
Development and Characterization of Inula britannica Extract-Loaded Liposomes: Potential as Anti-Inflammatory Functional Food Ingredients

 , , ,
, , ,
Abstract
:
1. Introduction
2. Materials and Methods
2.1. Preparation of I. britannica Aqueous Extract
2.2. Preparation of Encapsulated I. britannica with Nano-Liposome
2.3. Characterization of Encapsulated I. britannica with Nano-Liposome
2.3.1. Analysis of Particle Size and Stability
2.3.2. Morphological Analysis of Nano-Liposome Encapsulating I. britannica Extract
2.4. Purification of Encapsulated I. britannica with Nano-Liposome
2.5. Determination of Encapsulation Efficiency by Measuring Chlorogenic Acid
2.5.1. Quantification of Chlorogenic Acid in I. britannica Extract by High-Performance Liquid Chromatography (HPLC)
- 0 to 5 min, 10 to 20% B;
- 5 to 10 min, maintaining 20% B;
- 10 to 15 min, 20 to 30% B;
- 15 to 25 min, 30 to 80% B;
- 25 to 28 min, 80 to 10% B;
- 28 to 30 min, holding at 10% B;
2.5.2. Encapsulation Efficiency
2.6. Cell Culture
2.7. Cytotoxicity
2.8. Measurement of Nitric Oxide (NO) Production
2.9. Prostaglandin E2-Enzyme-Linked Immunosorbent Assay (PGE2-ELISA)
2.10. Measurement of Cytokine Expression Using Quantitative Real-Time Polymerase Chain Reaction (qRT-PCR)
2.11. Western Blot
2.12. Statistical Analysis
3. Results
3.1. Characterization of Liposomes Encapsulating I. britannica Extract
3.2. Encapsulation Efficiency of I. britannica Extract in Liposomes
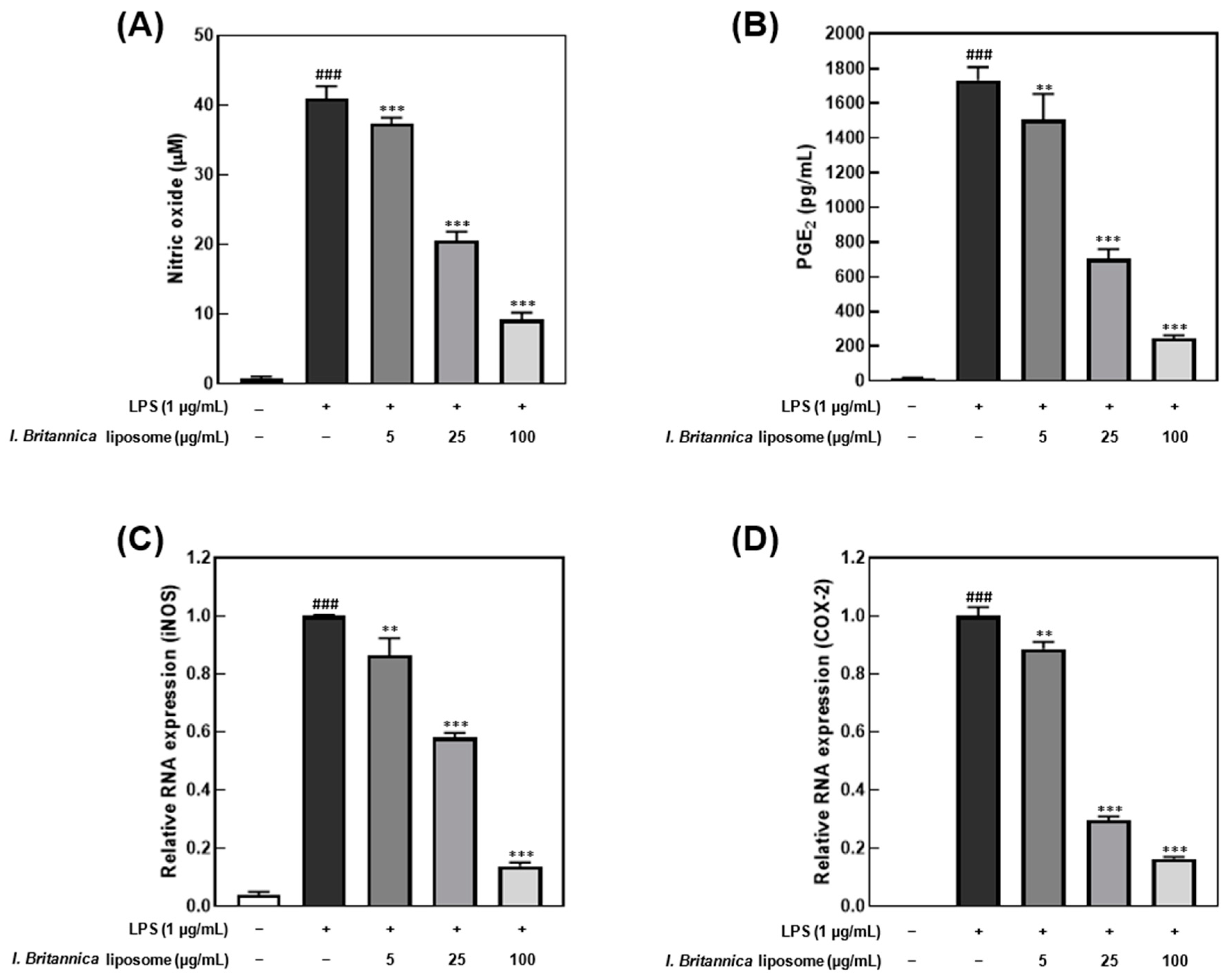
3.3. Effect of I. britannica Extract Encapsulated in Liposomes on LPS-Induced Inflammation in RAW 264.7 Cells
3.4. Effect of I. britannica Extract Encapsulated in Liposomes on Pro-Inflammatory Cytokine Production and mRNA Expression in LPS-Induced RAW 264.7 Cells
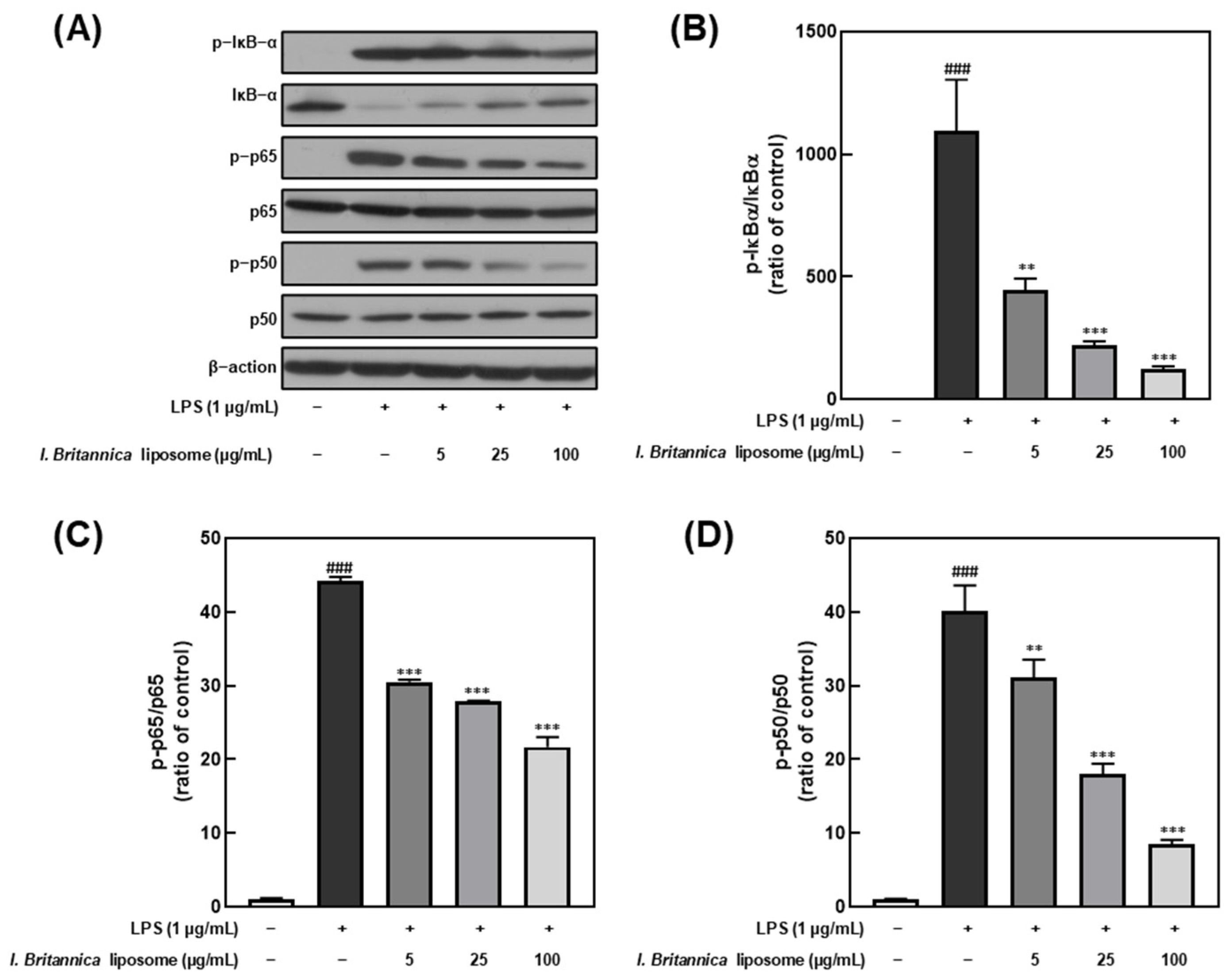
3.5. Effect of I. britannica Extract Encapsulated in Liposomes on NF-κB Activation in LPS-Induced RAW 264.7 Cells
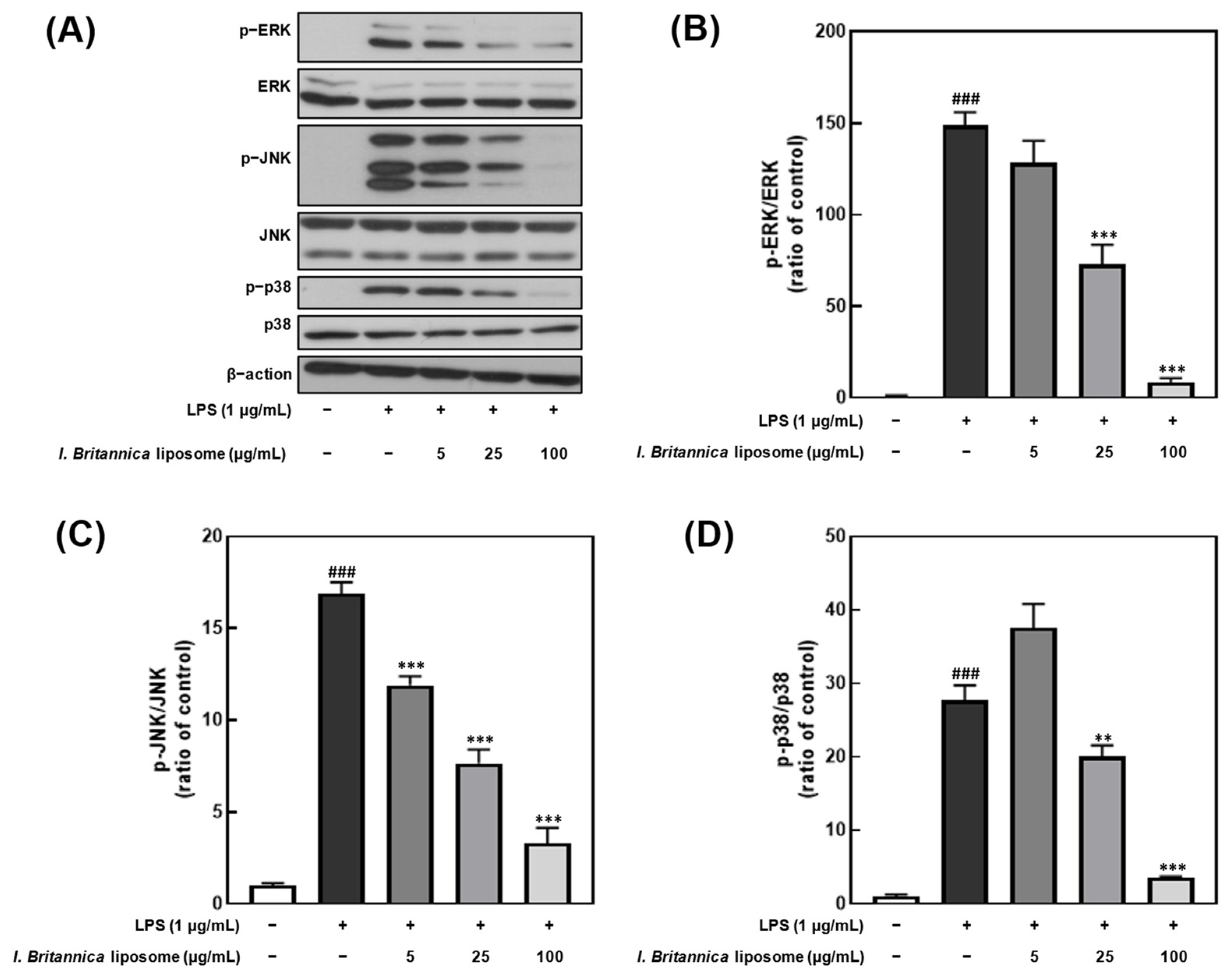
3.6. Effect of I. britannica Extract Encapsulated in Liposomes on the Phosphorylation of MAPKs in LPS-Induced RAW 264.7 Cells
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Ji, M.-Y.; Bo, A.; Yang, M.; Xu, J.-F.; Jiang, L.-L.; Zhou, B.-C.; Li, M.-H. The pharmacological effects and health benefits of Platycodon grandiflorus—A medicine food homology species. Foods 2020, 9, 142. [Google Scholar] [CrossRef]
- Chauhan, B.; Kumar, G.; Kalam, N.; Ansari, S.H. Current concepts and prospects of herbal nutraceutical: A review. J. Adv. Pharm. Technol. Res. 2013, 4, 4–8. [Google Scholar] [CrossRef]
- Zhang, J.; Onakpoya, I.J.; Posadzki, P.; Eddouks, M. The safety of herbal medicine: From prejudice to evidence. Evid.-Based Complement. Altern. Med. 2015, 2015, 316706. [Google Scholar] [CrossRef] [PubMed]
- Khan, A.L.; Hussain, J.; Hamayun, M.; Gilani, S.A.; Ahmad, S.; Rehman, G.; Kim, Y.-H.; Kang, S.-M.; Lee, I.-J. Secondary metabolites from Inula britannica L. and their biological activities. Molecules 2010, 15, 1562–1577. [Google Scholar] [CrossRef] [PubMed]
- Tang, J.-J.; Guo, C.; Peng, X.-N.; Guo, X.-C.; Zhang, Q.; Tian, J.-M.; Gao, J.-M. Chemical characterization and multifunctional neuroprotective effects of sesquiterpenoid-enriched Inula britannica flowers extract. Bioorg. Chem. 2021, 116, 105389. [Google Scholar] [CrossRef] [PubMed]
- Zhang, H.-B.; Wen, J.-K.; Wang, Y.-Y.; Zheng, B.; Han, M. Flavonoids from Inula britannica L. inhibit injury-induced neointimal formation by suppressing oxidative-stress generation. J. Ethnopharmacol. 2009, 126, 176–183. [Google Scholar] [CrossRef]
- Kobayashi, T.; Song, Q.-H.; Hong, T.; Kitamura, H.; Cyong, J.-C. Preventative effects of the flowers of Inula britannica on autoimmune diabetes in C57BL/KsJ mice induced by multiple low doses of streptozotocin. Phytother. Res. 2002, 16, 377–382. [Google Scholar] [CrossRef]
- Xie, L.-Y.; Yang, Z.; Wang, Y.; Hu, J.-N.; Lu, Y.-W.; Zhang, H.; Jiang, S.; Li, W. 1-O-Actylbritannilactone ameliorates alcohol-induced hepatotoxicity through regulation of ROS/Akt/NF-κB-mediated apoptosis and inflammation. ACS Omega 2022, 7, 18122–18130. [Google Scholar] [CrossRef]
- Yu, H.-S.; Kim, W.-J.; Bae, W.-Y.; Lee, N.-K.; Paik, H.-D. Inula britannica inhibits adipogenesis of 3T3-L1 preadipocytes via modulation of mitotic clonal expansion involving ERK 1/2 and Akt signaling pathways. Nutrients 2020, 12, 3037. [Google Scholar] [CrossRef]
- Goelzer Neto, C.F.; do Nascimento, P.; da Silveira, V.C.; de Mattos, A.B.N.; Bertol, C.D. Natural sources of melanogenic inhibitors: A systematic review. Int. J. Cosmet. Sci. 2022, 44, 143–153. [Google Scholar] [CrossRef]
- Umbayev, B.; Askarova, S.; Almabayeva, A.; Saliev, T.; Masoud, A.-R.; Bulanin, D. Galactose-Induced Skin Aging: The Role of Oxidative Stress. Oxid. Med. Cell. Longev. 2020, 2020, 7145656. [Google Scholar] [CrossRef] [PubMed]
- Öksüz, S.; Topcu, G. Triterpene fatty acid esters and flavonoids from Inula britannica. Phytochemistry 1987, 26, 3082–3084. [Google Scholar] [CrossRef]
- Bai, N.; Zhou, Z.; Zhu, N.; Zhang, L.; Quan, Z.; He, K.; Zheng, Q.Y.; Ho, C.-T. Antioxidative Flavonoids from the Flower of Inula britannica. J. Food Lipids 2005, 12, 141–149. [Google Scholar] [CrossRef]
- Bae, W.-Y.; Kim, H.-Y.; Kim, K.-T.; Paik, H.-D. Inhibitory effects of Inula britannica extract fermented by Lactobacillus plantarum KCCM 11613P on coagulase activity and growth of Staphylococcus aureus including methicillin-resistant strains. J. Food Biochem. 2019, 43, e12785. [Google Scholar] [CrossRef] [PubMed]
- Lee, Y.H.; Lee, N.-K.; Paik, H.-D. Antimicrobial characterization of Inula britannica against Helicobacter pylori on gastric condition. J. Microbiol. Biotechnol. 2016, 26, 1011–1017. [Google Scholar] [CrossRef] [PubMed]
- Pan, M.-H.; Chiou, Y.-S.; Cheng, A.-C.; Bai, N.; Lo, C.-Y.; Tan, D.; Ho, C.-T. Involvement of MAPK, Bcl-2 family, cytochrome c, and caspases in induction of apoptosis by 1,6-O,O-diacetylbritannilactone in human leukemia cells. Mol. Nutr. Food Res. 2007, 51, 229–238. [Google Scholar] [CrossRef]
- Chen, H.; Long, Y.; Guo, L. Antiaging effect of Inula britannica on aging mouse model induced by D-galactose. Evid.-Based Complement. Altern. Med. 2016, 2016, 6049083. [Google Scholar] [CrossRef]
- Tang, J.-J.; Huang, L.-F.; Deng, J.-L.; Wang, Y.-M.; Guo, C.; Peng, X.-N.; Liu, Z.; Gao, J.-M. Cognitive enhancement and neuroprotective effects of OABL, a sesquiterpene lactone in 5xFAD Alzheimer’s disease mice model. Redox Biol. 2022, 50, 102229. [Google Scholar] [CrossRef]
- Lobo, V.; Patil, A.; Phatak, A.; Chandra, N. Free radicals, antioxidants and functional foods: Impact on human health. Pharmacogn. Rev. 2010, 4, 118–126. [Google Scholar] [CrossRef]
- Valko, M.; Leibfritz, D.; Moncol, J.; Cronin, M.T.D.; Mazur, M.; Telser, J. Free radicals and antioxidants in normal physiological functions and human disease. Int. J. Biochem. Cell Biol. 2007, 39, 44–84. [Google Scholar] [CrossRef]
- Pham-Huy, L.A.; He, H.; Pham-Huy, C. Free radicals, antioxidants in disease and health. Int. J. Biomed. Sci. 2008, 4, 89–96. [Google Scholar] [PubMed]
- Cordero, M.D.; Alcocer-Gómez, E.; Culic, O.; Carrión, A.M.; de Miguel, M.; Díaz-Parrado, E.; Pérez-Villegas, E.M.; Bullón, P.; Battino, M.; Sánchez-Alcazar, J.A. NLRP3 inflammasome is activated in fibromyalgia: The effect of coenzyme Q10. Antioxid. Redox Signal. 2014, 20, 1169–1180. [Google Scholar] [CrossRef] [PubMed]
- Reuter, S.; Gupta, S.C.; Chaturvedi, M.M.; Aggarwal, B.B. Oxidative stress, inflammation, and cancer: How are they linked? Free Radic. Biol. Med. 2010, 49, 1603–1616. [Google Scholar] [CrossRef] [PubMed]
- Lu, B.; Zhao, Y. Photooxidation of phytochemicals in food and control: A review. Ann. N. Y. Acad. Sci. 2017, 1398, 72–82. [Google Scholar] [CrossRef] [PubMed]
- Li, Z.; Jiang, H.; Xu, C.; Gu, L. A review: Using nanoparticles to enhance absorption and bioavailability of phenolic phytochemicals. Food Hydrocoll. 2015, 43, 153–164. [Google Scholar] [CrossRef]
- Hong, S.-C.; Park, K.-M.; Hong, C.R.; Kim, J.-C.; Yang, S.-H.; Yu, H.-S.; Paik, H.-D.; Pan, C.-H.; Chang, P.-S. Microfluidic assembly of liposomes dual-loaded with catechin and curcumin for enhancing bioavailability. Colloids Surf. A Physicochem. Eng. Asp. 2020, 594, 124670. [Google Scholar] [CrossRef]
- Wang, C.; Piao, J.; Li, Y.; Tian, X.; Dong, Y.; Liu, D. Construction of liposomes mimicking cell membrane structure through frame-guided assembly. Angew. Chem. Int. Ed. 2020, 59, 15176–15180. [Google Scholar] [CrossRef]
- Carugo, D.; Bottaro, E.; Owen, J.; Stride, E.; Nastruzzi, C. Liposome production by microfluidics: Potential and limiting factors. Sci. Rep. 2016, 6, 25876. [Google Scholar] [CrossRef]
- Yang, E.; Yu, H.; Park, J.-Y.; Park, K.-M.; Chang, P.-S. Microfluidic Preparation of Liposomes Using Ethyl Acetate/n-Hexane Solvents as an Alternative to Chloroform. J. Chem. 2018, 2018, 7575201. [Google Scholar] [CrossRef]
- Schneider, C.A.; Rasband, W.S.; Eliceiri, K.W. NIH image to ImageJ: 25 years of image analysis. Nat. Methods 2012, 9, 671–675. [Google Scholar] [CrossRef]
- Fujihara, M.; Muroi, M.; Tanamoto, K.-I.; Suzuki, T.; Azuma, H.; Ikeda, H. Molecular mechanisms of macrophage activation and deactivation by lipopolysaccharide: Roles of the receptor complex. Pharmacol. Ther. 2003, 100, 171–194. [Google Scholar] [CrossRef] [PubMed]
- Sharif, O.; Bolshakov, V.N.; Raines, S.; Newham, P.; Perkins, N.D. Transcriptional profiling of the LPS induced NF-κB response in macrophages. BMC Immunol. 2007, 8, 1. [Google Scholar] [CrossRef] [PubMed]
- Stacey, N.; Winder, C. Toxicity of organic solvents. In Occupational Toxicology; CRC Press: Boca Raton, FL, USA, 2004; pp. 372–398. [Google Scholar]
- Smith, M.C.; Crist, R.M.; Clogston, J.D.; McNeil, S.E. Zeta potential: A case study of cationic, anionic, and neutral liposomes. Anal. Bioanal. Chem. 2017, 409, 5779–5787. [Google Scholar] [CrossRef]
- Xu, X.; Costa, A.; Burgess, D.J. Protein encapsulation in unilamellar liposomes: High encapsulation efficiency and a novel technique to assess lipid-protein interaction. Pharm. Res. 2012, 29, 1919–1931. [Google Scholar] [CrossRef] [PubMed]
- Düzgüneş, N.; Nir, S. Mechanisms and kinetics of liposome–cell interactions. Adv. Drug Deliv. Rev. 1999, 40, 3–18. [Google Scholar] [CrossRef]
- Pisoschi, A.M.; Pop, A. The role of antioxidants in the chemistry of oxidative stress: A review. Eur. J. Med. Chem. 2015, 97, 55–74. [Google Scholar] [CrossRef] [PubMed]
- Persson, T.; Popescu, B.O.; Cedazo-Minguez, A. Oxidative Stress in Alzheimer’s Disease: Why Did Antioxidant Therapy Fail? Oxid. Med. Cell. Longev. 2014, 2014, 427318. [Google Scholar] [CrossRef]
- Rani, V.; Deep, G.; Singh, R.K.; Palle, K.; Yadav, U.C.S. Oxidative stress and metabolic disorders: Pathogenesis and therapeutic strategies. Life Sci. 2016, 148, 183–193. [Google Scholar] [CrossRef]
- Aktan, F. iNOS-mediated nitric oxide production and its regulation. Life Sci. 2004, 75, 639–653. [Google Scholar] [CrossRef]
- Nagy, G.; Clark, J.M.; Buzás, E.I.; Gorman, C.L.; Cope, A.P. Nitric oxide, chronic inflammation and autoimmunity. Immunol. Lett. 2007, 111, 1–5. [Google Scholar] [CrossRef]
- Singh, S.; Gupta, A.K. Nitric oxide: Role in tumour biology and iNOS/NO-based anticancer therapies. Cancer Chemother. Pharmacol. 2011, 67, 1211–1224. [Google Scholar] [CrossRef] [PubMed]
- Vane, J.R.; Bakhle, Y.S.; Botting, R.M. Cyclooxygenases 1 and 2. Annu. Rev. Pharmacol. Toxicol. 1998, 38, 97–120. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.M.; An, J. Cytokines, inflammation, and pain. Int. Anesthesiol. Clin. 2007, 45, 27–37. [Google Scholar] [CrossRef] [PubMed]
- Zelová, H.; Hošek, J. TNF-α signalling and inflammation: Interactions between old acquaintances. Inflamm. Res. 2013, 62, 641–651. [Google Scholar] [CrossRef] [PubMed]
- Hirano, T. IL-6 in inflammation, autoimmunity and cancer. Int. Immunol. 2020, 33, 127–148. [Google Scholar] [CrossRef]
- Weber, A.; Wasiliew, P.; Kracht, M. Interleukin-1β (IL-1β) processing pathway. Sci. Signal. 2010, 3, 1–2. [Google Scholar] [CrossRef]
- Lawrence, T.; Gilroy, D.W.; Colville-Nash, P.R.; Willoughby, D.A. Possible new role for NF-κB in the resolution of inflammation. Nat. Med. 2001, 7, 1291–1297. [Google Scholar] [CrossRef]
- Reddy, D.B.; Reddanna, P. Chebulagic acid (CA) attenuates LPS-induced inflammation by suppressing NF-κB and MAPK activation in RAW 264.7 macrophages. Biochem. Biophys. Res. Commun. 2009, 381, 112–117. [Google Scholar] [CrossRef]
- Johnson, G.L.; Lapadat, R. Mitogen-activated protein kinase pathways mediated by ERK, JNK, and p38 protein kinases. Science 2002, 298, 1911–1912. [Google Scholar] [CrossRef]
- Kim, H.-G.; Yoon, D.-H.; Kim, C.-H.; Shrestha, B.; Chang, W.-C.; Lim, S.-Y.; Lee, W.-H.; Han, S.-G.; Lee, J.-O.; Lim, M.-H. Ethanol extract of Inonotus obliquus inhibits lipopolysaccharide-induced inflammation in RAW 264.7 macrophage cells. J. Med. Food 2007, 10, 80–89. [Google Scholar] [CrossRef]
- Park, H.-H.; Kim, M.J.; Li, Y.; Park, Y.N.; Lee, J.; Lee, Y.J.; Kim, S.-G.; Park, H.-J.; Son, J.K.; Chang, H.W.; et al. Britanin suppresses LPS-induced nitric oxide, PGE2 and cytokine production via NF-κB and MAPK inactivation in RAW 264.7 cells. Int. Immunopharmacol. 2013, 15, 296–302. [Google Scholar] [CrossRef] [PubMed]
- Santa-Cecília, F.V.; Socias, B.; Ouidja, M.O.; Sepulveda-Diaz, J.E.; Acuña, L.; Silva, R.L.; Michel, P.P.; Del-Bel, E.; Cunha, T.M.; Raisman-Vozari, R. Doxycycline Suppresses Microglial Activation by Inhibiting the p38 MAPK and NF-kB Signaling Pathways. Neurotox. Res. 2016, 29, 447–459. [Google Scholar] [CrossRef] [PubMed]



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Diameter Size (nm) | Polydiversity (PdI) | Z-Potential (mV) |
|---|---|---|
| 101.3 ± 5.6 | 0.012 ± 0.007 | −42.2 ± 0.7 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hong, C.R.; Lee, E.H.; Jung, Y.H.; Lee, J.-H.; Paik, H.-D.; Hong, S.-C.; Choi, S.J. Development and Characterization of Inula britannica Extract-Loaded Liposomes: Potential as Anti-Inflammatory Functional Food Ingredients. Antioxidants 2023, 12, 1636. https://doi.org/10.3390/antiox12081636
Hong CR, Lee EH, Jung YH, Lee J-H, Paik H-D, Hong S-C, Choi SJ. Development and Characterization of Inula britannica Extract-Loaded Liposomes: Potential as Anti-Inflammatory Functional Food Ingredients. Antioxidants. 2023; 12(8):1636. https://doi.org/10.3390/antiox12081636
Chicago/Turabian StyleHong, Chi Rac, Eun Ha Lee, Young Hoon Jung, Ju-Hoon Lee, Hyun-Dong Paik, Sung-Chul Hong, and Seung Jun Choi. 2023. "Development and Characterization of Inula britannica Extract-Loaded Liposomes: Potential as Anti-Inflammatory Functional Food Ingredients" Antioxidants 12, no. 8: 1636. https://doi.org/10.3390/antiox12081636
APA StyleHong, C. R., Lee, E. H., Jung, Y. H., Lee, J.-H., Paik, H.-D., Hong, S.-C., & Choi, S. J. (2023). Development and Characterization of Inula britannica Extract-Loaded Liposomes: Potential as Anti-Inflammatory Functional Food Ingredients. Antioxidants, 12(8), 1636. https://doi.org/10.3390/antiox12081636