
Teneligliptin Exerts Antinociceptive Effects in Rat Model of Partial Sciatic Nerve Transection Induced Neuropathic Pain





{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Reagents
2.2. Animals
2.3. Establishment of NP
2.4. Intrathecal Catheterization and Osmotic Pump Infusion
2.5. Behavior Test for Tactile Allodynia
2.6. Behavior Test for Thermal Hyperalgesia
2.7. Tail Flick Assay
2.8. Spinal Cord Sample Preparation and Western Blotting Analysis
2.9. Immunfluoroscence Studies
2.10. Experimental Protocol
2.11. Statistical Analysis
3. Results
3.1. TEN Has Mild Analgesic Effects against Acute Pain
3.2. PSNT Induces Tactile Allodynia and Thermal Hyperalgesia and TEN Alleviated PSNT Induced Thermal Hyperalgesia and Mechanical Allodynia
3.3. Intrathecal Naloxone Pre-Treatment Antagonized MOR Analgesic Effect but Not TEN Induced Analgesia on Both Acute Pain and NP
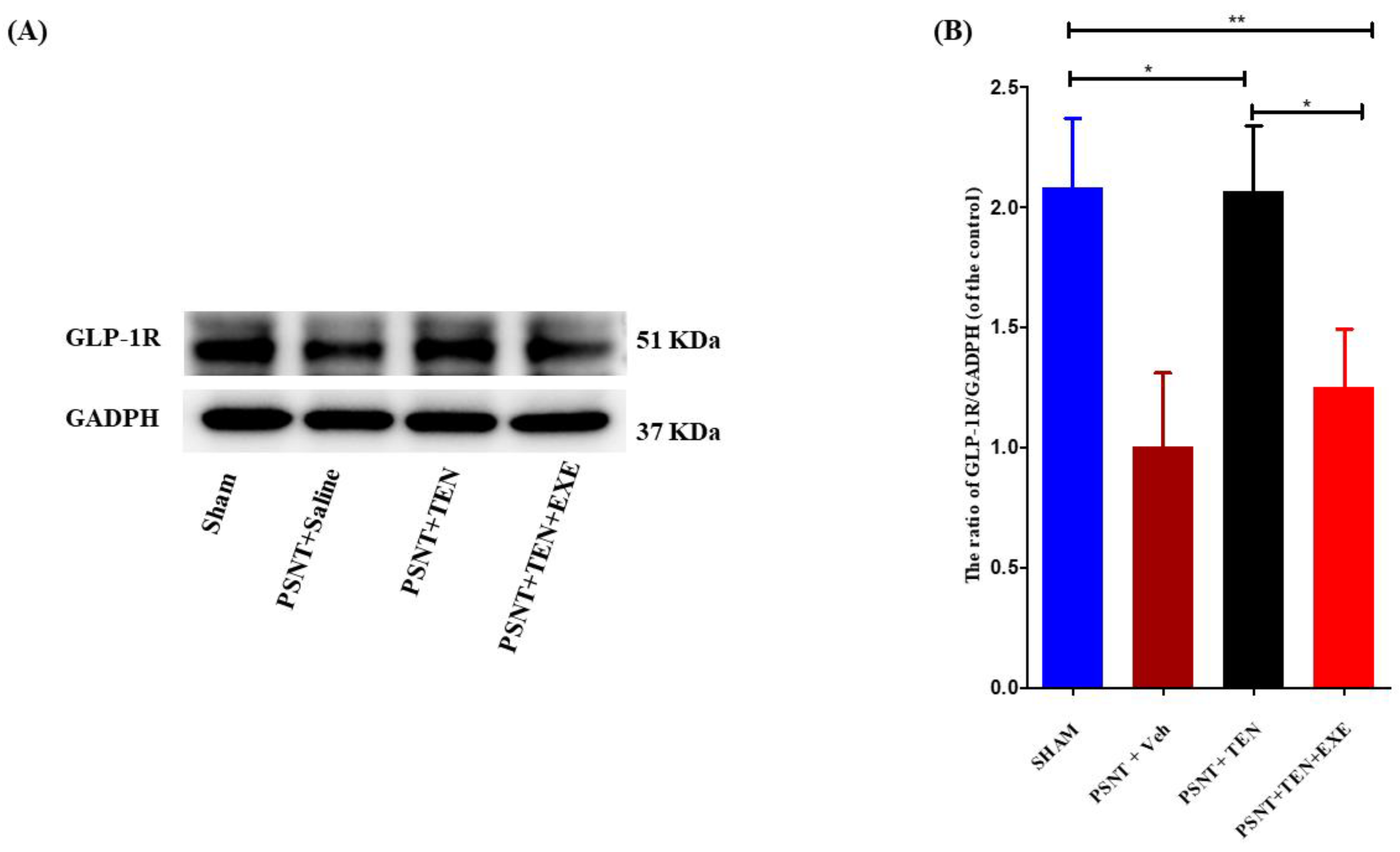
3.4. TEN Reverses the Decrease in Spinal GLP-1 Expression in the PSNT Rats
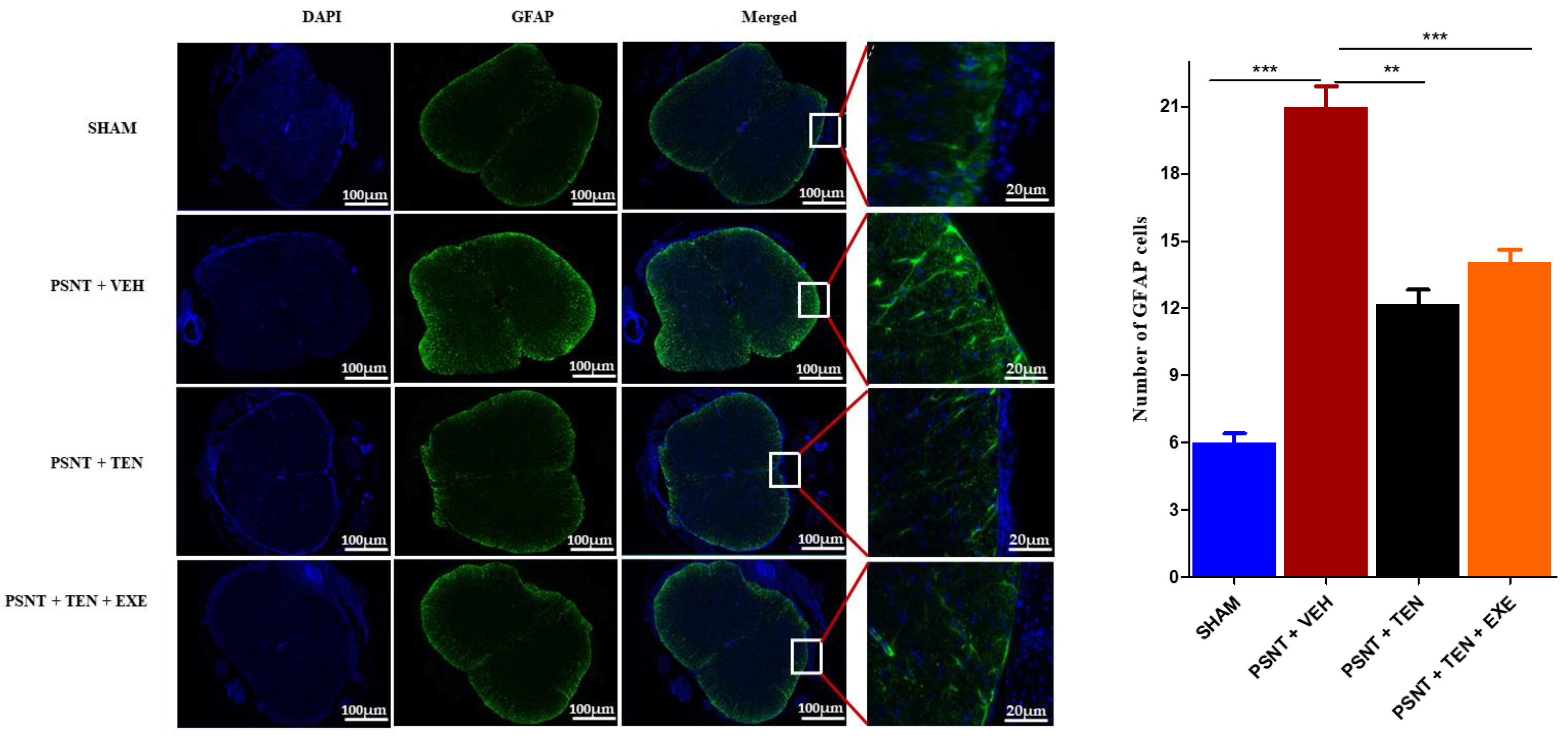
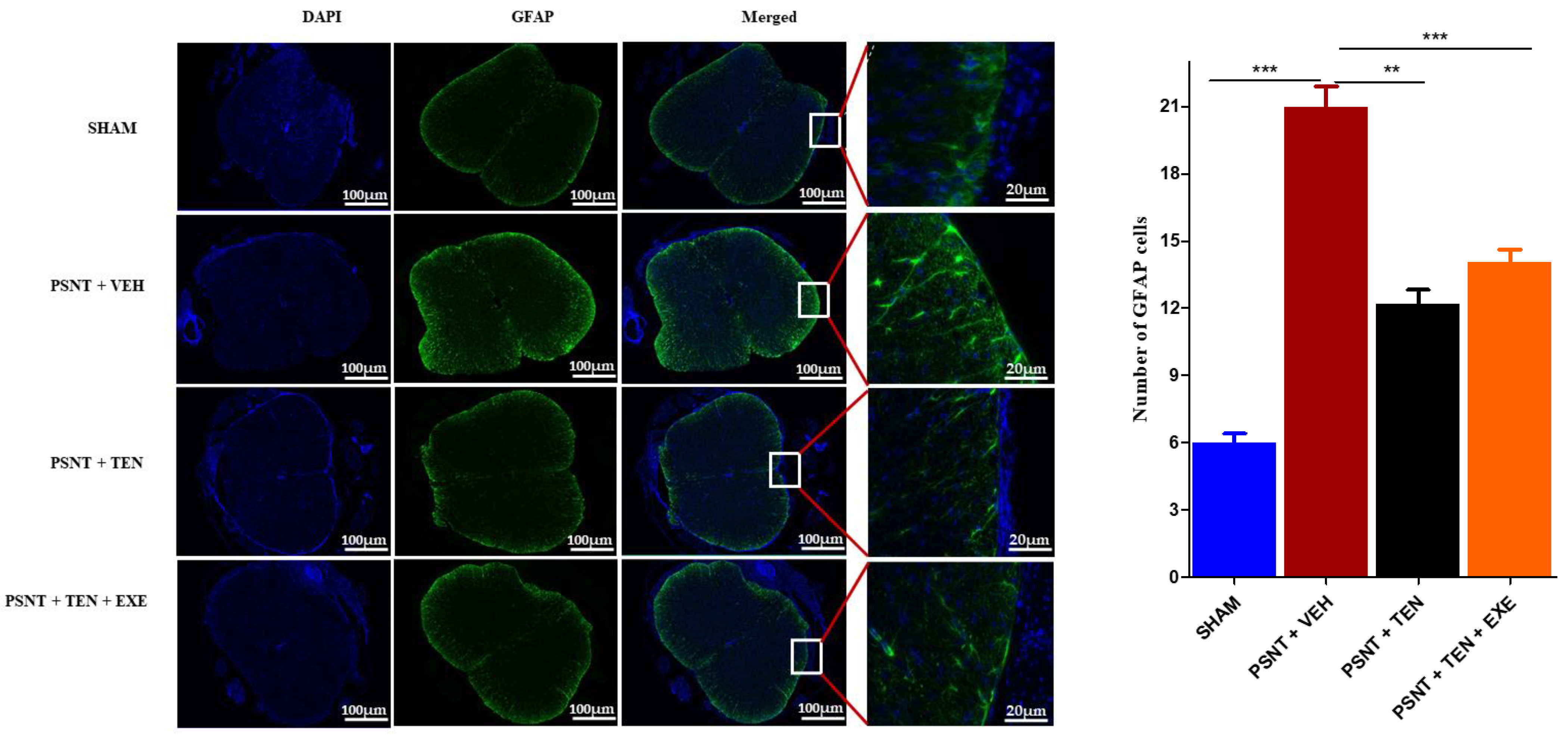
3.5. TEN Inhibits Spinal GFAP Activation in PSNT Rats
4. Discussions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Murnion, B.P. Neuropathic pain: Current definition and review of drug treatment. Aust. Prescr. 2018, 41, 60–63. [Google Scholar] [CrossRef] [PubMed]
- Cavalli, E.; Mammana, S.; Nicoletti, F.; Bramanti, P.; Mazzon, E. The neuropathic pain: An overview of the current treatment and future therapeutic approaches. Int. J. Immunopathol. Pharmacol. 2019, 33, 2058738419838383. [Google Scholar] [CrossRef] [Green Version]
- Eisenberg, E.; McNicol, E.D.; Carr, D.B. Efficacy and safety of opioid agonists in the treatment of neuropathic pain of nonmalignant origin: Systematic review and meta-analysis of randomized controlled trials. JAMA 2005, 293, 3043–3052. [Google Scholar] [CrossRef]
- Cooper, T.E.; Chen, J.; Wiffen, P.J.; Derry, S.; Carr, D.B.; Aldington, D.; Cole, P.; Moore, R.A. Morphine for chronic neuropathic pain in adults. Cochrane Database Syst. Rev. 2017, 5, CD011669. [Google Scholar] [CrossRef] [PubMed]
- Sommer, C.; Klose, P.; Welsch, P.; Petzke, F.; Häuser, W. Opioids for chronic non-cancer neuropathic pain. An updated systematic review and meta-analysis of efficacy, tolerability and safety in randomized placebo-controlled studies of at least 4 weeks duration. Eur. J. Pain 2020, 24, 3–18. [Google Scholar] [CrossRef] [PubMed]
- Sun, L.; Zhao, J.Y.; Gu, X.; Liang, L.; Wu, S.; Mo, K.; Feng, J.; Guo, W.; Zhang, J.; Bekker, A.; et al. Nerve injury-induced epigenetic silencing of opioid receptors controlled by DNMT3a in primary afferent neurons. Pain 2017, 158, 1153–1165. [Google Scholar] [CrossRef] [PubMed]
- Schembri, E. Are Opioids Effective in Relieving Neuropathic Pain? SN Compr. Clin. Med. 2019, 1, 30–46. [Google Scholar] [CrossRef] [Green Version]
- Liu, L.; Liu, J.; Tian, X.Y.; Wong, W.T.; Lau, C.W.; Xu, A.; Xu, G.; Ng, C.F.; Yao, X.; Gao, Y.; et al. Uncoupling protein-2 mediates DPP-4 inhibitor-induced restoration of endothelial function in hypertension through reducing oxidative stress. Antioxid. Redox Signal. 2014, 21, 1571–1581. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Civantos, E.; Bosch, E.; Ramirez, E.; Zhenyukh, O.; Egido, J.; Lorenzo, O.; Mas, S. Sitagliptin ameliorates oxidative stress in experimental diabetic nephropathy by diminishing the miR-200a/Keap-1/Nrf2 antioxidant pathway. Diabetes Metab. Syndr. Obes. 2017, 10, 207–222. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Abdelsalam, R.M.; Safar, M.M. Neuroprotective effects of vildagliptin in rat rotenone Parkinson’s disease model: Role of RAGE-NFκB and Nrf2-antioxidant signaling pathways. J. Neurochem. 2015, 133, 700–707. [Google Scholar] [CrossRef]
- Sharma, S.; Sharma, R.; Hatware, K.; Patil, K. Review on Chemistry, Analysis and Pharmacology of Teneligliptin: A Novel DPP-4 Inhibitor. Mini Rev. Med. Chem. 2020, 20, 1091–1100. [Google Scholar] [CrossRef] [PubMed]
- Yoshida, T.; Akahoshi, F.; Sakashita, H.; Kitajima, H.; Nakamura, M.; Sonda, S.; Takeuchi, M.; Tanaka, Y.; Ueda, N.; Sekiguchi, S.; et al. Discovery and preclinical profile of teneligliptin (3-[(2S,4S)-4-[4-(3-methyl-1-phenyl-1H-pyrazol-5-yl)piperazin-1-yl]pyrrolidin-2-ylcarbonyl]thiazolidine): A highly potent, selective, long-lasting and orally active dipeptidyl peptidase IV inhibitor for the treatment of type 2 diabetes. Bioorg. Med. Chem. 2012, 20, 5705–5719. [Google Scholar] [CrossRef] [PubMed]
- Kadowaki, T.; Haneda, M.; Ito, H.; Sasaki, K.; Matsukawa, M.; Yamada, Y. Long-Term, Real-World Safety and Efficacy of Teneligliptin: A Post-Marketing Surveillance of More Than 10,000 Patients with Type 2 Diabetes in Japan. Adv. Ther. 2020, 37, 1065–1086. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Butul, M.; Pingali, U.; Nutalapati, C. Evaluation of the analgesic activity of single and multiple oral doses of teneligliptin (20 mg/day), using hot air analgesiometer in healthy human volunteers: A randomized, double blind, placebo controlled, cross over study. Int. J. Basic Clin. Pharmacol. 2020, 9, 498–505. [Google Scholar] [CrossRef]
- Nakagami, H.; Pang, Z.; Shimosato, T.; Moritani, T.; Kurinami, H.; Koriyama, H.; Tenma, A.; Shimamura, M.; Morishita, R. The dipeptidyl peptidase-4 inhibitor teneligliptin improved endothelial dysfunction and insulin resistance in the SHR/NDmcr-cp rat model of metabolic syndrome. Hypertens. Res. 2014, 37, 629–635. [Google Scholar] [CrossRef]
- Zhang, Z.; Jin, X.; Yang, C.; Li, Y. Teneligliptin protects against hypoxia/reoxygenation-induced endothelial cell injury. Biomed. Pharm. 2019, 109, 468–474. [Google Scholar] [CrossRef] [PubMed]
- Kimura, S.; Inoguchi, T.; Yamasaki, T.; Yamato, M.; Ide, M.; Sonoda, N.; Yamada, K.; Takayanagi, R. A novel DPP-4 inhibitor teneligliptin scavenges hydroxyl radicals: In vitro study evaluated by electron spin resonance spectroscopy and in vivo study using DPP-4 deficient rats. Metabolism 2016, 65, 138–145. [Google Scholar] [CrossRef]
- De Nigris, V.; Prattichizzo, F.; Mancuso, E.; Spiga, R.; Pujadas, G.; Ceriello, A. Teneligliptin enhances the beneficial effects of GLP-1 in endothelial cells exposed to hyperglycemic conditions. Oncotarget 2017, 9, 8898–8910. [Google Scholar] [CrossRef] [Green Version]
- Gong, N.; Xiao, Q.; Zhu, B.; Zhang, C.Y.; Wang, Y.C.; Fan, H.; Ma, A.N.; Wang, Y.X. Activation of spinal glucagon-like peptide-1 receptors specifically suppresses pain hypersensitivity. J. Neurosci. 2014, 34, 5322–5334. [Google Scholar] [CrossRef] [Green Version]
- Lee, C.H.; Jeon, S.J.; Cho, K.S.; Moon, E.; Sapkota, A.; Jun, H.S.; Ryu, J.H.; Choi, J.W. Activation of Glucagon-Like Peptide-1 Receptor Promotes Neuroprotection in Experimental Autoimmune Encephalomyelitis by Reducing Neuroinflammatory Responses. Mol. Neurobiol. 2018, 55, 3007–3020. [Google Scholar] [CrossRef] [PubMed]
- Jing, F.; Zou, Q.; Wang, Y.; Cai, Z.; Tang, Y. Activation of microglial GLP-1R in the trigeminal nucleus caudalis suppresses central sensitization of chronic migraine after recurrent nitroglycerin stimulation. J. Headache Pain 2021, 22, 86. [Google Scholar] [CrossRef] [PubMed]
- Xu, M.; Wu, H.Y.; Liu, H.; Gong, N.; Wang, Y.R.; Wang, Y.X. Morroniside, a secoiridoid glycoside from Cornus officinalis, attenuates neuropathic pain by activation of spinal glucagon-like peptide-1 receptors. Br. J. Pharmacol. 2017, 174, 580–590. [Google Scholar] [CrossRef] [Green Version]
- Zhu, B.; Gong, N.; Fan, H.; Peng, C.S.; Ding, X.J.; Jiang, Y.; Wang, Y.X. Lamiophlomis rotata, an orally available Tibetan herbal painkiller, specifically reduces pain hypersensitivity states through the activation of spinal glucagon-like peptide-1 receptors. Anesthesiology 2014, 121, 835–851. [Google Scholar] [CrossRef] [Green Version]
- Zhang, L.Q.; Zhang, W.; Li, T.; Yang, T.; Yuan, X.; Zhou, Y.; Zou, Q.; Yang, H.; Gao, F.; Tian, Y.; et al. GLP-1R activation ameliorated novel-object recognition memory dysfunction via regulating hippocampal AMPK/NF-κB pathway in neuropathic pain mice. Neurobiol. Learn. Mem 2021, 182, 107463. [Google Scholar] [CrossRef]
- Fan, H.; Gong, N.; Li, T.F.; Ma, A.N.; Wu, X.Y.; Wang, M.W.; Wang, Y.X. The non-peptide GLP-1 receptor agonist WB4-24 blocks inflammatory nociception by stimulating β-endorphin release from spinal microglia. Br. J. Pharmacol. 2015, 172, 64–79. [Google Scholar] [CrossRef] [Green Version]
- Yoon, G.; Kim, Y.-K.; Song, J. Glucagon-like peptide-1 suppresses neuroinflammation and improves neural structure. Pharmacol. Res. 2020, 152, 104615. [Google Scholar] [CrossRef]
- Fujita, S.; Ushio, S.; Ozawa, N.; Masuguchi, K.; Kawashiri, T.; Oishi, R.; Egashira, N. Exenatide Facilitates Recovery from Oxaliplatin-Induced Peripheral Neuropathy in Rats. PLoS ONE 2015, 10, e0141921. [Google Scholar] [CrossRef]
- Cui, S.-S.; Feng, X.-B.; Zhang, B.-H.; Xia, Z.-Y.; Zhan, L.-Y. Exendin-4 attenuates pain-induced cognitive impairment by alleviating hippocampal neuroinflammation in a rat model of spinal nerve ligation. Neural Regen. Res. 2020, 15, 1333–1339. [Google Scholar] [CrossRef]
- Han, W.; Li, Y.; Cheng, J.; Zhang, J.; Chen, D.; Fang, M.; Xiang, G.; Wu, Y.; Zhang, H.; Xu, K.; et al. Sitagliptin improves functional recovery via GLP-1R-induced anti-apoptosis and facilitation of axonal regeneration after spinal cord injury. J. Cell. Mol. Med. 2020, 24, 8687–8702. [Google Scholar] [CrossRef]
- Ma, J.; Shi, M.; Zhang, X.; Liu, X.; Chen, J.; Zhang, R.; Wang, X.; Zhang, H. GLP-1R agonists ameliorate peripheral nerve dysfunction and inflammation via p38 MAPK/NF-κB signaling pathways in streptozotocin-induced diabetic rats. Int. J. Mol. Med. 2018, 41, 2977–2985. [Google Scholar] [CrossRef] [Green Version]
- Kuthati, Y.; Goutham Davuluri, V.N.; Yang, C.P.; Chang, H.C.; Chang, C.P.; Wong, C.S. Melatonin MT2 receptor agonist IIK-7 produces antinociception by modulation of ROS and suppression of spinal microglial activation in neuropathic pain rats. J. Pain Res. 2019, 12, 2473–2485. [Google Scholar] [CrossRef] [Green Version]
- Lv, J.; Li, Z.; She, S.; Xu, L.; Ying, Y. Effects of intrathecal injection of rapamycin on pain threshold and spinal cord glial activation in rats with neuropathic pain. Neurol Res. 2015, 37, 739–743. [Google Scholar] [CrossRef]
- Kuthati, Y.; Busa, P.; Goutham Davuluri, V.N.; Wong, C.S. Manganese Oxide Nanozymes Ameliorate Mechanical Allodynia in a Rat Model of Partial Sciatic Nerve-Transection Induced Neuropathic Pain. Int. J. Nanomed. 2019, 14, 10105–10117. [Google Scholar] [CrossRef] [Green Version]
- Kuthati, Y.; Busa, P.; Tummala, S.; Rao, V.N.; Davuluri, V.N.G.; Ho, Y.-P.; Wong, C.-S. Mesoporous Polydopamine Nanoparticles Attenuate Morphine Tolerance in Neuropathic Pain Rats by Inhibition of Oxidative Stress and Restoration of the Endogenous Antioxidant System. Antioxidants 2021, 10, 195. [Google Scholar] [CrossRef] [PubMed]
- Yang, C.-P.; Cherng, C.-H.; Wu, C.-T.; Huang, H.-Y.; Tao, P.-L.; Lee, S.-O.; Wong, C.-S. Intrathecal Ultra-Low Dose Naloxone Enhances the Antihyperalgesic Effects of Morphine and Attenuates Tumor Necrosis Factor-α and Tumor Necrosis Factor-α Receptor 1 Expression in the Dorsal Horn of Rats with Partial Sciatic Nerve Transection. Anesth. Analg. 2013, 117, 1493–1502. [Google Scholar] [CrossRef]
- Ouyang, H.; Bai, X.; Huang, W.; Chen, D.; Dohi, S.; Zeng, W. The antinociceptive activity of intrathecally administered amiloride and its interactions with morphine and clonidine in rats. J. Pain 2012, 13, 41–48. [Google Scholar] [CrossRef]
- Jaiswal, S.R.; Sontakke, S.D. Experimental evaluation of analgesic and anti-inflammatory activity of simvastatin and atorvastatin. Indian J. Pharmacol. 2012, 44, 475–479. [Google Scholar] [CrossRef]
- Haigler, H.J.; Spring, D.D. Comparison of the Analgesic Effects of Dimethyl Sulfoxide And Morphine. Ann. N. Y. Acad. Sci. 1983, 411, 19–27. [Google Scholar] [CrossRef]
- Hewedy, W.A. Linagliptin attenuates chronic post-ischemia pain: Possible anti-inflammatory and anti-oxidant mechanisms. Eur. J. Pharmacol. 2018, 828, 110–118. [Google Scholar] [CrossRef]
- Lindenlaub, T.; Sommer, C. Partial sciatic nerve transection as a model of neuropathic pain: A qualitative and quantitative neuropathological study. Pain 2000, 89, 97–106. [Google Scholar] [CrossRef]
- Challa, S.R. Surgical animal models of neuropathic pain: Pros and Cons. Int. J. Neurosci. 2015, 125, 170–174. [Google Scholar] [CrossRef] [PubMed]
- Drucker, D.J. The Cardiovascular Biology of Glucagon-like Peptide-1. Cell Metab. 2016, 24, 15–30. [Google Scholar] [CrossRef]
- Grieco, M.; Giorgi, A.; Gentile, M.C.; d’Erme, M.; Morano, S.; Maras, B.; Filardi, T. Glucagon-Like Peptide-1: A Focus on Neurodegenerative Diseases. Front. Neurosci. 2019, 13, 1112. [Google Scholar] [CrossRef] [Green Version]
- Llewellyn-Smith, I.J.; Marina, N.; Manton, R.N.; Reimann, F.; Gribble, F.M.; Trapp, S. Spinally projecting preproglucagon axons preferentially innervate sympathetic preganglionic neurons. Neuroscience 2015, 284, 872–887. [Google Scholar] [CrossRef] [Green Version]
- Oh, Y.S.; Jun, H.-S. Effects of Glucagon-Like Peptide-1 on Oxidative Stress and Nrf2 Signaling. Int. J. Mol. Sci 2017, 19, 26. [Google Scholar] [CrossRef] [Green Version]
- Sharma, S.K.; Panneerselvam, A.; Singh, K.P.; Parmar, G.; Gadge, P.; Swami, O.C. Teneligliptin in management of type 2 diabetes mellitus. Diabetes Metab. Syndr. Obes. Targets Ther. 2016, 9, 251–260. [Google Scholar] [CrossRef] [Green Version]
- Kamiko, K.; Aoki, K.; Kamiyama, H.; Taguri, M.; Shibata, E.; Ashiya, Y.; Minagawa, F.; Shinoda, K.; Nakajima, S.; Terauchi, Y. Comparison of the administration of teneligliptin every day versus every other day in Japanese patients with type 2 diabetes: A randomized non-inferior test. J. Clin. Pharmacol. 2015, 55, 144–151. [Google Scholar] [CrossRef] [PubMed]
- Syngle, A.; Chahal, S.; Vohra, K. Efficacy and tolerability of DPP4 inhibitor, teneligliptin, on autonomic and peripheral neuropathy in type 2 diabetes: An open label, pilot study. Neurol Sci. 2021, 42, 1429–1436. [Google Scholar] [CrossRef] [PubMed]
- Noda, T.; Ebihara, E.; Ueno, H.; Sadohara, K.; Tanaka, Y.; Nagatomo, Y.; Murakami, Y.; Yonamine, S.; Tsuchimochi, W.; Sakoda, H.; et al. Concurrent Use of Teneligliptin and Canagliflozin Improves Glycemic Control with Beneficial Effects on Plasma Glucagon and Glucagon-Like Peptide-1: A Single-Arm Study. Diabetes Ther. 2019, 10, 1835–1846. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Salcedo, I.; Tweedie, D.; Li, Y.; Greig, N.H. Neuroprotective and neurotrophic actions of glucagon-like peptide-1: An emerging opportunity to treat neurodegenerative and cerebrovascular disorders. Br. J. Pharmacol. 2012, 166, 1586–1599. [Google Scholar] [CrossRef] [Green Version]
- Li, H.; Jia, Z.; Li, G.; Zhao, X.; Sun, P.; Wang, J.; Fan, Z.; Lv, G. Neuroprotective effects of exendin-4 in rat model of spinal cord injury via inhibiting mitochondrial apoptotic pathway. Int. J. Clin. Exp. Pathol. 2015, 8, 4837–4843. [Google Scholar] [PubMed]
- Wiciński, M.; Wódkiewicz, E.; Słupski, M.; Walczak, M.; Socha, M.; Malinowski, B.; Pawlak-Osińska, K. Neuroprotective Activity of Sitagliptin via Reduction of Neuroinflammation beyond the Incretin Effect: Focus on Alzheimer’s Disease. Biomed. Res. Int. 2018, 2018, 6091014. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Perry, T.; Kindy, M.S.; Harvey, B.K.; Tweedie, D.; Holloway, H.W.; Powers, K.; Shen, H.; Egan, J.M.; Sambamurti, K.; et al. GLP-1 receptor stimulation preserves primary cortical and dopaminergic neurons in cellular and rodent models of stroke and Parkinsonism. Proc. Natl. Acad. Sci. USA 2009, 106, 1285–1290. [Google Scholar] [CrossRef] [Green Version]
- Chen, J.; Wang, Z.; Mao, Y.; Zheng, Z.; Chen, Y.; Khor, S.; Shi, K.; He, Z.; Li, J.; Gong, F.; et al. Liraglutide activates autophagy via GLP-1R to improve functional recovery after spinal cord injury. Oncotarget 2017, 8, 85949–85968. [Google Scholar] [CrossRef]
- Salim, H.M.; Fukuda, D.; Higashikuni, Y.; Tanaka, K.; Hirata, Y.; Yagi, S.; Soeki, T.; Shimabukuro, M.; Sata, M. Teneligliptin, a dipeptidyl peptidase-4 inhibitor, attenuated pro-inflammatory phenotype of perivascular adipose tissue and inhibited atherogenesis in normoglycemic apolipoprotein-E-deficient mice. Vasc. Pharm. 2017, 96–98, 19–25. [Google Scholar] [CrossRef] [PubMed]
- Aso, Y.; Kase, M.; Sagara, M.; Sakurai, S.; Iijima, T.; Tomaru, T.; Jojima, T.; Usui, I. Teneligliptin, a DPP-4 Inhibitor, Decreases Plasma Levels of Inflammatory Chemokines During a Standard Meal Test in Patients With Type 2 Diabetes. Am. J. Med. Sci. 2020, 360, 261–267. [Google Scholar] [CrossRef]
- Liu, X.; Cao, Y.; Zhang, Y.; Sun, B.; Liang, H. Teneligliptin inhibits lipopolysaccharide-induced cytotoxicity and inflammation in dental pulp cells. Int. Immunopharmacol. 2019, 73, 57–63. [Google Scholar] [CrossRef] [PubMed]
- Iwakura, T.; Zhao, Z.; Marschner, J.A.; Devarapu, S.K.; Yasuda, H.; Anders, H.J. Dipeptidyl peptidase-4 inhibitor teneligliptin accelerates recovery from cisplatin-induced acute kidney injury by attenuating inflammation and promoting tubular regeneration. Nephrol. Dial. Transplant. 2019, 34, 1669–1680. [Google Scholar] [CrossRef]
- Garrison, C.J.; Dougherty, P.M.; Kajander, K.C.; Carlton, S.M. Staining of glial fibrillary acidic protein (GFAP) in lumbar spinal cord increases following a sciatic nerve constriction injury. Brain Res. 1991, 565, 1–7. [Google Scholar] [CrossRef]
- Garrison, C.J.; Dougherty, P.M.; Carlton, S.M. GFAP expression in lumbar spinal cord of naive and neuropathic rats treated with MK-801. Exp. Neurol 1994, 129, 237–243. [Google Scholar] [CrossRef]
- Nesic, O.; Lee, J.; Johnson, K.M.; Ye, Z.; Xu, G.Y.; Unabia, G.C.; Wood, T.G.; McAdoo, D.J.; Westlund, K.N.; Hulsebosch, C.E.; et al. Transcriptional profiling of spinal cord injury-induced central neuropathic pain. J. NeuroChem. 2005, 95, 998–1014. [Google Scholar] [CrossRef] [PubMed]
- Song, P.; Zhao, Z.Q. The involvement of glial cells in the development of morphine tolerance. Neurosci. Res. 2001, 39, 281–286. [Google Scholar] [CrossRef]


Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kuthati, Y.; Rao, V.N.; Busa, P.; Wong, C.-S. Teneligliptin Exerts Antinociceptive Effects in Rat Model of Partial Sciatic Nerve Transection Induced Neuropathic Pain. Antioxidants 2021, 10, 1438. https://doi.org/10.3390/antiox10091438
Kuthati Y, Rao VN, Busa P, Wong C-S. Teneligliptin Exerts Antinociceptive Effects in Rat Model of Partial Sciatic Nerve Transection Induced Neuropathic Pain. Antioxidants. 2021; 10(9):1438. https://doi.org/10.3390/antiox10091438
Chicago/Turabian StyleKuthati, Yaswanth, Vaikar Navakanth Rao, Prabhakar Busa, and Chih-Shung Wong. 2021. "Teneligliptin Exerts Antinociceptive Effects in Rat Model of Partial Sciatic Nerve Transection Induced Neuropathic Pain" Antioxidants 10, no. 9: 1438. https://doi.org/10.3390/antiox10091438
APA StyleKuthati, Y., Rao, V. N., Busa, P., & Wong, C.-S. (2021). Teneligliptin Exerts Antinociceptive Effects in Rat Model of Partial Sciatic Nerve Transection Induced Neuropathic Pain. Antioxidants, 10(9), 1438. https://doi.org/10.3390/antiox10091438