neural Function of Adult Flies under Oxidative Stress Conditions. Antioxidants, 13(1), 102. https://doi.org/10.3390/antiox13010102