
Antioxidant Metabolism Underlies Different Metabolic Strategies for Primary Root Growth Maintenance under Water Stress in Cotton and Maize

 ,
, 
Abstract
1. Introduction
2. Materials and Methods
2.1. Plant Materials and Growth Conditions
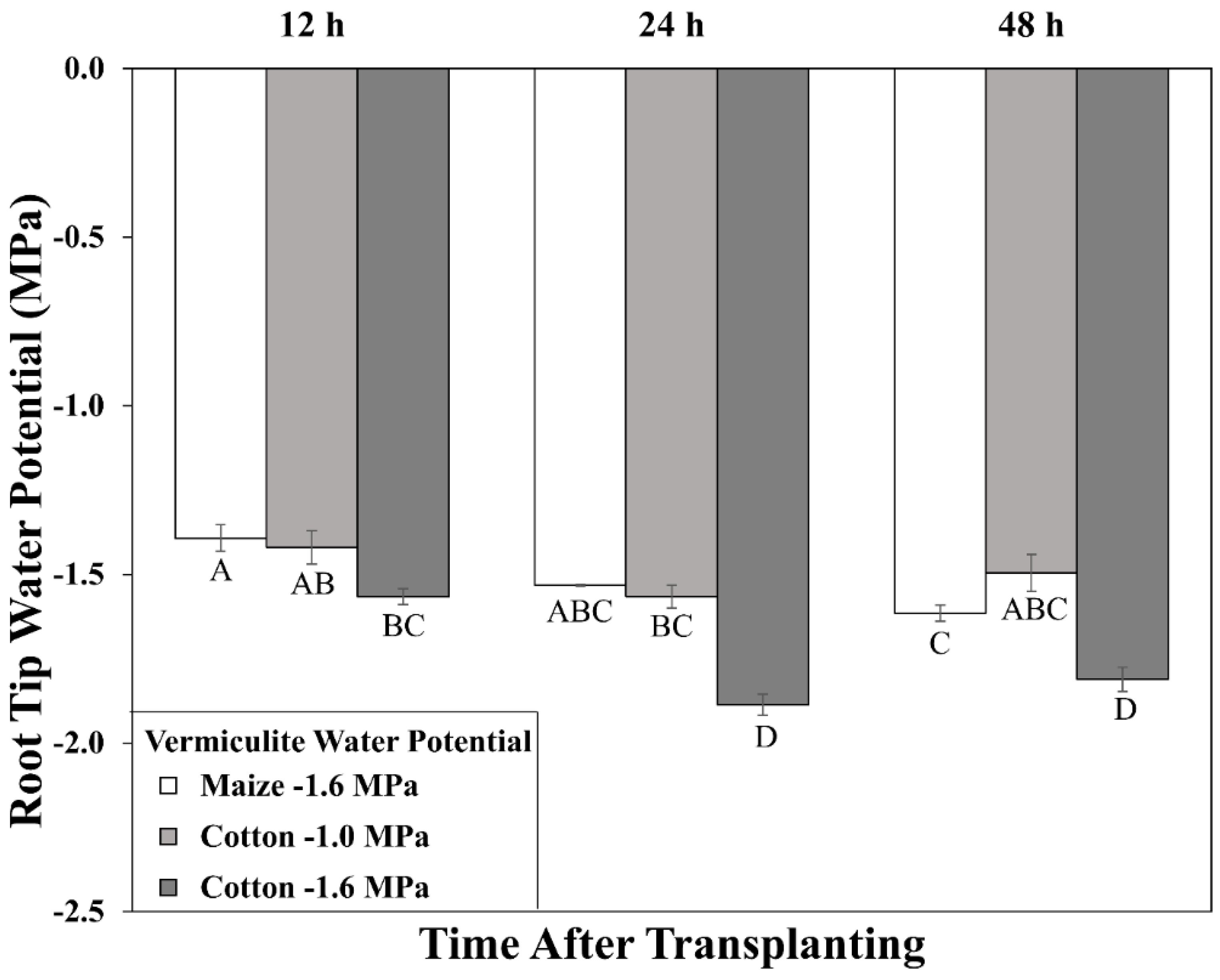
2.2. Root Elongation Zone Water Potentials
2.3. Kinematic Analysis
2.4. Metabolomics Analysis
2.5. Glutathione, Hydrogen Peroxide and Antioxidant Enzyme Assays
2.6. Statistical Analyses
3. Results
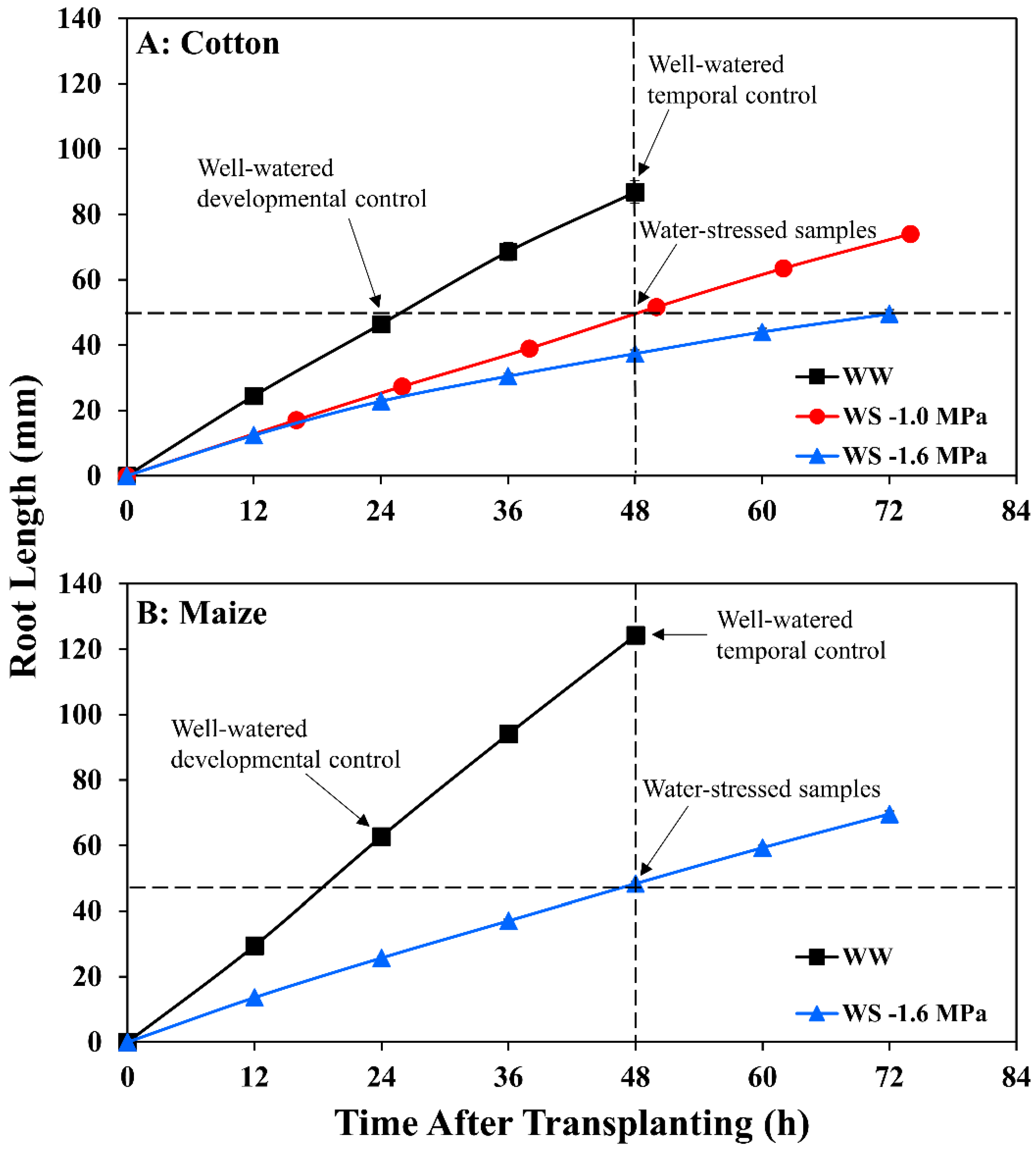
3.1. Cotton and Maize Primary Root Growth Responses to Equivalent Tissue Water Stress
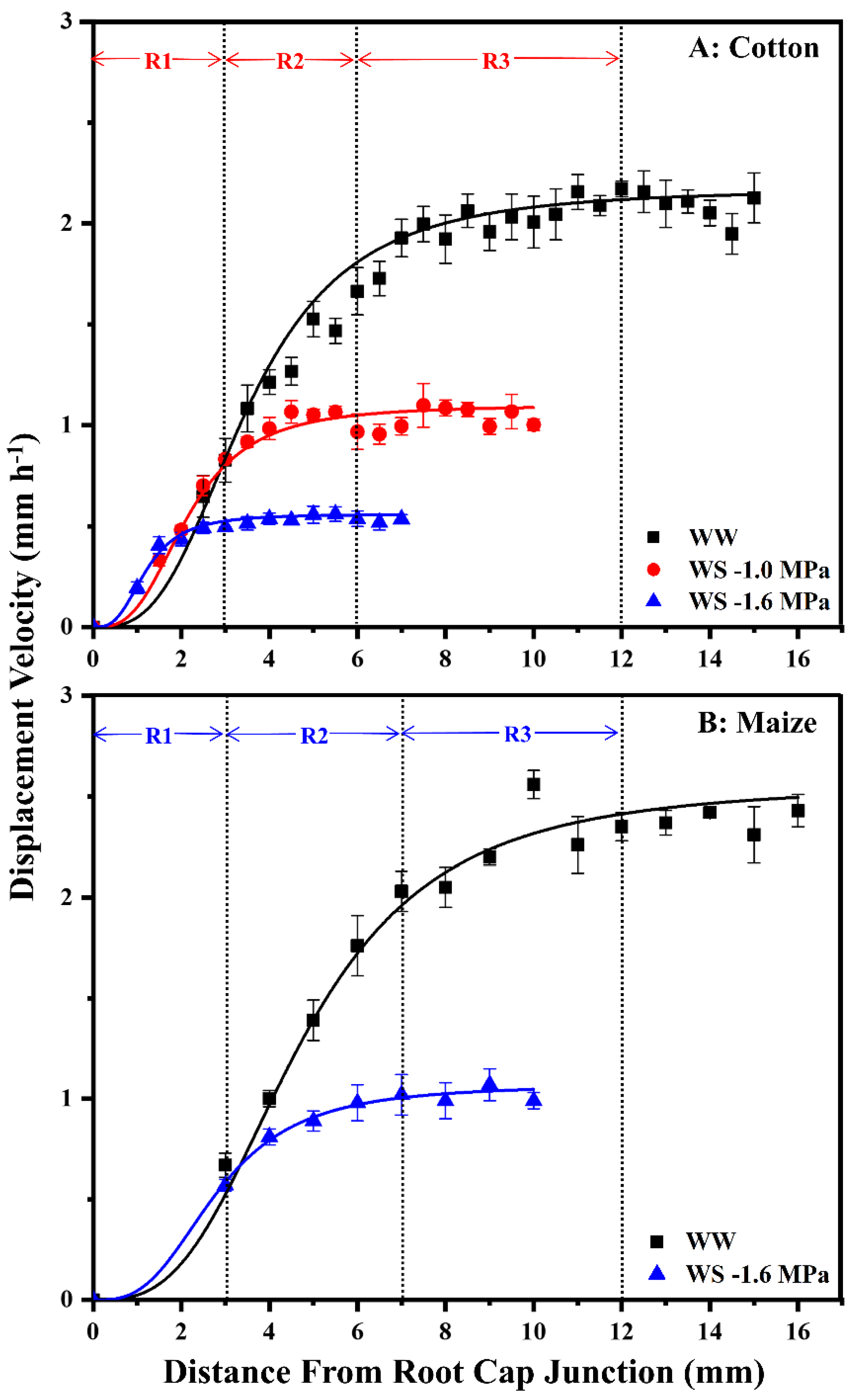
3.2. Kinematic Analysis of Cell Elongation Profiles and Cell Flux
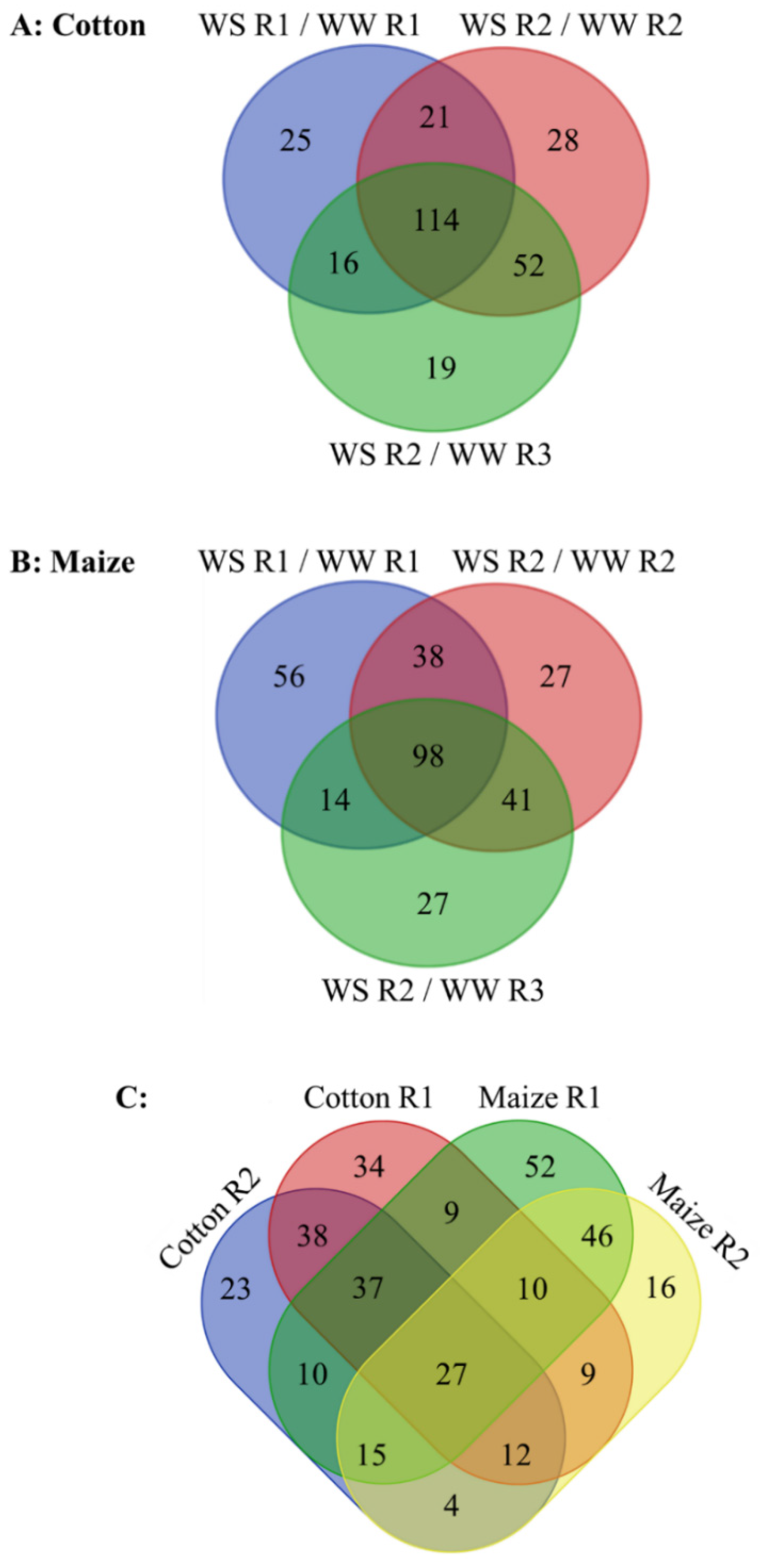
3.3. Overview of Comparative Metabolomics
3.4. Metabolic Responses to Water Stress in the Whole Elongation Zone
3.5. Metabolic Responses to Water Stress in R1 of the Elongation Zone
3.6. Metabolic Responses to Water Stress in R2 of the Elongation Zone
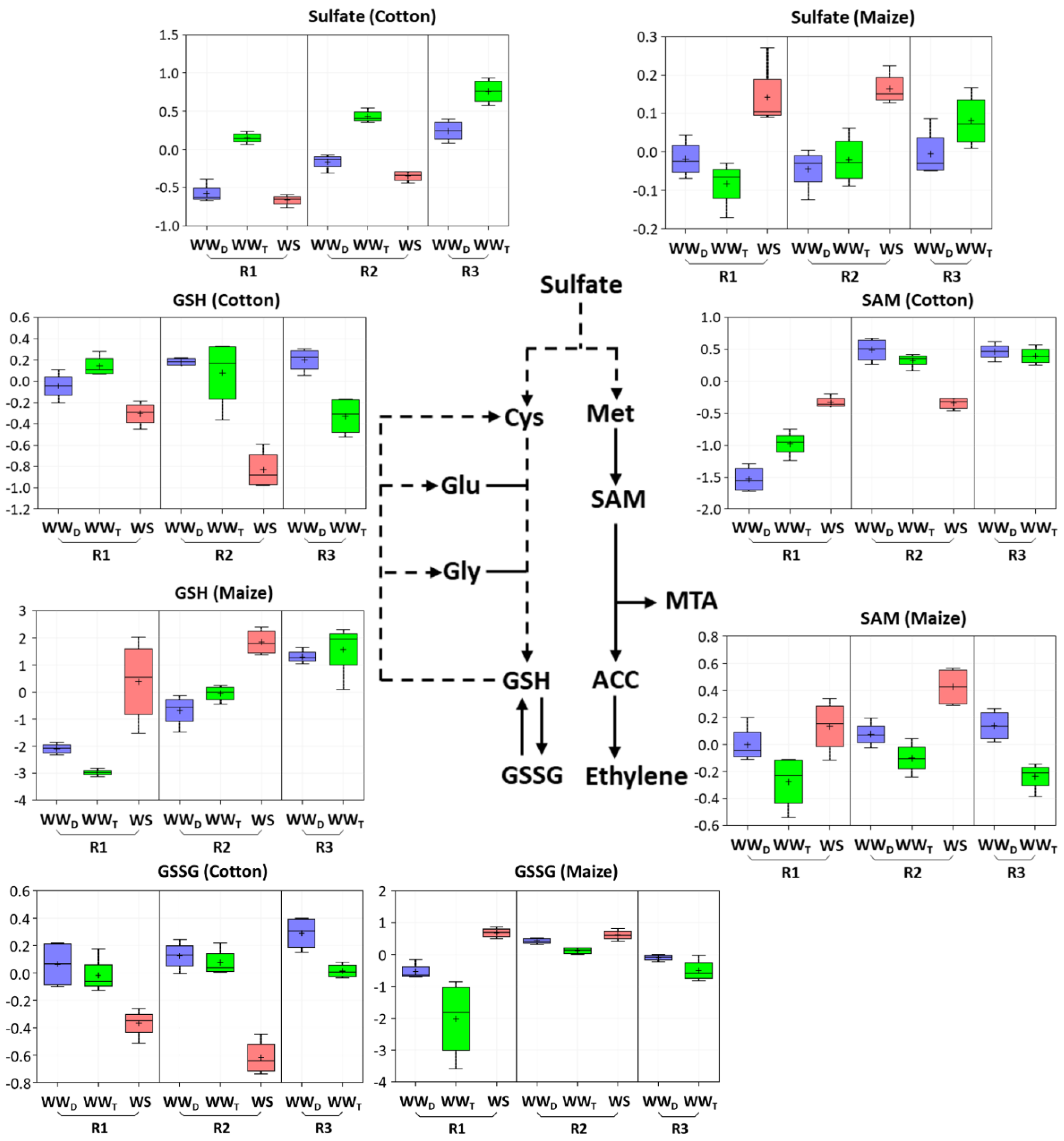
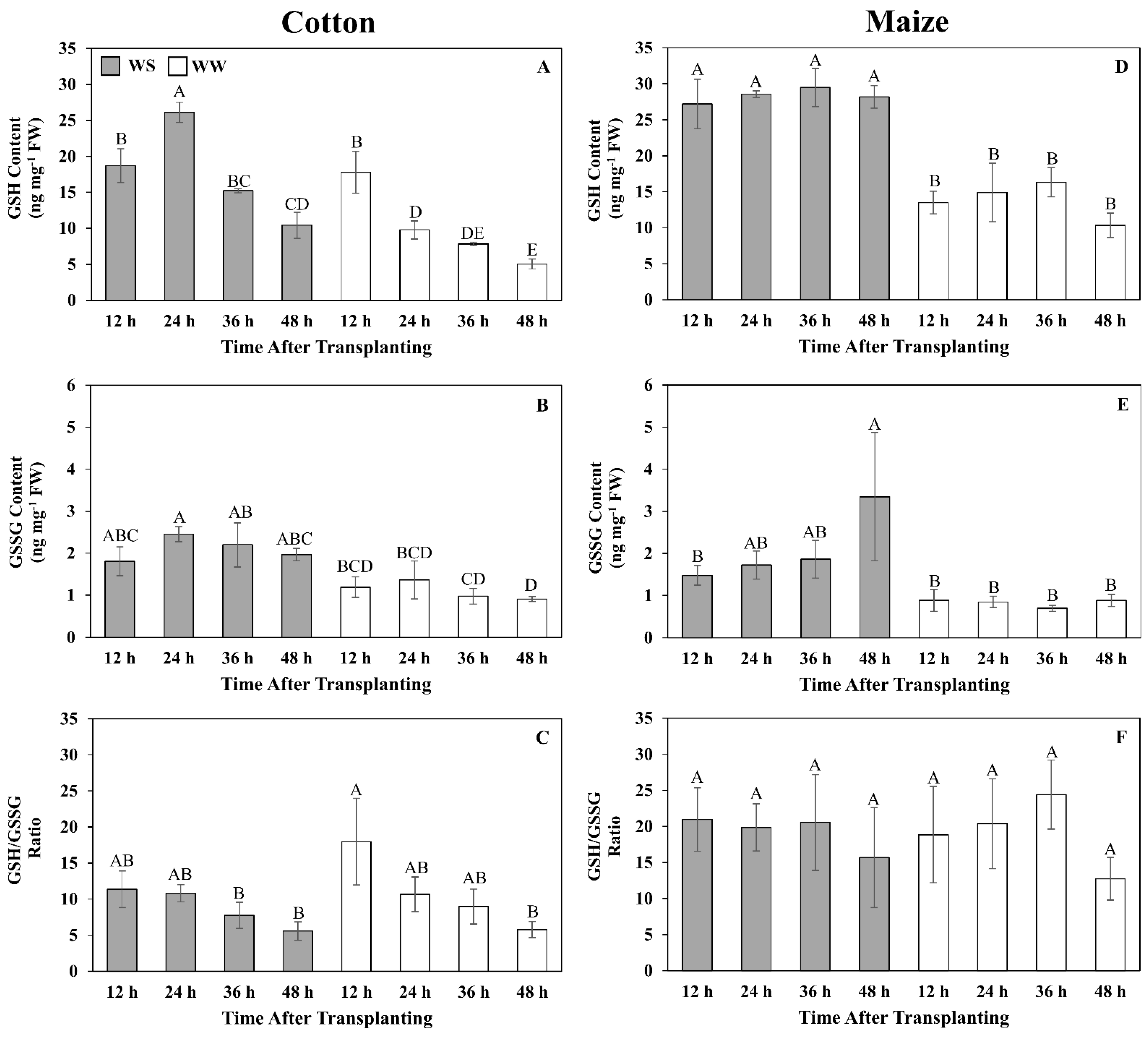
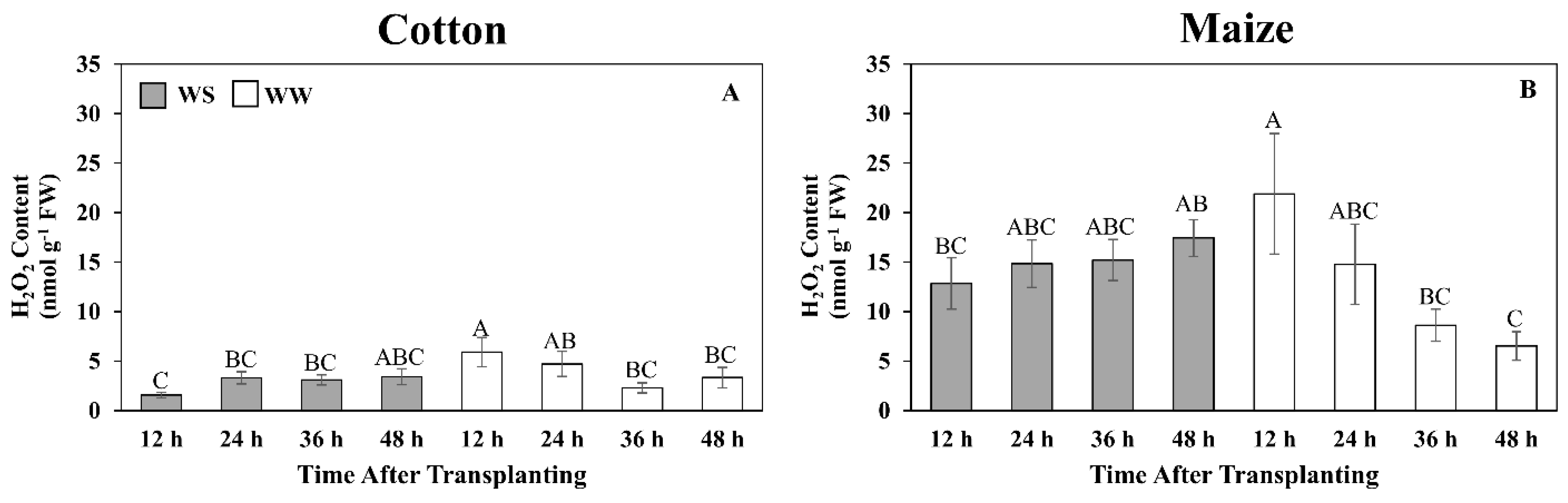
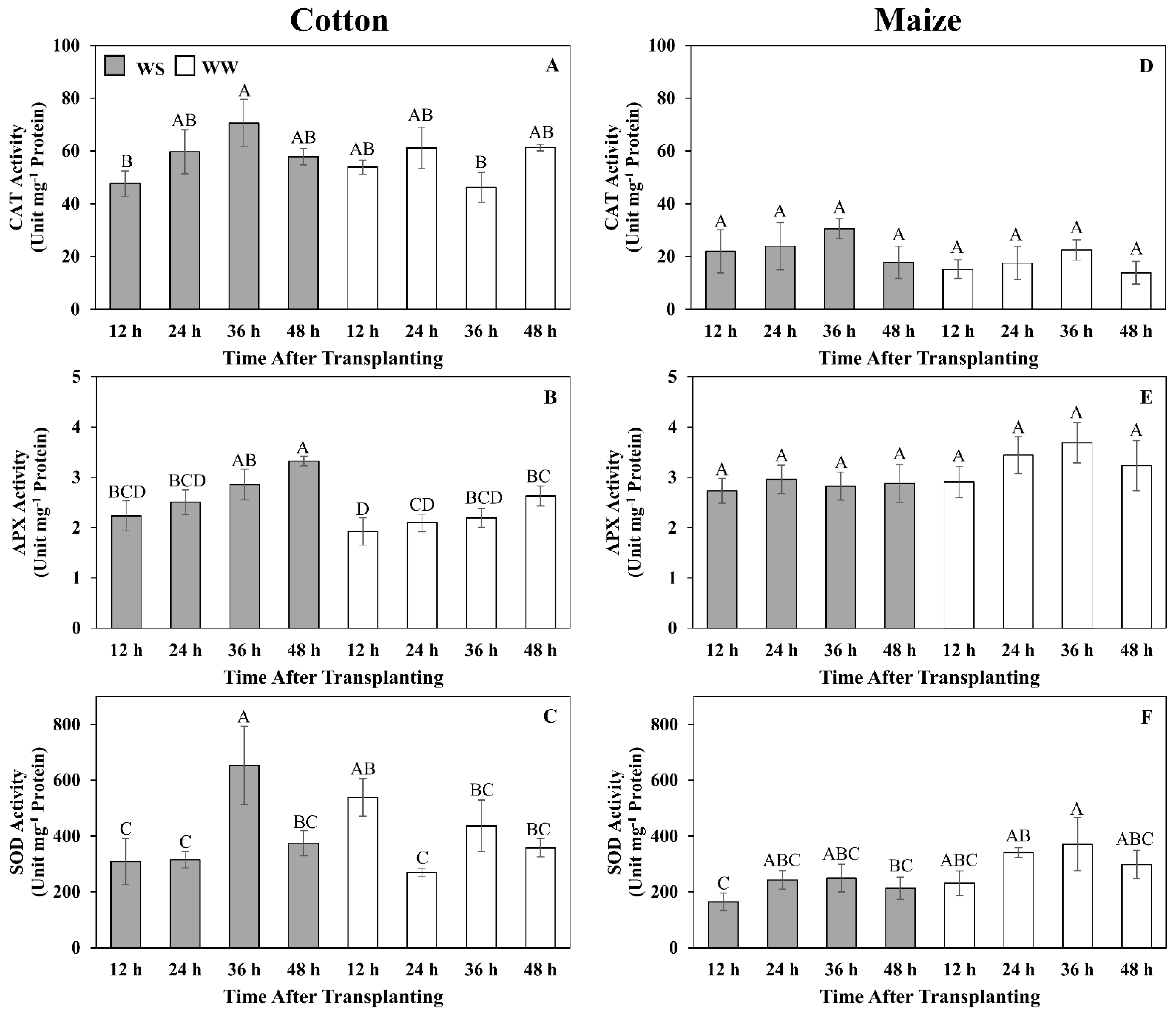
3.7. Quantitative Assessment of Glutathione Metabolism and Oxidative Stress
4. Discussion
4.1. Reducing Water Loss during Exposure to Low Water Potentials
4.2. Metabolic Responses to Water Stress Associated with Growth
4.3. Antioxidative Metabolism
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Zhang, H.; Zhao, Y.; Zhu, J.-K. Thriving under stress: How plants balance growth and the stress response. Dev. Cell 2020, 55, 529–543. [Google Scholar] [CrossRef] [PubMed]
- Ober, E.S.; Sharp, R.E. Maintaining root growth in drying soil: A review of progress and gaps in understanding. In Plant Roots: The Hidden Half, 4th ed.; Eshel, A., Beekman, T., Eds.; CRC Press: New York, NY, USA, 2013; Chapter 35; pp. 1–11. [Google Scholar]
- Sharp, R.E.; Silk, W.K.; Hsiao, T.C. Growth of the maize primary root at low water potentials. I. Spatial distribution of expansive growth. Plant Physiol. 1988, 87, 50–57. [Google Scholar] [CrossRef] [PubMed]
- Spollen, W.G.; Sharp, R.E.; Saab, I.N.; Wu, Y. Regulation of cell expansion in roots and shoots at low water potentials. In Water Deficits: Plant Responses from Cell to Community; Smith, J.A.C., Griffiths, H., Eds.; BIOS Scientific Publishers: Oxford, UK, 1993; pp. 37–52. [Google Scholar]
- van der Weele, C.M.; Spollen, W.G.; Sharp, R.E.; Baskin, T.I. Growth of Arabidopsis thaliana seedlings under water deficit by control of water potential in nutrient-agar media. J. Exp. Bot. 2000, 51, 1555–1562. [Google Scholar] [CrossRef] [PubMed]
- Yamaguchi, M.; Valliyodan, B.; Zhang, J.; LeNoble, M.E.; Yu, O.; Rogers, E.E.; Nguyen, H.T.; Sharp, R.E. Regulation of growth response to water stress in the soybean primary root. I. Proteomic analysis reveals region-specific regulation of phenylpropanoid metabolism and control of free iron in the elongation zone. Plant Cell Environ. 2010, 33, 223–243. [Google Scholar] [CrossRef] [PubMed]
- Yamaguchi, M.; Sharp, R.E. Complexity and coordination of root growth at low water potentials: Recent advances from transcriptomic and proteomic analyses. Plant Cell Environ. 2010, 33, 590–603. [Google Scholar] [CrossRef] [PubMed]
- Rosenow, D.T.; Quisenberry, J.E.; Wendt, C.W.; Clark, L.E. Drought tolerant sorghum and cotton germplasm. Agric. Water Manag. 1983, 7, 207–222. [Google Scholar] [CrossRef]
- Chapagain, A.K.; Hoekstra, A.Y.; Savenije, H.H.G.; Gautam, R. The water footprint of cotton consumption: An assessment of the impact of worldwide consumption of cotton products on the water resources in the cotton producing countries. Ecol. Econ. 2006, 60, 186–203. [Google Scholar] [CrossRef]
- Esmaeili, N.; Cai, Y.; Tang, F.; Zhu, X.; Smith, J.; Mishra, N.; Hequet, E.; Ritchie, G.; Jones, D.; Shen, G.; et al. Towards doubling fibre yield for cotton in the semiarid agricultural area by increasing tolerance to drought, heat and salinity simultaneously. Plant Biotechnol. J. 2021, 19, 462–476. [Google Scholar] [CrossRef]
- Liu, S.; Remley, M.; Bourland, F.M.; Nichols, R.L.; Stevens, W.E.; Phillips Jones, A.; Fritschi, F.B. Early vigor of advanced breeding lines and modern cotton cultivars. Crop Sci. 2015, 55, 1729–1740. [Google Scholar] [CrossRef]
- Virk, G.; Snider, J.L.; Pilon, C. Physiological contributors to early season whole-crop vigor in cotton. Crop Sci. 2019, 59, 2774–2783. [Google Scholar] [CrossRef]
- Klepper, B.; Taylor, H.M.; Huck, M.G.; Fiscus, E.L. Water relations and growth of cotton in drying soil. Agron. J. 1973, 65, 307–310. [Google Scholar] [CrossRef]
- McMichael, B.L.; Quisenberry, J.E. Genetic variation for root-shoot relationships among cotton germplasm. Environ. Exp. Bot. 1991, 31, 461–470. [Google Scholar] [CrossRef]
- Cook, C.G.; El-Zik, K.M. Cotton seedling and first-bloom plant characteristics: Relationships with drought-influenced boll abscission and lint yield. Crop Sci. 1992, 32, 1464–1467. [Google Scholar] [CrossRef]
- Basal, H.; Smith, C.W.; Thaxton, P.S.; Hemphill, J.K. Seedling drought tolerance in upland cotton. Crop Sci. 2005, 45, 766–771. [Google Scholar] [CrossRef]
- Taylor, H.M.; Ratliff, L.F. Root elongation rates of cotton and peanuts as a function of soil strength and soil water content. Soil Sci. 1969, 108, 113–119. [Google Scholar] [CrossRef]
- Pace, P.F.; Cralle, H.T.; El-Halawany, S.H.M.; Cothren, J.T.; Senseman, S.A. Drought-induced changes in shoot and root growth of young cotton plants. J. Cotton Sci. 1999, 3, 183–187. [Google Scholar]
- Bowne, J.B.; Erwin, T.A.; Juttner, J.; Schnurbusch, T.; Langridge, P.; Bacic, A.; Roessner, U. Drought responses of leaf tissues from wheat cultivars of differing drought tolerance at the metabolite level. Mol. Plant 2012, 5, 418–429. [Google Scholar] [CrossRef]
- Hochberg, U.; Degu, A.; Toubiana, D.; Gendler, T.; Nikoloski, Z.; Rachmilevitch, S.; Fait, A. Metabolite profiling and network analysis reveal coordinated changes in grapevine water stress response. BMC Plant Biol. 2013, 13, 184. [Google Scholar] [CrossRef]
- Boyer, J.S.; Knipling, E.B. Isopiestic technique for measuring leaf water potentials with a thermocouple psychrometer. Proc. Natl. Acad. Sci. USA 1965, 54, 1044–1051. [Google Scholar]
- Saab, I.N.; Sharp, R.E.; Pritchard, J.; Voetberg, G.S. Increased endogenous abscisic acid maintains primary root growth and inhibits shoot growth of maize seedlings at low water potentials. Plant Physiol. 1990, 93, 1329–1336. [Google Scholar] [CrossRef]
- Verslues, P.E.; Ober, E.S.; Sharp, R.E. Root growth and oxygen relations at low water potentials: Impact of oxygen availability in polyethylene glycol solutions. Plant Physiol. 1998, 116, 1403–1412. [Google Scholar] [CrossRef] [PubMed]
- Nonami, H.; Boyer, J.S. Turgor and growth at low water potentials. Plant Physiol. 1989, 89, 798–804. [Google Scholar] [CrossRef] [PubMed]
- Silk, W.K.; Lord, E.M.; Eckard, K.J. Growth patterns inferred from anatomical records: Empirical tests using longisections of roots of Zea mays L. Plant Physiol. 1989, 90, 708–713. [Google Scholar] [CrossRef] [PubMed]
- Sharp, R.E.; Poroyko, V.; Hejlek, L.G.; Spollen, W.G.; Springer, G.K.; Bohnert, H.J.; Nguyen, H.T. Root growth maintenance during water deficits: Physiology to functional genomics. J. Exp. Bot. 2004, 55, 2343–2351. [Google Scholar] [CrossRef] [PubMed]
- Erickson, R.O. Probability of division of cells in the epidermis of the Phleum root. Am. J. Bot. 1961, 48, 268–274. [Google Scholar] [CrossRef]
- Evans, A.M.; DeHaven, C.D.; Barrett, T.; Mitchell, M.; Milgram, E. Integrated, nontargeted ultrahigh performance liquid chromatography/electrospray ionization tandem mass spectrometry platform for the identification and relative quantification of the small-molecule complement of biological systems. Anal. Chem. 2009, 81, 6656–6667. [Google Scholar] [CrossRef]
- Yobi, A.; Wone, B.W.M.; Xu, W.; Alexander, D.C.; Guo, L.; Ryals, J.A.; Oliver, M.J.; Cushman, J.C. Metabolomic profiling in Selaginella lepidophylla at various hydration states provides new insights into the mechanistic basis of desiccation tolerance. Mol. Plant 2013, 6, 369–385. [Google Scholar] [CrossRef]
- Noctor, G.; Mhamdi, A.; Foyer, C.H. Oxidative stress and antioxidative systems: Recipes for successful data collection and interpretation. Plant Cell Environ. 2016, 39, 1140–1160. [Google Scholar] [CrossRef]
- Le, C.T.T.; Brumbarova, T.; Ivanov, R.; Stoof, C.; Weber, E.; Mohrbacher, J.; Fink-Straube, C.; Bauer, P. Zinc finger of arabidopsis thaliana12 (ZAT12) interacts with fer-like iron deficiency-induced transcription factor (FIT) linking iron deficiency and oxidative stress responses. Plant Physiol. 2016, 170, 540–557. [Google Scholar] [CrossRef]
- Bradford, M.M. A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein-dye binding. Anal. Biochem. 1976, 72, 248–254. [Google Scholar] [CrossRef]
- Aebi, H. Catalase in vitro. Meth. Enzymol. 1984, 105, 121–126. [Google Scholar]
- Amako, K.; Chen, G.-X.; Asada, K. Separate assays specific for ascorbate peroxidase and guaiacol peroxidase and for chloroplastic and cytosolic isozymes of ascorbate peroxidase in plants. Plant Cell Physiol. 1994, 35, 497–504. [Google Scholar]
- Giannopolitis, C.N.; Ries, S.K. Superoxide dismutases: I. Occurrence in higher plants. Plant Physiol. 1977, 59, 309–314. [Google Scholar] [CrossRef] [PubMed]
- Leach, K.A.; Hejlek, L.G.; Hearne, L.B.; Nguyen, H.T.; Sharp, R.E.; Davis, G.L. Primary root elongation rate and abscisic acid levels of maize in response to water stress. Crop Sci. 2011, 51, 157–172. [Google Scholar] [CrossRef]
- Spollen, W.G.; Tao, W.; Valliyodan, B.; Chen, K.; Hejlek, L.G.; Kim, J.J.; LeNoble, M.E.; Zhu, J.; Bohnert, H.J.; Henderson, D.; et al. Spatial distribution of transcript changes in the maize primary root elongation zone at low water potential. BMC Plant Biol. 2008, 8, 32. [Google Scholar] [CrossRef] [PubMed]
- Zhu, J.; Alvarez, S.; Marsh, E.L.; LeNoble, M.E.; Cho, I.J.; Sivaguru, M.; Chen, S.; Nguyen, H.T.; Wu, Y.; Schachtman, D.P.; et al. Cell wall proteome in the maize primary root elongation zone. II. Region-specific changes in water soluble and lightly ionically bound proteins under water deficit. Plant Physiol. 2007, 145, 1533–1548. [Google Scholar] [CrossRef] [PubMed]
- Voothuluru, P.; Anderson, J.C.; Sharp, R.E.; Peck, S.C. Plasma membrane proteomics in the maize primary root growth zone: Novel insights into root growth adaptation to water stress. Plant Cell Environ. 2016, 39, 2043–2054. [Google Scholar] [CrossRef]
- Voothuluru, P.; Sharp, R.E. Apoplastic hydrogen peroxide in the growth zone of the maize primary root under water stress. I. Increased levels are specific to the apical region of growth maintenance. J. Exp. Bot. 2013, 64, 1223–1233. [Google Scholar] [CrossRef]
- Seeve, C.M.; Cho, I.-J.; Hearne, L.B.; Srivastava, G.P.; Joshi, T.; Smith, D.O.; Sharp, R.E.; Oliver, M.J. Water deficit-induced changes in transcription factor expression in maize seedlings. Plant Cell Environ. 2017, 40, 686–701. [Google Scholar] [CrossRef]
- Delauney, A.J.; Verma, D.P.S. Proline biosynthesis and osmoregulation in plants. Plant J. 1993, 4, 215–223. [Google Scholar] [CrossRef]
- Sharp, R.E.; Davies, W.J. Solute regulation and growth by roots and shoots of water-stressed maize plants. Planta 1979, 147, 43–49. [Google Scholar] [CrossRef] [PubMed]
- Westgate, M.E.; Boyer, J.S. Osmotic adjustment and the inhibition of leaf, root, stem and silk growth at low water potentials in maize. Planta 1985, 164, 540–549. [Google Scholar] [CrossRef] [PubMed]
- Sharp, R.E.; Hsiao, T.C.; Silk, W.K. Growth of the maize primary root at low water potentials. II. Role of growth and deposition of hexose and potassium in osmotic adjustment. Plant Physiol. 1990, 93, 1337–1346. [Google Scholar] [CrossRef] [PubMed]
- Voetberg, G.S.; Sharp, R.E. Growth of the maize primary root at low water potentials. III. Role of increased proline deposition in osmotic adjustment. Plant Physiol. 1991, 96, 1125–1130. [Google Scholar] [CrossRef] [PubMed]
- Zhang, L.; Peng, J.; Chen, T.T.; Zhao, X.H.; Zhang, S.P.; Liu, S.D.; Dong, H.L.; Feng, L.; Yu, S.X. Effect of drought stress on lipid peroxidation and proline content in cotton roots. J. Anim. Plant Sci. 2014, 24, 1729–1736. [Google Scholar]
- Ahmad, N.; Malagoli, M.; Wirtz, M.; Hell, R. Drought stress in maize causes differential acclimation responses of glutathione and sulfur metabolism in leaves and roots. BMC Plant Biol. 2016, 16, 247. [Google Scholar] [CrossRef]
- Ober, E.S.; Sharp, R.E. Proline accumulation in maize (Zea mays L.) primary roots at low water potentials. I. Requirement for increased levels of abscisic acid. Plant Physiol. 1994, 105, 981–987. [Google Scholar] [CrossRef]
- Ashraf, M.; Foolad, M.R. Roles of glycine betaine and proline in improving plant abiotic stress resistance. Environ. Exp. Bot. 2007, 59, 206–216. [Google Scholar] [CrossRef]
- Lum, M.S.; Hanafi, M.M.; Rafii, Y.M.; Akmar, A.S.N. Effect of drought stress on growth, proline and antioxidant enzyme activities of upland rice. J. Anim. Plant Sci. 2014, 24, 1487–1493. [Google Scholar]
- Moulin, M.; Deleu, C.; Larher, F.; Bouchereau, A. The lysine-ketoglutarate reductase–saccharopine dehydrogenase is involved in the osmo-induced synthesis of pipecolic acid in rapeseed leaf tissues. Plant Physiol. Biochem. 2006, 44, 474–482. [Google Scholar] [CrossRef]
- Nishizawa, A.; Yabuta, Y.; Shigeoka, S. Galactinol and raffinose constitute a novel function to protect plants from oxidative damage. Plant Physiol. 2008, 147, 1251–1263. [Google Scholar] [CrossRef] [PubMed]
- Van den Ende, W. Multifunctional fructans and raffinose family oligosaccharides. Front. Plant Sci. 2013, 4, 247. [Google Scholar] [PubMed]
- van der Rest, B.; Boisson, A.M.; Gout, E.; Bligny, R.; Douce, R. Glycerophosphocholine metabolism in higher plant cells. Evidence of a new glyceryl-phosphodiester phosphodiesterase. Plant Physiol. 2002, 130, 244–255. [Google Scholar] [CrossRef]
- Larsson, K.E.; Nyström, B.; Liljenberg, C. A phosphatidylserine decarboxylase activity in root cells of oat (Avena sativa) is involved in altering membrane phospholipid composition during drought stress acclimation. Plant Physiol. Biochem. 2006, 44, 211–219. [Google Scholar] [CrossRef] [PubMed]
- Cheng, L.; Bucciarelli, B.; Liu, J.; Zinn, K.; Miller, S.; Patton-Vogt, J.; Allan, D.; Shen, J.; Vance, C.P. White lupin cluster root acclimation to phosphorus deficiency and root hair development involve unique glycerophosphodiester phosphodiesterases. Plant Physiol. 2011, 156, 1131–1148. [Google Scholar] [CrossRef]
- Du, Y.; Zhao, Q.; Chen, L.; Yao, X.; Zhang, W.; Zhang, B.; Xie, F. Effect of drought stress on sugar metabolism in leaves and roots of soybean seedlings. Plant Physiol. Biochem. 2020, 146, 1–12. [Google Scholar] [CrossRef]
- Verslues, P.E.; Sharma, S. Proline metabolism and its implications for plant-environment interaction. Arab. Book 2010, 8, e0140. [Google Scholar] [CrossRef]
- Kopriva, S.; Malagoli, M.; Takahashi, H. Sulfur nutrition: Impacts on plant development, metabolism, and stress responses. J. Exp. Bot. 2019, 70, 4069–4073. [Google Scholar] [CrossRef]
- Morgan, P.W.; Drew, M.C. Ethylene and plant responses to stress. Physiol. Plant. 1997, 100, 620–630. [Google Scholar] [CrossRef]
- Sobeih, W.Y.; Dodd, I.C.; Bacon, M.A.; Grierson, D.; Davies, W.J. Long-distance signals regulating stomatal conductance and leaf growth in tomato (Lycopersicon esculentum) plants subjected to partial root-zone drying. J. Exp. Bot. 2004, 55, 2353–2363. [Google Scholar] [CrossRef]
- Haworth, I.S.; Rodger, A.; Richards, W.G. A molecular mechanics study of spermine complexation to DNA: A new model for spermine-poly (dG-dC) binding. Proc. R. Soc. Lond. Ser. B Biol. Sci. 1991, 244, 107–116. [Google Scholar]
- Chan, K.X.; Wirtz, M.; Phua, S.Y.; Estavillo, G.M.; Pogson, B.J. Balancing metabolites in drought: The sulfur assimilation conundrum. Trends Plant Sci. 2013, 18, 18–29. [Google Scholar] [CrossRef] [PubMed]
- Sweetlove, L.J.; Beard, K.F.M.; Nunes-Nesi, A.; Fernie, A.R.; Ratcliffe, R.G. Not just a circle: Flux modes in the plant TCA cycle. Trends Plant Sci. 2010, 15, 462–470. [Google Scholar] [CrossRef] [PubMed]
- Bouché, N.; Fromm, H. GABA in plants: Just a metabolite? Trends Plant Sci. 2004, 9, 110–115. [Google Scholar] [CrossRef] [PubMed]
- Liu, C.; Zhao, L.; Yu, G. The dominant glutamic acid metabolic flux to produce γ-amino butyric acid over proline in Nicotiana tabacum leaves under water stress relates to its significant role in antioxidant activity. J. Integr. Plant Biol. 2011, 53, 608–618. [Google Scholar] [CrossRef]
- Gakière, B.; Hao, J.; de Bont, L.; Pétriacq, P.; Nunes-Nesi, A.; Fernie, A.R. NAD+ biosynthesis and signaling in plants. Crit. Rev. Plant Sci. 2018, 37, 259–307. [Google Scholar] [CrossRef]
- Wei, M.; Zhuang, Y.; Li, H.; Li, P.; Huo, H.; Shu, D.; Huang, W.; Wang, S. The cloning and characterization of hypersensitive to salt stress mutant, affected in quinolinate synthase, highlights the involvement of NAD in stress-induced accumulation of ABA and proline. Plant J. 2020, 102, 85–98. [Google Scholar] [CrossRef]
- Noctor, G.; Foyer, C.H. Ascorbate and glutathione: Keeping active oxygen under control. Annu. Rev. Plant Physiol. Plant Mol. Biol. 1998, 49, 249–279. [Google Scholar] [CrossRef]
- Noctor, G.; Foyer, C.H. Intracellular redox compartmentation and ROS-related communication in regulation and signaling. Plant Physiol. 2016, 171, 1581–1592. [Google Scholar] [CrossRef]
- Nayyar, H.; Gupta, D. Differential sensitivity of C3 and C4 plants to water deficit stress: Association with oxidative stress and antioxidants. Environ. Exp. Bot. 2006, 58, 106–113. [Google Scholar] [CrossRef]
- Tausz, M.; Šircelj, H.; Grill, D. The glutathione system as a stress marker in plant ecophysiology: Is a stress-response concept valid? J. Exp. Bot. 2004, 55, 1955–1962. [Google Scholar] [CrossRef] [PubMed]
- Foyer, C.H.; Noctor, G. Ascorbate and glutathione: The heart of the redox hub. Plant Physiol. 2011, 155, 2–18. [Google Scholar] [CrossRef] [PubMed]
- Vernoux, T.; Wilson, R.C.; Seeley, K.A.; Reichheld, J.P.; Muroy, S.; Brown, S.; Maughan, S.C.; Cobbett, C.S.; Van Montagu, M.; Inzé, D.; et al. The root meristemless1/cadmium sensitive2 gene defines a glutathione-dependent pathway involved in initiation and maintenance of cell division during postembryonic root development. Plant Cell 2000, 12, 97–110. [Google Scholar] [CrossRef] [PubMed]
- Diaz Vivancos, P.; Dong, Y.; Ziegler, K.; Markovic, J.; Pallardó, F.V.; Pellny, T.K.; Verrier, P.J.; Foyer, C.H. Recruitment of glutathione into the nucleus during cell proliferation adjusts whole-cell redox homeostasis in Arabidopsis thaliana and lowers the oxidative defence shield. Plant J. 2010, 64, 825–838. [Google Scholar] [CrossRef]
- Voothuluru, P.; Mäkelä, P.; Zhu, J.; Yamaguchi, M.; Cho, I.-J.; Oliver, M.J.; Simmonds, J.; Sharp, R.E. Apoplastic hydrogen peroxide in the growth zone of the maize primary root. Increased levels differentially modulate root elongation under well-watered and water-stressed conditions. Front. Plant Sci. 2020, 11, 392. [Google Scholar] [CrossRef]
- Lappartient, A.G.; Touraine, B. Glutathione-mediated regulation of ATP sulfurylase activity, SO42− uptake, and oxidative stress response in intact canola roots. Plant Physiol. 1997, 114, 177–183. [Google Scholar] [CrossRef][Green Version]
- Munné-Bosch, S. The role of α-tocopherol in plant stress tolerance. J. Plant Physiol. 2005, 162, 743–748. [Google Scholar] [CrossRef]
- Zhang, H.; Ni, Z.; Chen, Q.; Guo, Z.; Gao, W.; Su, X.; Qu, Y. Proteomic responses of drought-tolerant and drought-sensitive cotton varieties to drought stress. Mol. Genet. Genom. 2016, 291, 1293–1303. [Google Scholar] [CrossRef]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
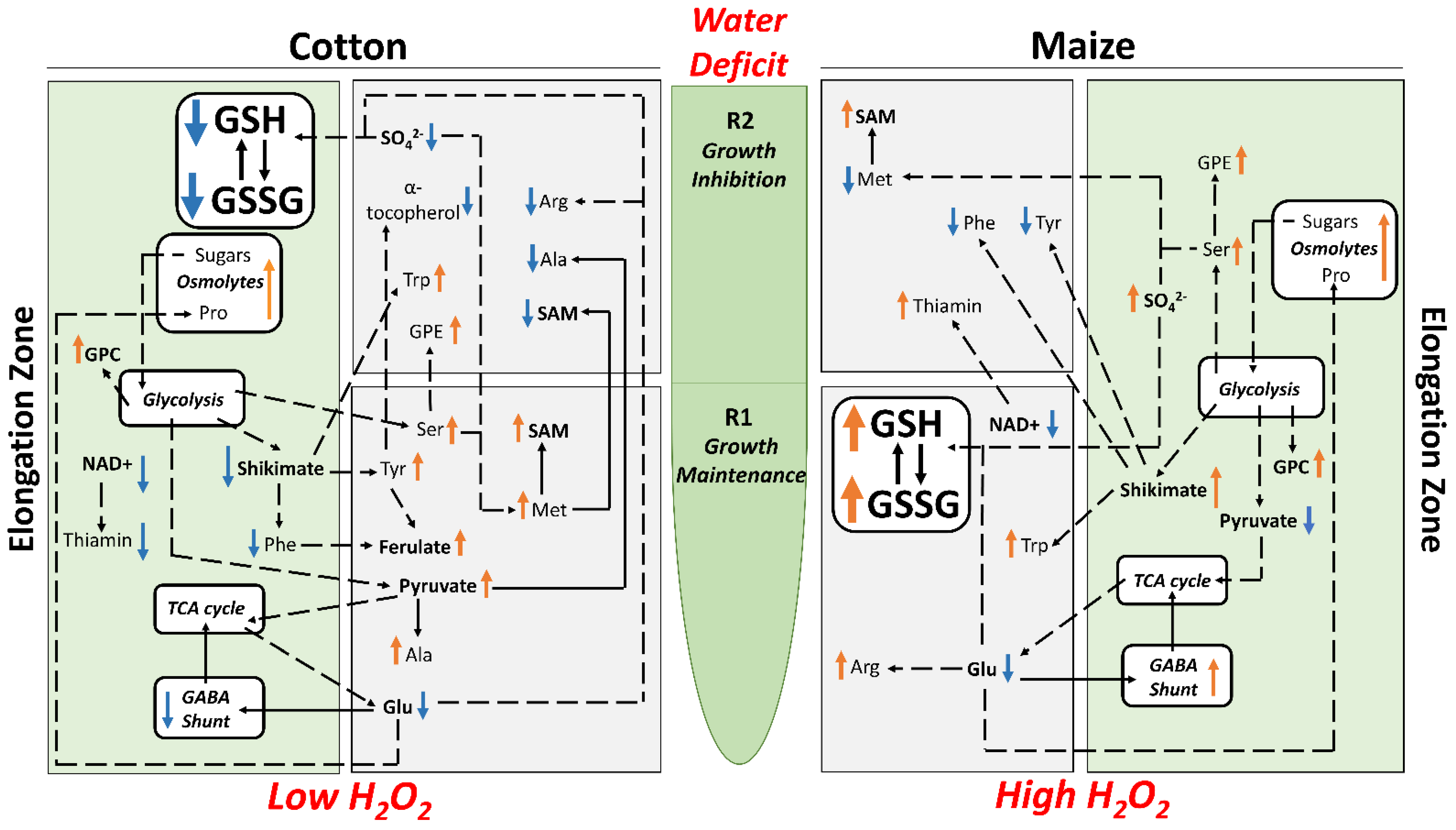{kind=link}
| Super Pathway | Metabolite | R1 | R2 | ||
|---|---|---|---|---|---|
| Cotton | Maize | Cotton | Maize | ||
| Carbohydrate | Raffinose | 26.76 | 10.42 | 71.6 | 6.97 |
| Sucrose | 4.62 | 11.95 | 17.33 | 4.88 | |
| Fructose | 3.26 | 2.61 | 1.81 | 2.3 | |
| Glucose | 2.9 | 1.77 | 1.71 | 1.35 | |
| Myo-inositol | 2.59 | 1.45 | 2.11 | 1.75 | |
| Lipids | GPC | 2.27 | 338.42 | 3.74 | 342.28 |
| Phosphoethanolamine | 3.66 | 6.49 | 1.74 | 2.67 | |
| Amino acid | Saccharopine | 8.9 | 19.51 | 4.13 | 16.39 |
| Pipecolate | 8.76 | 3.63 | 5.05 | 4.02 | |
| Proline | 5.62 | 16.34 | 5.04 | 4.63 | |
| N-methylproline | 2.4 | 2.99 | 2.37 | 3.6 | |
| Taurine | 3.67 | 3 | 3.97 | 1.95 | |
| β-hydroxyisovalerate | 5.05 | 0.49 | 3.43 | 0.46 | |
| N6-acetyllysine | 3.75 | 0.34 | 2.28 | 0.52 | |
| 2-aminoadipate | 2.49 | 0.11 | 1.9 | 0.28 | |
| S-carboxymethyl-L-cysteine | 0.64 | 0.26 | 0.62 | 0.4 | |
| 5-oxoproline | 0.64 | 0.53 | 0.39 | 0.37 | |
| Glutamine | 0.54 | 0.52 | 0.2 | 0.36 | |
| Histidine | 0.53 | 0.61 | 0.48 | 0.65 | |
| N-formylphenylalanine | 0.39 | 0.09 | 0.4 | 0.11 | |
| GABA | 0.69 | 2.79 | 0.54 | 3.97 | |
| Shikimate | 0.51 | 1.95 | 0.33 | 4.89 | |
| S-methylglutathione | 0.49 | 2.45 | 0.63 | 1.99 | |
| Others | Threonate | 0.77 | 0.57 | 0.73 | 0.69 |
| Riboflavin | 0.69 | 0.5 | 0.45 | 0.67 | |
| Adenosine | 0.55 | 0.61 | 0.45 | 0.68 | |
| Xanthosine | 0.27 | 0.35 | 0.39 | 0.28 | |
| Super Pathway | Metabolite | R1 | R2 | ||
|---|---|---|---|---|---|
| Cotton | Maize | Cotton | Maize | ||
| Carbohydrate | Pyruvate | 1.94 | 0.53 | NS | 0.6 |
| Malate | NS | 0.67 | 0.5 | NS | |
| Amino acid | N-acetylproline | 4.65 | 1.75 | 5.04 | NS |
| Serine | 1.76 | 2.58 | NS | 1.82 | |
| Carboxyethyl-GABA | 1.92 | 0.41 | 1.64 | NS | |
| Tyrosine | 1.35 | NS | NS | 0.5 | |
| Arginine | NS | 1.47 | 0.47 | NS | |
| Tryptophan | NS | 1.55 | 1.58 | NS | |
| Glutamate | 0.72 | 0.71 | NS | NS | |
| Quinate | 0.5 | NS | 0.51 | 2.16 | |
| Phenylalanine | 0.7 | NS | 0.42 | 0.65 | |
| Lipids | GPE | NS | 31.92 | 1.87 | 12.27 |
| 1-palmitoyl-GPG (16:0) | 0.62 | NS | 0.38 | 2.92 | |
| Others | 2’-AMP | 1.6 | NS | NS | 0.53 |
| IMP | 2.32 | 0.6 | 0.36 | NS | |
| Ferulate | 1.76 | NS | NS | NS | |
| Allantoin | NS | 3.04 | 0.59 | 1.52 | |
| FAD | 0.51 | 0.35 | 0.49 | NS | |
| Thiamin | 0.51 | NS | 0.56 | 2.29 | |
| α-tocopherol | NS | NS | 0.51 | NS | |
| Metabolite | Cotton | Maize | ||
|---|---|---|---|---|
| R1 | R2 | R1 | R2 | |
| 5-oxoproline | 0.64 | 0.39 | 0.53 | 0.37 |
| Glutathione, oxidized (GSSG) | 0.68 | 0.48 | 6.4 | NS |
| Glutathione, reduced (GSH) | 0.71 | 0.43 | 41.38 | NS |
| γ-glutamylleucine | 2.5 | 1.43 | NS | NS |
| γ-glutamylthreonine | NS | NS | 1.87 | 2.75 |
| Ophthalmate | 4.42 | NS | NS | NS |
| Norophthalmate | 1.64 | NS | ||
| Cys-gly, oxidized | 2.54 | NS | 4.85 | 7.07 |
| Sulfate | NS | 0.54 | 1.22 | 1.18 |
| Cysteine | NS | NS | 1.7 | NS |
| Methionine | 3.08 | NS | NS | 0.31 |
| S-adenosylmethionine (SAM) | 2.59 | 0.47 | NS | 1.61 |
| 5-methylthioadenosine (MTA) | 1.56 | 0.6 | NS | NS |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kang, J.; Voothuluru, P.; Hoyos-Miernyk, E.; Alexander, D.; Oliver, M.J.; Sharp, R.E. Antioxidant Metabolism Underlies Different Metabolic Strategies for Primary Root Growth Maintenance under Water Stress in Cotton and Maize. Antioxidants 2022, 11, 820. https://doi.org/10.3390/antiox11050820
Kang J, Voothuluru P, Hoyos-Miernyk E, Alexander D, Oliver MJ, Sharp RE. Antioxidant Metabolism Underlies Different Metabolic Strategies for Primary Root Growth Maintenance under Water Stress in Cotton and Maize. Antioxidants. 2022; 11(5):820. https://doi.org/10.3390/antiox11050820
Chicago/Turabian StyleKang, Jian, Priyamvada Voothuluru, Elizabeth Hoyos-Miernyk, Danny Alexander, Melvin J. Oliver, and Robert E. Sharp. 2022. "Antioxidant Metabolism Underlies Different Metabolic Strategies for Primary Root Growth Maintenance under Water Stress in Cotton and Maize" Antioxidants 11, no. 5: 820. https://doi.org/10.3390/antiox11050820
APA StyleKang, J., Voothuluru, P., Hoyos-Miernyk, E., Alexander, D., Oliver, M. J., & Sharp, R. E. (2022). Antioxidant Metabolism Underlies Different Metabolic Strategies for Primary Root Growth Maintenance under Water Stress in Cotton and Maize. Antioxidants, 11(5), 820. https://doi.org/10.3390/antiox11050820