Abstract
Salt stress can cause cellular dehydration, which induces oxidative stress by increasing the production of reactive oxygen species (ROS) in plants. They may play signaling roles and cause structural damages to the cells. To overcome the negative impacts, the plant ROS scavenging system plays a vital role in maintaining the cellular redox homeostasis. The special sugar beet apomictic monosomic additional M14 line (BvM14) showed strong salt stress tolerance. Comparative proteomics revealed that six antioxidant enzymes (glycolate oxidase (GOX), peroxiredoxin (PrxR), thioredoxin (Trx), ascorbate peroxidase (APX), monodehydroascorbate reductase (MDHAR), and dehydroascorbate reductase3 (DHAR3)) in BvM14 were responsive to salt stress. In this work, the full-length cDNAs of genes encoding these enzymes in the redox system were cloned from the BvM14. Ectopic expression of the six genes reduced the oxidative damage of transgenic plants by regulating the contents of hydrogen peroxide (H2O2), malondialdehyde (MDA), ascorbic acid (AsA), and glutathione (GSH), and thus enhanced the tolerance of transgenic plants to salt stress. This work has charecterized the roles that the antioxidant enzymes play in the BvM14 response to salt stress and provided useful genetic resources for engineering and marker-based breeding of crops that are sensitive to salt stress.
1. Introduction
Wild sugar beet (Beta corolliflora Zoss.) has excellent characteristics of drought resistance, frost resistance, salt tolerance, cold tolerance, and apomixis. In the early stage of the study, diploid cultivated beet (B. vulgaris L.) and tetraploid wild sugar beet (B. corolliflora Zoss.) were crossed by distant hybridization. After obtaining allotriploid, they were further backcrossed with cultivated sugar beet, and the M14 with the wild sugar beet chromosome 9 (BvM14) was selected for apomixix and high salt tolerance. It is a rare germplasm for studying plant salt stress tolerance mechanisms [1,2,3,4,5].
Reactive oxygen species (ROS), including superoxide anions (O2•−), hydroxyl radicals (HO2•), hydrogen peroxide (H2O2), and singlet oxygen (1O2), play an important role in plant metabolism, signal transduction, photosynthesis regulation, bacterial defense, and cell apoptosis [6,7,8]. Salinity affects plants growth and development through osmotic stress, ion toxicity, overproduction of ROS, and oxidative stress [9]. It is known that chloroplasts, mitochondria, peroxisomes, apoplast, and plasma membranes are the main sites of cellular ROS generation [10]. Overproduction of ROS can lead to severe damage of protein, membrane lipid, DNA, and other cellular components [11]. To cope with this challenge, plants have antioxidative mechanisms that consist of enzymatic and non-enzymatic components to regulate ROS synthesis and scavenging. The antioxidant enzymes of ROS scavenging in plants mainly include superoxide dismutase (SOD), ascorbate perxidase (APX), catalase (CAT), glutathione peroxidase (GPX), and thioredoxin (Trx) [12,13,14]. Some antioxidative genes have been cloned from rice [12], Arabidopsis [15], maize [14], and soybean [16], while only partial sequences of peroxisome APX gene coding region and GOX gene have been obtained from salt-tolerant sugar beet [17]. However, there are few reports about the different roles of these enzymes under salt stress.
In the past years, the salt tolerance characteristics of BvM14 have been well-studied [1,2,4,18,19,20]. The differentially expressed proteins (DEPs) under salt stress (0, 200, 400 mM) have been studied by iTRAQ LC-MS/MS. A total of 76 DEPs have been identified in leaves of the BvM14. These proteins involve photosynthesis, metabolism, protein synthesis, protein folding and degradation, stress and defense, cell structure, transcription, and transport processes. Among them, six main proteins (GOX, PrxR, Trx, APX, DHAR3, and MDHAR) of the antioxidant enzymes system changed most significantly in the ROS scavenging system under salt stress [4]. However, to the best of our knowledge, salt tolerance of the GOX, PrxR, Trx, APX, DHAR3, and MDHAR genes and their relationships have not been characterized in sugar beet M14.
In this work, we aimed to investigate the following related questions. What are the reactions that are directly responsible for these six genes in the antioxidant enzyme system during salt stress? What is the relationship between these six genes in antioxidant enzyme system under salt stress conditions? How important are the six genes in antioxidant enzyme system during salt stress? To answer these questions, molecular biological methods were used to evaluate the functions of the six major antioxidant enzymes in BvM14 under salt stress [3,4]. We generated a complete set of single mutants for the six key genes in Arabidopsis and analyzed the function of the six genes under salt stress. Understanding the mechanism of ROS scavenging allows for a powerful strategy to enhance crop salt tolerance.
2. Materials and Methods
2.1. Plant Materials, and Growth Conditions
Sugar beet M14 seeds were initially sowed in vermiculite for seven days. Then, the seedlings were planted in Hoagland solution and kept at 23 °C, 450 μmol.m−2. s−1, and 14 h/10 h light/dark cycles [3,4]. Arabidopsis thaliana (Columubia 0) seeds were treated with 5% (w/v) NaClO for 6 min, rinsed with sterilized water to remove NaClO. Arabidopsis plants were grown in soil under controlled conditions (22 °C, 300 μmol m−2 s−1 and 16 h/8 h light/dark cycles) [21].
2.2. Isolation and Sequence Analysis of Genes
Total RNA was isolated and subjected to reverse transcription by using the SuperScriptTM III Reverse Transcriptase kit (Invitrogen, Carlsbad, CA, USA). The full-length coding regions were PCR-amplified from the sugar beet M14 cDNA with gene specific primers (Supplementary Table S1).
2.3. Quantitative Real-Time PCR Analysis
Quantitative real-time PCR analysis (qRT-PCR) analyses were performed using SYBR Premix ExTaqTM II Mix (TaKaRa, Shiga, Japan). GAPDH (glyceraldehyde-3-phosphate dehydrogenase, accession no. DQ355800) was used as a reference. The expression levels of all candidate genes were analyzed by the 2−ΔΔCT CT method. The primers used for qRT-PCR were listed in Supplementary Table S1 [1,2,22].
2.4. Generation and Phenotypic Analyses of Transgenic A. thaliana
At present, the genetic transformation system in sugar beet is not successful, so the gene function research will be carried out in A. thaliana. Currently, our team is experimenting with various approaches to the genetic transformation system in sugar beet.
We obtained the mutants gox, prxr, trx, apx, mdhar, and dhar3 from the Arabidopsis Biological Resource Center (ABRC), genotyping PCR was performed to identify the T-DNA insertion. To construct 35S::BvM14-GOX, 35S:: BvM14-PrxR, 35S:: BvM14-Trx, 35S::APX, 35S:: BvM14-MDHAR, and 35S:: BvM14-DHAR3 in Arabidopsis, the open-reading frames (ORF) were cloned into pCAMBIA1305.1, and transformed into Arabidopsis as described [23]. The T0 Arabidopsis seeds were screened on MS plates [21] containing 30 mg/L hygromycin, and the survival seedlings were further verified by reverse transcription polymerase chain reaction (RT-PCR) analyses. Homozygous T3 generation plants were used for further analyses [21].
2.5. Quantification of Biomass, MDA Content, H2O2 Content, Na+ and K+ Content, Morphological Index, and Physiological Indicators
The root length was measured in Murashige and Skoog (MS) medium. The 7-day-old seedlings were transferred to MS medium containing 150 mM NaCl for salt treatment 10 days. Three biological replicates were carried out.
The fresh weight and dry weight and physiological indicators of leaves were measured in the soil. The one-month wild type (WT) and transgenic plants were treated with 150 mM NaCl for 7 days.
The MDA content was measured as described by Wang [21]. The contents of H2O2 were measured by H2O2 assay kit (Comin Botechnology, Suzhou, China), the contents of glutathione (GSH), and ascorbic acid (AsA) were determined using the GSH and AsA assay kits, respectively (Comin Botechnology, Suzhou, China). The activities of catalase (CAT), ascorbate peroxidase (APX), glycolate oxidase (GOX), peroxiredoxins (PrxR), and thioredoxins (Trx) were determined in WT and transgenic leaves using an assay kit (Comin Botechnology, Suzhou, China). The activities of dehydroascorbate reductase (DHAR) and monodehydroascorbate reductase (MDHAR) were determined using an ELISA assay kit (Mlbio, Shanghai, China).
2.6. Cis-Regulatory Elements(CREs) Analysis
The promoters of the six stress-responsive genes in BvM14 were analyzed for putative cis-elements using available genomic sequences and our transcriptomic data [19]. In addition, the 2000 bp genomic sequences located on the 5′ upstream of the Transcriptional Start Site (TSS) of the six stress-responsive genes sequences were extracted and analyzed with PlantCARE (https://bioinformatics.psb.ugent.be/webtools/plantcare/html/, accessed on 15 November 2022) and TB tools. The Promoter binding of transcription factors was predicted by PlantTFDB (http://planttfdb.gao-lab.org/prediction.php, accessed on 3 June 2022) analysis [16].
2.7. Statistical Analysis
For all the experiments, three biological replicates with three technical replicates of each treatment were measured. All data were analyzed using GraphPad Prism 8 (Dr. Harvey Motulsky, San Diego, CA, USA). For multiple comparions, one-way analysis of variance (ANOVA) was used to determine statistical significance among treatments at p < 0.05.
3. Results
3.1. Ectopic Expression of BvM14 Antioxidant Enzymes Enhanced Plant Salt Tolerance
Ectopic expression of the six genes encoding the antioxidant enzymes promoted root elongation of transgenic plants under salt stress. With WT plants as control, the root length of plants overexpressing (OX) the BvM14-Trx, BvM14-PrxR, and BvM14-APX increased significantly by 1.3-fold, 1.2-fold, and 1.2-fold after 150 mM NaCl treatment. Compared with knock-out (KO) mutant plants of Trx, PrxR, and APX genes, the root length of complementation transgenic lines (CO) was restored by complementation with the BvM14-Trx, BvM14- PrxR, and BvM14-APX genes to the levels of OX plants (Figure 1). Plants overexpressing other enzyme-coding genes also showed similar phenotypes (Supplementary Figure S1).
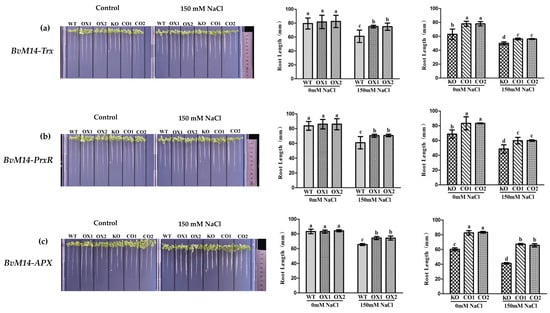
Figure 1.
Effects of salt stress on the root length of seedings with different genotypes (WT, OX, KO, and CO): (a) Root length of WT and Trx-related plants under salt stress; (b) root length of of WT and PrxR-related plants under salt stress; (c) root length of WT and APX-related plants under salt stress. Data were analyzed by Duncan’s analysis of variance, and different lowercase letters (a, b, c, d) indicate differences in root length.
The overexpression of these six genes increased the fresh and dry weights of the OX plants under salt stress. Compared to the WT plants, the fresh and dry weights of the plants overexpressing BvM14-APX, BvM14-DHAR3, and BvM14-MDHAR genes increased by 1.2-fold, 1.3-fold, and 1.2-fold under 150 mM NaCl stress, respectively, the fresh and dry weights of CO plants were restored, in contrast to the KO plants with mutated APX, DHAR3, and MDHAR genes (Figure 2). Similar phenotypic changes were observed for other OX lines of the antioxidant enzyme-coding genes (Supplementary Figure S2).
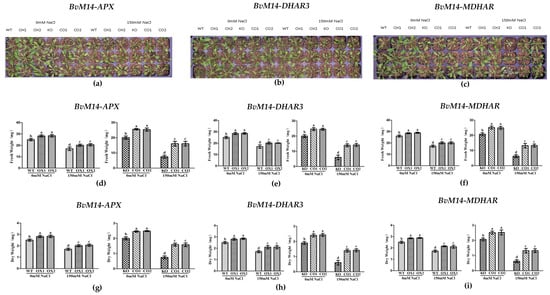
Figure 2.
Effects of salt stress on the phenotype and biomass: (a) Phenotype of WT and APX-related plants under salt stress; (b) phenotype of of WT and DHAR3-related plants under salt stress; (c) phenotype of WT and MDHAR-related plants under salt stress; (d,g) biomass of WT and APX-related plants under salt stress; (e,h) biomass of of WT and DHAR3-related plants under salt stress; (f,i) biomass of WT and MDHAR-related plants under salt stress. Data were analyzed by Duncan’s analysis of variance, and different lowercase letters (a, b, c, d) indicate differences in fresh and dry weight.
3.2. Ectopic Expression of Genes Encoding the Antioxidant Enzymes of BvM14 Improved Plant Antioxidant Capacity
The H2O2 contents in the WT and OX plants of BvM14-APX, BvM14-DHAR3, BvM14-PrxR, BvM14-Trx, and BvM14-MDHAR were similar under normal conditions. However, the results became significantly different under salt stress (Figure 3). Interestingly, except for OX plants of BvM14-GOX, the rest of the OX plants showed less H2O2 (Figure 3). Compared to CO lines, the H2O2 contents in the KO of the genes APX, DHAR3, PrxR, Trx, and MDHAR showed significantly elevated by1.5-fold, 2.0-fold, 1.2-fold, 1.3-fold, and 1.6-fold under salt stress, respectively.
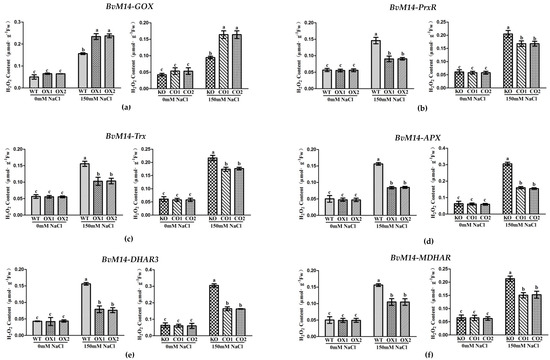
Figure 3.
H2O2 contents of WT and GOX, PrxR, Trx, APX, DHAR3, and MDHAR transgenic plants under salt stress: (a) GOX-related plants; (b) PrxR-related plants; (c) Trx-related plants; (d) APX-related plants; (e) DHAR3-related plants; (f) MDHAR-related plant. Data were analyzed by Duncan’s analysis of variance, and different lowercase letters (a, b, c) indicate differences in H2O2 contents.
MDA concentrations were noticeably reduced in BvM14-DHAR3, BvM14-GOX, and BvM14-MDHAR OX compared with WT plants, but there were no significant changes in other OX plants under salt stress conditions, MDA concentrations were significantly elevated in MDHAR KO compared with CO plants (Supplementary Figure S3).
The contents of AsA were similar in the BvM14-Trx, BvM14-GOX, and BvM14-PrxR OX plant leaves compared with WT leaves under salt stress. However, the contents of AsA were increased by 1.8-fold and 1.5-fold in the BvM14-DHAR3 and BvM14-MDHAR OX plants, while the content of AsA was decreased by 0.73-fold in the BvM14-APX OX plants under salt stress (Supplementary Figure S4). The content of GSH was significantly induced by 1.7-fold in the BvM14-MDHAR OX plant leaves compared with WT leaves and reduced by 0.7-fold in the MDHAR KO plant leaves compared with CO leaves under salt stress. However, the content of GSH was decreased by 0.6-fold in the BvM14-DHAR3 OX plants under salt stress. The other transgenic plants did not show significant changes under salt stress (Figure 4).
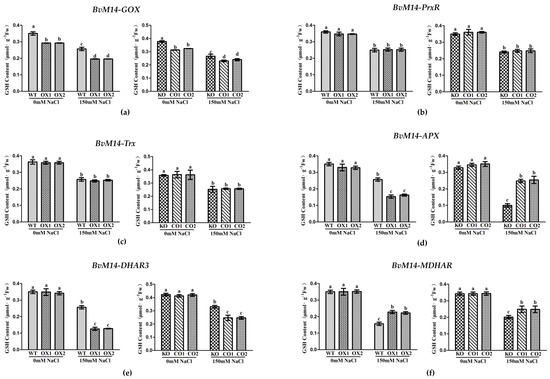
Figure 4.
The GSH contents of WT and GOX, PrxR, Trx, APX, DHAR3, and MDHAR transgenic plants under salt stress: (a) GOX-related; (b) PrxR-related; (c) Trx-related; (d) APX-related; (e) DHAR3-related; (f) MDHAR-related plants. Data were analyzed by Duncan’s analysis of variance, and different lowercase letters (a, b, c, d) indicate differences in GSH contents.
3.3. Antioxidant Enzyme Activities of WT, Different KO, and Transgenic Plants under Salt Stress
Compared with WT plants, overexpression of BvM14-GOX gene increased significantly in gene expression levels and the antioxidant enzyme activities under 150 mM NaCl stress. The Trx and PrxR activities in BvM14-GOX gene OX lines were significantly increased by 1.4-fold and 1.2-fold relative to WT under salt stress (Figure 5).
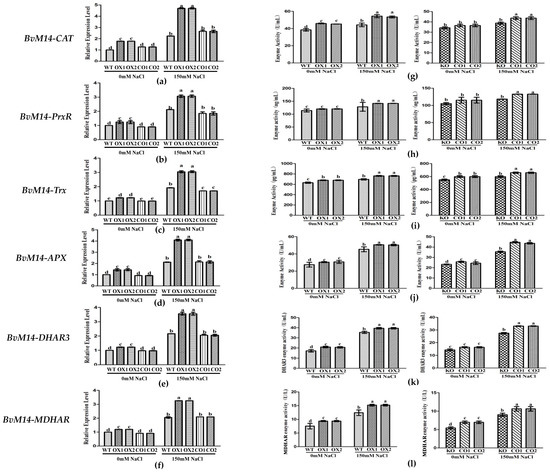
Figure 5.
Analyses of relative gene expression and enzyme activities in GOX-related plants under salt stress: (a–f) Expression of antioxidative genes (BvM14-CAT, BvM14-PrxR, BvM14-Trx, BvM14-APX, BvM14-DHAR3, and BvM14-MDHAR) in WT and GOX-related plants under salt stress; (g–l) antioxidant enzyme activities (BvM14-CAT, BvM14-PrxR, BvM14-Trx, BvM14-APX, BvM14-DHAR3, and BvM14-MDHAR) in WT and GOX-related plants under salt stress. Data were analyzed by Duncan’s analysis of variance, and different lowercase letters (a, b, c, d, e) indicate differences in gene expression and enzyme activities.
The Trx and PrxR enzyme activities in BvM14-PrxR gene OX lines and BvM14-Trx gene OX lines were significantly higher by 1.5-fold and 1.3-fold than WT under salt stress, respectively (Figure 6 and Supplementary Figure S5). Therefore, it is inferred that the two genes have synergistic effects in plant tolerance to salt stress.
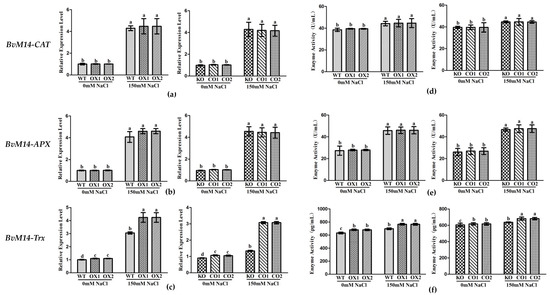
Figure 6.
Analyses of relative gene expression and enzyme activities in PrxR-related plants under salt stress: (a–c) Analysis of the expression of antioxidative genes (BvM14-CAT, BvM14-APX, and BvM14-Trx) in WT and PrxR-related plants under salt stress; (d–f) antioxidant enzyme activity (BvM14-CAT, BvM14-APX, and BvM14-Trx) in WT and PrxR-related plants under salt stress. Data were analyzed by Duncan’s analysis of variance, and different lowercase letters (a, b, c, d) indicate differences in gene expression and enzyme activities.
In BvM14-APX OX lines, the MDHAR and DHAR3 enzyme activity was positively correlated with that of the APX, and both increased by 1.9-fold and 1.3-fold under salt stress. The MDHAR and DHAR3 gene expression levels was increased by 2.3-fold and 2.1-fold in BvM14-APX OX lines, and both reduced by 0.6-fold and 0.6-fold in APX KO lines under salt stress (Figure 7).
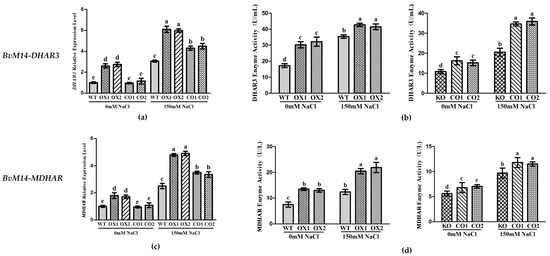
Figure 7.
Analyses of relative gene expression and enzyme activities in APX-related plants under salt stress: (a,c) Analysis of the expression of antioxidative genes (BvM14-DHAR3 and BvM14-MDHAR) in WT and APX-related plants under salt stress; (b,d) antioxidant enzyme activity (BvM14-DHAR3 and BvM14-MDHAR) in WT and APX-related plants under salt stress. Data were analyzed by Duncan’s analysis of variance, and different lowercase letters (a, b, c, d, e) indicate differences in gene expression and enzyme activities.
Both in BvM14-DHAR3 and BvM14-MDHAR OX lines, the APX enzyme activity was significantly higher by 1.5-fold and 1.3-fold than WT, in DHAR3 and MDHAR KO lines, the APX enzyme activity was significantly reduced by 0.3-fold and 0.7-fold relative to CO under salt stress (Supplementary Figure S6). Increased DHAR and MDHAR activities were reported in different plants subjected to abiotic stresses [24,25,26].
3.4. Different Levels of Regulations of the Six Antioxidant Enzymes of BvM14
Overexpression of BvM14-GOX gene significantly increased the expression and activity of Trx and PrxR enzymes in antioxidant system (Figure 5). The BvM14-Trx gene expression and enzyme activity showed the same trend in BvM14-PrxR transgenic plants. Compared with BvM14-APX KO plants, CO plants with BvM14-APX gene restored significantly increased BvM14-DHAR3 gene and BvM14-DHAR3 gene expression and enzyme activity after 150 mM NaCl stress. In BvM14-DHAR3 transgenic plants, BvM14-APX gene expression and enzyme activity were both increased, the BvM14-MDHAR gene expression and enzyme activity were consistent with BvM14-APX. The expression of BvM14-APX gene and enzyme activity existed positive correlation to BvM14-MDHAR, but the expression of BvM14-DHAR3 gene and enzyme activity showed the opposite trend (Figure 8a).
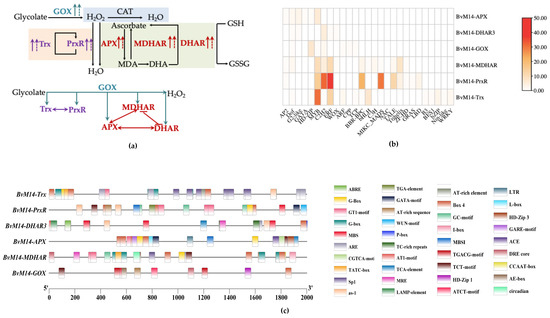
Figure 8.
Regulatory networks of the six antioxidant enzymes in the BvM14: (a) The role of BvM14-GOX, BvM14-PrxR, BvM14-Trx, BvM14-APX, BvM14-DHAR3, and BvM14-MDHAR gene in antioxidant enzyme system; (b) the TF binding motifs in the promoter regions of BvM14-GOX, BvM14-PrxR, BvM14-Trx, BvM14-APX, BvM14-DHAR3, and BvM14-MDHAR; (c) localization of the more frequent cis-acting elements among the BvM14-GOX, BvM14-PrxR, BvM14-Trx, BvM14-APX, BvM14-DHAR3, and BvM14-MDHAR in the promoter regions. The cis-regulatory elements presented are labled with different colors and illustrated on the right side. The TATA-box and CAAT-box are not shown. (The dashed line indicates the transcript level and the solid line indicates the activity level; the arrows indicates positive; the lines indicates negative).
To understand the potential transcriptional regulatory mechanisms of BvM14-GOX, BvM14-Trx, BvM14-PrxR, BvM14-APX, BvM14-DHAR3, and BvM14-MDHAR genes, we analyzed the presence of cis-regulating elements 2000 bp upstream the TSS. The common cis-acting elements (such as enhancer element CAAT-box and core promoter element TATA-box) were not shown in the results, the most abundant elements were salt stress-responsive elements, including ABA-responsive element (ABRE), GT-1 motifs that were present in BvM14-Trx, BvM14-PrxR, BvM14-APX, BvM14-DHAR3, and BvM14-MDHAR. Moreover, we detected hormone-related cis-acting elements, including the jasmonate-responsive element (TGACG motif and CGTCA motif), and the salicylic acid-responsive element (TCA element), as widespread among the BvM14-Trx, BvM14-PrxR, BvM14-APX, BvM14-DHAR3, BvM14-GOX, BvM14-GOX, BvM14-DHAR3, and BvM14-APX. This suggests that these genes are regulated by abiotic stresses and hormones. The binding sites for the salt stress-responsive TFs MYB and APETALA2/ethylene-responsive element binding factors (AP2/ERF) were found in BvM14-Trx, BvM14-PrxR, and BvM14-MDHAR3 (Figure 8b).
4. Discussion
Based on the previous transcriptome database of BvM14 under salt stress, the open-reading frames (ORF) of the coding genes BvM14-GOX, BvM14-PrxR, BvM14-Trx, BvM14-APX, BvM14-MDHAR, BvM14-DHAR3 encoding major enzymes in the antioxidant system were cloned and analyzed. Arabidopsis KO mutants of the homologous genes, CO lines, OX lines, and WT plants were used to characterize the roles of these enzymes in plant salt stress tolerance. Enzyme activity and transcriptional level of six major enzymes in transgenic lines were detected.
ROS (mainly H2O2 and O2•−) accumulation can be used as an important indicator for cellular oxidative stress [27]. MDA content can reflect the degree of membrane lipid peroxidation in plant [28]. The results showed that overexpression of the six genes in Arabidopsis increased the tolerance of the OX plants to salt stress, which may be attributed to enhanced antioxidante capacity and reduced oxidative damage by decreasing the contents of H2O2 and MDA and increasing the AsA and GSH contents. Further analysis showed that overexpression of the BvM14-GOX led to significant increases in the expression levels and enzyme activities of other key enzymes in the antioxidant enzyme system. DHAR recycles ascorbic acid (AsA), which is then oxidized to form MDHAR. MDHAR is further converted to dehydroascorbate (DHA). AsA is essential to main the cellular redox state under abiotic stresses. MDHAR accompanies APX and scavenges H2O2 in the mitochondria and peroxisome [29,30,31,32,33,34]. For example, the BvM14-Trx and the BvM14-PrxR mutually promote each other; the expression level and enzyme activity of BvM14-APX were positively correlated with the expression levels and enzyme activities of BvM14-DHAR3 and BvM14-MDHAR, but the expression level and enzyme activity of BvM14-MDHAR were negatively correlated with the expression level and enzyme activity of BvM14-DHAR3; the PrxR/Trx pathway had no significant interaction with the CAT pathway or the AsA/GSH pathway and participated in the plant antioxidant process independently. Based on these results, a salt stress response regulatory network of BvM14 antioxidant system was constructed.
GOX plays an important role in the glycolate-glyoxylate conversion during photorespiration, which catalyzes the oxidation of glycolate to generate glyoxylate and H2O2 [35,36,37,38]. BvM14-GOX regulates the activity of other key enzymes in the antioxidant enzyme system and the expression of corresponding genes by catalyzing the production of H2O2 from glycolic acid, the regulation of H2O2 may occur in a fluctuating manner because the association-dissociation of GOX and CAT could take place dynamically and transiently in response to environmental stresses or stimuli. Consistently, related research showed that in spite of the constant and high production of ROS caused by the transgenic GOX in rice, but it can assist innate antioxidative systems in modulating ionic and redox homeostasis for salt stress tolerance [39], so as to improve the antioxidant capacity of plants and reduce the inhibition of salt stress on plant growth and development [35].
PrxR/Trx pathway-related coding genes PrxR and Trx have a vital function in cellular antioxidative defense via eliminating excessive ROS [15,40,41,42]. BvM14-Trx gene and BvM14-PrxR gene act synergistically to eliminate excess of H2O2 in plants, but they do not participate in MDA and GSH metabolic pathways since OX plants did not show differences in AsA/GSH pathway and CAT pathway. In Arabidopsis, AtTrxh2 overexpressing transgenic plants exhibited higher activities of antioxidant enzymes including peroxidase (POD), catalase (CAT), and superoxide dismutase (SOD), compared with the plants expressing the empty vector control [43]. In tomato, SlTrxh enhanced nitrate stress tolerance with decreased oxidative damage by increased antioxidant enzyme activities and interacted with SlPrx [42]. The AsA-GSH cycle is one of the important antioxidant systems in plants [44,45]. APX is involved in the initial step of the AsA-GSH cycle that scavenges excess ROS and protects the plant from salt stress [46,47]. In AsA/GSH pathway, BvM14-APX expression and enzyme activity were positively correlated with BvM14-DHAR3 and BvM14-MDHAR expression and enzyme activity, while BvM14-MDHAR expression and enzyme activity were negatively correlated with BvM14-DHAR3 expression and enzyme activity. In tobacco, overexpression of MnSOD, MDHAR, DHAR, and CAT in transgenic plants exhibited the improvement of salt tolerance [29,48]. The improved MnSOD, CAT, POD, APX, DHAR, MDHAR, and GR expression was also detected in wheat lines and two Chrysanthemum species under cold acclimation [49], which was similar with our results. PrxR/Trx pathway has no obvious interaction with CAT pathway and AsA/GSH pathway in plant antioxidant enzymes system.
The distribution and type of CREs in promoters affect the activities and functions of genes. In this study, through a systematic analysis of CRE in the promoter regions of the six genes, we identified various types of CRE (Figure 8c). Related to salt stress, previous studies have shown that abscisic acid (ABA) responsive element binding protein (AREB)/ABRE binding factors (ABFs) in bZIP transcription factors were involved in salt stress [50,51]. Other research indicated that the GT-1 element directly controls the salt response of OsRAV2. The study provided a better understanding of the putative functions of OsRAVs and the molecular regulatory mechanisms of plant genes under salt stress [52,53]. In this work, we found the CREs in promoters of the six genes can bind many transcription factors, it contains transcription factors (e.g., MYB, C2H2, ERF) related to salt stress (Figure 8b). Recent studies suggested that the AP2/ERF TF family are involved in abiotic stress adaptation [54,55]. Functional analysis of the SmAP2-17 gene confirmed its role in plant salt tolerance [56]. It has also been demonstrated that C2H2 zinc finger proteins and MYB transcription factors play vital roles in biotic and abiotic stress tolerance [57].
5. Conclusions
In conclusion, we characterized the key genes encoding the major antioxidant enzymes of ROS scavenging system in plant salt stress tolerance. In different pathways, the key enzymes synergistically or antagonistically play important role in plant salt stress tolerance. In the PrxR/Trx pathway, the BvM14-Trx and the BvM14-PrxR mutually promote each other, but the BvM14-Trx gene and BvM14-PrxR gene do not affect AsA/GSH pathway and CAT pathway. In the AsA/GSH pathway, BvM14-APX expression and enzyme activity were positively correlated with BvM14-DHAR3 and BvM14-MDHAR expression and enzyme activity. Meanwhile, the CREs in promoters contain salt stress-responsive elements ABRE, GT-1 motifs and the CREs in promoters of the six genes can bind transcription factor MYB, C2H2, and ERF related to salt stress. Based on the working model, it became clear that multiple levels of regulations, including transcription and translation, are important in controlling plant salt stress tolerance. Future efforts in improving crop salt stress tolerance can benefit from the results from this study and need to consider multiple genes and markers to achieve optimal outcomes.
Supplementary Materials
The following supporting information can be downloaded at: https://www.mdpi.com/article/10.3390/antiox12010057/s1, Table S1: List of the primer sequences for the six genes tested by RT-PCR; Figure S1: Effects of salt stress on the root length of seedlings with different genotypes (WT, OX, KO, and CO). (a) Root length of WT and GOX-related plants under salt stress; (b) Root length of of WT and DHAR3-related plants under salt stress; (c) Root length of WT and MDHAR-related plants under salt stress; Figure S2: Effects of salt stress on the phenotype and biomass. (a) Phenotype of WT and Trx-related plants under salt stress; (b) Phenotype of of WT and PrxR-related plants under salt stress; (c) Phenotype of WT and GOX-related plants under salt stress; (d,g) Biomass of WT and Trx-related plants under salt stress; (e,h) Biomass of of WT and PrxR-related plants under salt stress; (f,i) Biomass of WT and GOX-related plants under salt stress; Figure S3: MDA content of WT and GOX, PrxR, Trx, APX, DHAR3, and MDHAR-related plants under salt stress; Figure S4: AsA contents of WT and GOX, APX, DHAR3, and MDHAR-related plants under salt stress; Figure S5: (a-c) Analysis of the expression of antioxidative genes (BvM14-CAT, BvM14-APX and BvM14-PrxR) in WT and Trx-related plants under salt stress; (d-f) Antioxidant enzyme activity (BvM14-CAT, BvM14-APX, and BvM14-PrxR) in WT and Trx-related plants under salt stress; Figure S6: Analyses of relative gene expression and enzyme activities in DHAR3-related and MDHAR-related plants under salt stress. (a,c) Analysis of the expression of antioxidative genes (BvM14-APX and BvM14-MDHAR) in WT and DHAR3-related plants under salt stress; (b,d) Antioxidant enzyme activity (BvM14-APX and BvM14-MDHAR) in WT and DHAR3-related plants under salt stress; (e,f) Analysis of the expression of antioxidative genes (BvM14-MDHAR and BvM14-APX) in WT and MDHAR-related plants under salt stress; (g,h) Antioxidant enzyme activity (BvM14-MDHAR and BvM14-APX) in WT and MDHAR-related plants under salt stress; Figure S7: Identification of overexpression of BvM14-GOX, BvM14-PrxR, BvM14-Trx, BvM14-APX, BvM14-DHAR3, BvM14-MDHAR and atpox, atprxr, attrx, atapx, atdhar3, atmdhar mutant in Arabidopsis. (a) qRT-PCR analysis of the expression levels of the overexpressed BvM14-GOX (OX1 and OX2) in Arabidopsis; (b) qRT-PCR analysis of the expression levels of AtPOX in the atpox mutant; (c) qRT-PCR analysis of the expression levels of the overexpressed BvM14-PrxR (OX1 and OX2) in Arabidopsis; (d) qRT-PCR analysis of the expression levels of AtPrxR in the atprxr mutant; I qRT-PCR analysis of the expression levels of the overexpressed BvM14-Trx (OX1 and OX2) in Arabidopsis; (f) qRT-PCR analysis of the expression levels of AtTrx in the attrx mutant; (g) qRT-PCR analysis of the expression levels of the overexpressed BvM14-APX (OX1 and OX2) in Arabidopsis; (h) qRT-PCR analysis of the expression levels of AtAPX in the atapx mutant; (i) qRT-PCR analysis of the expression levels of the overexpressed BvM14- DHAR3 (OX1 and OX2) in Arabidopsis; (j) qRT-PCR analysis of the expression levels of AtDHAR3 in the atdhar3 mutant; (k) qRT-PCR analysis of the expression levels of the overexpressed BvM14-MDHAR (OX1 and OX2) in Arabidopsis; (l) qRT-PCR analysis of the expression levels of AtMDHAR in the atmdhar mutant.
Author Contributions
J.L.: conducted gene transcription analysis, conducted biochemical experiments, and wrote the first draft; S.C.: assisted with editing of the manuscript; C.M. and H.L. (Hongli Li): experimental design; D.J., J.N., M.X. and H.L. (He Liu): data analysis; B.Y., H.D. and H.L. (Haiying Li): funding acquisition, project supervision, and finalized the manuscript. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by the National Science Foundation of China (Project 32072122: The transcription factor BvGT-1 regulates the molecular mechanism of glyoxalase I gene involved in the salt tolerance of sugar beet M14); the National Science Foundation of China (Project 32272058: The molecular mechanism research of BvM14-STPK in regulation of its interaction proteins to salt tolerance).
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
The data are contained within the article and supplementary materials.
Acknowledgments
This work was supported by Heilongjiang Provincial Key Laboratory of Plant Genetic Engineering and Biological Fermentation Engineering for Cold Region.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Li, J.; Wang, K.; Ji, M.; Zhang, T.; Yang, C.; Liu, H.; Chen, S.; Li, H.; Li, H. Cys-SH based quantitative redox proteomics of salt induced response in sugar beet monosomic addition line M14. Bot Stud. 2021, 62, 16. [Google Scholar] [CrossRef] [PubMed]
- Liu, H.; Du, X.; Zhang, J.; Li, J.; Chen, S.; Duanmu, H.; Li, H. Quantitative redox proteomics revealed molecular mechanisms of salt tolerance in the roots of sugar beet monomeric addition line M14. Bot Stud. 2022, 63, 5. [Google Scholar] [CrossRef] [PubMed]
- Yang, L.; Ma, C.; Wang, L.; Chen, S.; Li, H. Salt stress induced proteome and transcriptome changes in sugar beet monosomic addition line M14. J. Plant. Physiol. 2012, 169, 839–850. [Google Scholar] [CrossRef] [PubMed]
- Yang, L.; Zhang, Y.; Zhu, N.; Koh, J.; Ma, C.; Pan, Y.; Yu, B.; Chen, S.; Li, H. Proteomic analysis of salt tolerance in sugar beet monosomic addition line M14. J. Proteome Res. 2013, 12, 4931–4950. [Google Scholar] [CrossRef]
- Mulet, J.M. Shaping the Sugar Beet of Tomorrow: Current Advances in Sugar Beet Biotechnology and New Breeding Techniques. In Sugar Beet Cultivation, Management and Processing; Springer: Berlin/Heidelberg, Germany, 2022; pp. 49–74. [Google Scholar]
- Chapman, J.M.; Muhlemann, J.K.; Gayomba, S.R.; Muday, G.K. RBOH-Dependent ROS Synthesis and ROS Scavenging by Plant Specialized Metabolites To Modulate Plant Development and Stress Responses. Chem. Res Toxicol. 2019, 32, 370–396. [Google Scholar] [CrossRef]
- Hasanuzzaman, M.; Bhuyan, M.; Zulfiqar, F.; Raza, A.; Mohsin, S.M.; Mahmud, J.A.; Fujita, M.; Fotopoulos, V. Reactive Oxygen Species and Antioxidant Defense in Plants under Abiotic Stress: Revisiting the Crucial Role of a Universal Defense Regulator. Antioxidants 2020, 9, 681. [Google Scholar] [CrossRef]
- Mhamdi, A.; Van Breusegem, F. Reactive oxygen species in plant development. Development 2018, 145, dev164376. [Google Scholar] [CrossRef]
- Qi, J.; Song, C.P.; Wang, B.; Zhou, J.; Kangasjärvi, J.; Zhu, J.K.; Gong, Z. Reactive oxygen species signaling and stomatal movement in plant responses to drought stress and pathogen attack. J. Integr. Plant Biol. 2018, 60, 805–826. [Google Scholar] [CrossRef]
- You, J.; Chan, Z. ROS Regulation During Abiotic Stress Responses in Crop Plants. Front Plant Sci. 2015, 6, 1092. [Google Scholar] [CrossRef]
- Zhao, C.; Zhang, H.; Song, C.; Zhu, J.K.; Shabala, S. Mechanisms of Plant Responses and Adaptation to Soil Salinity. Innovation 2020, 1, 100017. [Google Scholar] [CrossRef]
- Steffens, B. The role of ethylene and ROS in salinity, heavy metal, and flooding responses in rice. Front Plant Sci. 2014, 5, 685. [Google Scholar] [CrossRef]
- Tyagi, S.; Shah, A.; Karthik, K.; Rathinam, M.; Rai, V.; Chaudhary, N.; Sreevathsa, R. Reactive oxygen species in plants: An invincible fulcrum for biotic stress mitigation. Appl. Microbiol. Biotechnol. 2022, 106, 5945–5955. [Google Scholar] [CrossRef] [PubMed]
- Wang, M.; Gong, S.; Fu, L.; Hu, G.; Li, G.; Hu, S.; Yang, J. The Involvement of Antioxidant Enzyme System, Nitrogen Metabolism and Osmoregulatory Substances in Alleviating Salt Stress in Inbred Maize Lines and Hormone Regulation Mechanisms. Plants 2022, 11, 1547. [Google Scholar] [CrossRef] [PubMed]
- Yokochi, Y.; Fukushi, Y.; Wakabayashi, K.I.; Yoshida, K.; Hisabori, T. Oxidative regulation of chloroplast enzymes by thioredoxin and thioredoxin-like proteins in Arabidopsis thaliana. Proc. Natl. Acad. Sci USA 2021, 118, e2114952118. [Google Scholar] [CrossRef] [PubMed]
- Sainz, M.M.; Filippi, C.V.; Eastman, G.; Sotelo-Silveira, J.; Borsani, O.; Sotelo-Silveira, M. Analysis of Thioredoxins and Glutaredoxins in Soybean: Evidence of Translational Regulation under Water Restriction. Antioxidants 2022, 11, 1622. [Google Scholar] [CrossRef] [PubMed]
- Hossain, M.S.; ElSayed, A.I.; Moore, M.; Dietz, K.J. Redox and Reactive Oxygen Species Network in Acclimation for Salinity Tolerance in Sugar Beet. J. Exp. Bot. 2017, 68, 1283–1298. [Google Scholar] [CrossRef] [PubMed]
- Li, H.; Pan, Y.; Zhang, Y.; Wu, C.; Ma, C.; Yu, B.; Zhu, N.; Koh, J.; Chen, S. Salt stress response of membrane proteome of sugar beet monosomic addition line M14. J. Proteom. 2015, 127 (Pt A), 18–33. [Google Scholar] [CrossRef]
- Lv, X.; Jin, Y.; Wang, Y. De novo transcriptome assembly and identification of salt-responsive genes in sugar beet M14. Comput. Biol. Chem. 2018, 75, 1–10. [Google Scholar] [CrossRef]
- Zhang, Y.; Nan, J.; Yu, B. OMICS Technologies and Applications in Sugar Beet. Front Plant Sci. 2016, 7, 900. [Google Scholar] [CrossRef]
- Wang, Y.; Wang, S.; Tian, Y.; Wang, Q.; Chen, S.; Li, H.; Ma, C.; Li, H. Functional Characterization of a Sugar Beet BvbHLH93 Transcription Factor in Salt Stress Tolerance. Int. J. Mol. Sci. 2021, 22, 3669. [Google Scholar] [CrossRef]
- Yu, B.; Li, J.; Koh, J.; Dufresne, C.; Yang, N.; Qi, S.; Zhang, Y.; Ma, C.; Duong, B.V.; Chen, S.; et al. Quantitative proteomics and phosphoproteomics of sugar beet monosomic addition line M14 in response to salt stress. J. Proteomics. 2016, 143, 286–297. [Google Scholar] [CrossRef] [PubMed]
- Luan, J.; Dong, J.; Song, X.; Jiang, J.; Li, H. Overexpression of Tamarix hispida ThTrx5 Confers Salt Tolerance to Arabidopsis by Activating Stress Response Signals. Int. J. Mol. Sci. 2020, 21, 1165. [Google Scholar] [CrossRef] [PubMed]
- Lu, Z.; Liu, D.; Liu, S. Two rice cytosolic ascorbate peroxidases differentially improve salt tolerance in transgenic Arabidopsis. Plant Cell Rep. 2007, 26, 1909–1917. [Google Scholar] [CrossRef] [PubMed]
- Noshi, M.; Hatanaka, R.; Tanabe, N.; Terai, Y.; Maruta, T.; Shigeoka, S. Redox regulation of ascorbate and glutathione by a chloroplastic dehydroascorbate reductase is required for high-light stress tolerance in Arabidopsis. Biosci. Biotechnol. Biochem. 2016, 80, 870–877. [Google Scholar] [CrossRef]
- Qin, A.; Shi, Q.; Yu, X. Ascorbic acid contents in transgenic potato plants overexpressing two dehydroascorbate reductase genes. Mol. Biol. Rep. 2011, 38, 1557–1566. [Google Scholar] [CrossRef]
- Castro, B.; Citterico, M.; Kimura, S.; Stevens, D.M.; Wrzaczek, M.; Coaker, G. Stress-induced reactive oxygen species compartmentalization, perception and signalling. Nat. Plants. 2021, 7, 403–412. [Google Scholar] [CrossRef] [PubMed]
- Nadarajah, K.K. ROS Homeostasis in Abiotic Stress Tolerance in Plants. Int. J. Mol. Sci. 2020, 21, 5208. [Google Scholar] [CrossRef]
- Eltayeb, A.E.; Kawano, N.; Badawi, G.H.; Kaminaka, H.; Sanekata, T.; Shibahara, T.; Inanaga, S.; Tanaka, K. Overexpression of monodehydroascorbate reductase in transgenic tobacco confers enhanced tolerance to ozone, salt and polyethylene glycol stresses. Planta 2007, 225, 1255–1264. [Google Scholar] [CrossRef]
- Leterrier, M.; Corpas, F.J.; Barroso, J.B.; Sandalio, L.M.; del Río, L.A. Peroxisomal monodehydroascorbate reductase. Genomic clone characterization and functional analysis under environmental stress conditions. Plant Physiol. 2005, 138, 2111–2123. [Google Scholar]
- Rahantaniaina, M.S.; Li, S.; Chatel-Innocenti, G.; Tuzet, A.; Issakidis-Bourguet, E.; Mhamdi, A.; Noctor, G. Cytosolic and Chloroplastic DHARs Cooperate in Oxidative Stress-Driven Activation of the Salicylic Acid Pathway. Plant Physiol. 2017, 174, 956–971. [Google Scholar] [CrossRef]
- Sultana, S.; Khew, C.Y.; Morshed, M.M.; Namasivayam, P.; Napis, S.; Ho, C.L. Overexpression of monodehydroascorbate reductase from a mangrove plant (AeMDHAR) confers salt tolerance on rice. J. Plant Physiol. 2012, 169, 311–318. [Google Scholar] [CrossRef] [PubMed]
- Vanacker, H.; Guichard, M.; Bohrer, A.S.; Issakidis-Bourguet, E. Redox Regulation of Monodehydroascorbate Reductase by Thioredoxin y in Plastids Revealed in the Context of Water Stress. Antioxidants 2018, 7, 183. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Li, Z.; Peng, Y.; Wang, X.; Peng, D.; Li, Y.; He, X.; Zhang, X.; Ma, X.; Huang, L.; et al. Clones of FeSOD, MDHAR, DHAR Genes from White Clover and Gene Expression Analysis of ROS-Scavenging Enzymes during Abiotic Stress and Hormone Treatments. Molecules 2015, 20, 20939–20954. [Google Scholar] [CrossRef] [PubMed]
- Li, X.; Liao, M.; Huang, J.; Xu, Z.; Lin, Z.; Ye, N.; Zhang, Z.; Peng, X. Glycolate oxidase-dependent H(2)O(2) production regulates IAA biosynthesis in rice. BMC Plant Biol. 2021, 21, 326. [Google Scholar]
- Liu, Y.; Wu, W.; Chen, Z. Structures of glycolate oxidase from Nicotiana benthamiana reveal a conserved pH sensor affecting the binding of FMN. Biochem. Biophys. Res. Commun. 2018, 503, 3050–3056. [Google Scholar] [CrossRef]
- Schmitz, J.; Hüdig, M.; Meier, D.; Linka, N.; Maurino, V.G. The genome of Ricinus communis encodes a single glycolate oxidase with different functions in photosynthetic and heterotrophic organs. Planta 2020, 252, 100. [Google Scholar] [CrossRef]
- Zhang, Z.; Xu, Y.; Xie, Z.; Li, X.; He, Z.H.; Peng, X.X. Association-Dissociation of Glycolate Oxidase with Catalase in Rice: A Potential Switch to Modulate Intracellular H2O2 Levels. Mol. Plant. 2016, 9, 737–748. [Google Scholar] [CrossRef]
- Formentin, E.; Sudiro, C.; Ronci, M.B.; Locato, V.; Barizza, E.; Stevanato, P.; Ijaz, B.; Zottini, M.; De Gara, L.; Lo Schiavo, F. Signature and Innate Antioxidative Profile Make the Difference Between Sensitivity and Tolerance to Salt in Rice Cells. Front. Plant Sci. 2018, 9, 1549. [Google Scholar] [CrossRef]
- Liebthal, M.; Maynard, D.; Dietz, K.J. Peroxiredoxins and Redox Signaling in Plants. Antioxid. Redox. Signal. 2018, 28, 609–624. [Google Scholar] [CrossRef]
- Meyer, Y.; Belin, C.; Delorme-Hinoux, V.; Reichheld, J.P.; Riondet, C. Thioredoxin and glutaredoxin systems in plants: Molecular mechanisms, crosstalks, and functional significance. Antioxid. Redox. Signal. 2012, 17, 1124–1160. [Google Scholar] [CrossRef]
- Zhai, J.; Qi, Q.; Wang, M.; Yan, J.; Li, K.; Xu, H. Overexpression of tomato thioredoxin h (SlTrxh) enhances excess nitrate stress tolerance in transgenic tobacco interacting with SlPrx protein. Plant Sci. 2022, 315, 111137. [Google Scholar] [CrossRef] [PubMed]
- Ji, M.G.; Park, H.J.; Cha, J.Y.; Kim, J.A.; Shin, G.I.; Jeong, S.Y.; Lee, E.S.; Yun, D.J.; Lee, S.Y.; Kim, W.Y. Expression of Arabidopsis thaliana Thioredoxin-h2 in Brassica napus enhances antioxidant defenses and improves salt tolerance. Plant Physiol. Biochem. 2020, 147, 313–321. [Google Scholar] [CrossRef] [PubMed]
- Hasanuzzaman, M.; Bhuyan, M.; Anee, T.I.; Parvin, K.; Nahar, K.; Mahmud, J.A.; Fujita, M. Regulation of Ascorbate-Glutathione Pathway in Mitigating Oxidative Damage in Plants under Abiotic Stress. Antioxidants 2019, 8, 384. [Google Scholar] [CrossRef] [PubMed]
- Sofo, A.; Scopa, A.; Nuzzaci, M.; Vitti, A. Ascorbate Peroxidase and Catalase Activities and Their Genetic Regulation in Plants Subjected to Drought and Salinity Stresses. Int J Mol Sci. 2015, 16, 13561–13578. [Google Scholar] [CrossRef]
- Aleem, M.; Aleem, S.; Sharif, I.; Aleem, M.; Shahzad, R.; Khan, M.I.; Batool, A.; Sarwar, G.; Farooq, J.; Iqbal, A.; et al. Whole-Genome Identification of APX and CAT Gene Families in Cultivated and Wild Soybeans and Their Regulatory Function in Plant Development and Stress Response. Antioxidants 2022, 11, 1626. [Google Scholar] [CrossRef]
- Zhang, Z.; Zhang, Q.; Wu, J.; Zheng, X.; Zheng, S.; Sun, X.; Qiu, Q.; Lu, T. Gene knockout study reveals that cytosolic ascorbate peroxidase 2(OsAPX2) plays a critical role in growth and reproduction in rice under drought, salt and cold stresses. PLoS ONE 2013, 8, e57472. [Google Scholar] [CrossRef]
- Wang, Y.C.; Qu, G.Z.; Li, H.Y.; Wu, Y.J.; Wang, C.; Liu, G.F.; Yang, C.P. Enhanced salt tolerance of transgenic poplar plants expressing a manganese superoxide dismutase from Tamarix androssowii. Mol. Biol. Rep. 2010, 37, 1119–1124. [Google Scholar] [CrossRef]
- Chen, Y.; Jiang, J.; Chang, Q.; Gu, C.; Song, A.; Chen, S.; Dong, B.; Chen, F. Cold acclimation induces freezing tolerance via antioxidative enzymes, proline metabolism and gene expression changes in two chrysanthemum species. Mol. Biol. Rep. 2014, 41, 815–822. [Google Scholar] [CrossRef]
- Wang, D.R.; Yang, K.; Wang, X.; You, C.X. A C2H2-type zinc finger transcription factor, MdZAT17, acts as a positive regulator in response to salt stress. J. Plant Physiol. 2022, 275, 153737. [Google Scholar] [CrossRef]
- Zhang, H.; Mao, L.; Xin, M.; Xing, H.; Zhang, Y.; Wu, J.; Xu, D.; Wang, Y.; Shang, Y.; Wei, L.; et al. Overexpression of GhABF3 increases cotton(Gossypium hirsutum L.) tolerance to salt and drought. BMC Plant Biol. 2022, 22, 313. [Google Scholar]
- Duan, Y.B.; Li, J.; Qin, R.Y.; Xu, R.F.; Li, H.; Yang, Y.C.; Ma, H.; Li, L.; Wei, P.C.; Yang, J.B. Identification of a regulatory element responsible for salt induction of rice OsRAV2 through ex situ and in situ promoter analysis. Plant Mol. Biol. 2016, 90, 49–62. [Google Scholar] [CrossRef] [PubMed]
- Nawaz, I.; Iqbal, M.; Hakvoort, H.W.J.; de Boer, A.H.; Schat, H. Analysis of Arabidopsis thaliana HKT1 and Eutrema salsugineum/botschantzevii HKT1;2 Promoters in Response to Salt Stress in Athkt1:1 Mutant. Mol. Biotechnol. 2019, 61, 442–450. [Google Scholar] [CrossRef] [PubMed]
- Mizoi, J.; Shinozaki, K.; Yamaguchi-Shinozaki, K. AP2/ERF family transcription factors in plant abiotic stress responses. Biochim. Biophys. Acta 2012, 1819, 86–96. [Google Scholar] [CrossRef] [PubMed]
- Zhou, Y.; Wen, L.; Liao, L.; Lin, S.; Zheng, E.; Li, Y.; Zhang, Y. Comparative transcriptome analysis unveiling reactive oxygen species scavenging system of Sonneratia caseolaris under salinity stress. Front Plant Sci. 2022, 13, 953450. [Google Scholar] [CrossRef]
- Chen, Y.; Dai, Y.; Li, Y.; Yang, J.; Jiang, Y.; Liu, G.; Yu, C.; Zhong, F.; Lian, B.; Zhang, J. Overexpression of the Salix matsudana SmAP2-17 gene improves Arabidopsis salinity tolerance by enhancing the expression of SOS3 and ABI5. BMC Plant Biol. 2022, 22, 102. [Google Scholar]
- Yu, Z.; Yan, H.; Liang, L.; Zhang, Y.; Yang, H.; Li, W.; Choi, J.; Huang, J.; Deng, S. A C2H2-Type Zinc-Finger Protein from Millettia pinnata, MpZFP1, Enhances Salt Tolerance in Transgenic Arabidopsis. Int. J. Mol. Sci. 2021, 22, 10832. [Google Scholar] [CrossRef]








Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).