The Anatomical and Functional Heterogeneity of the Mediodorsal Thalamus



{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
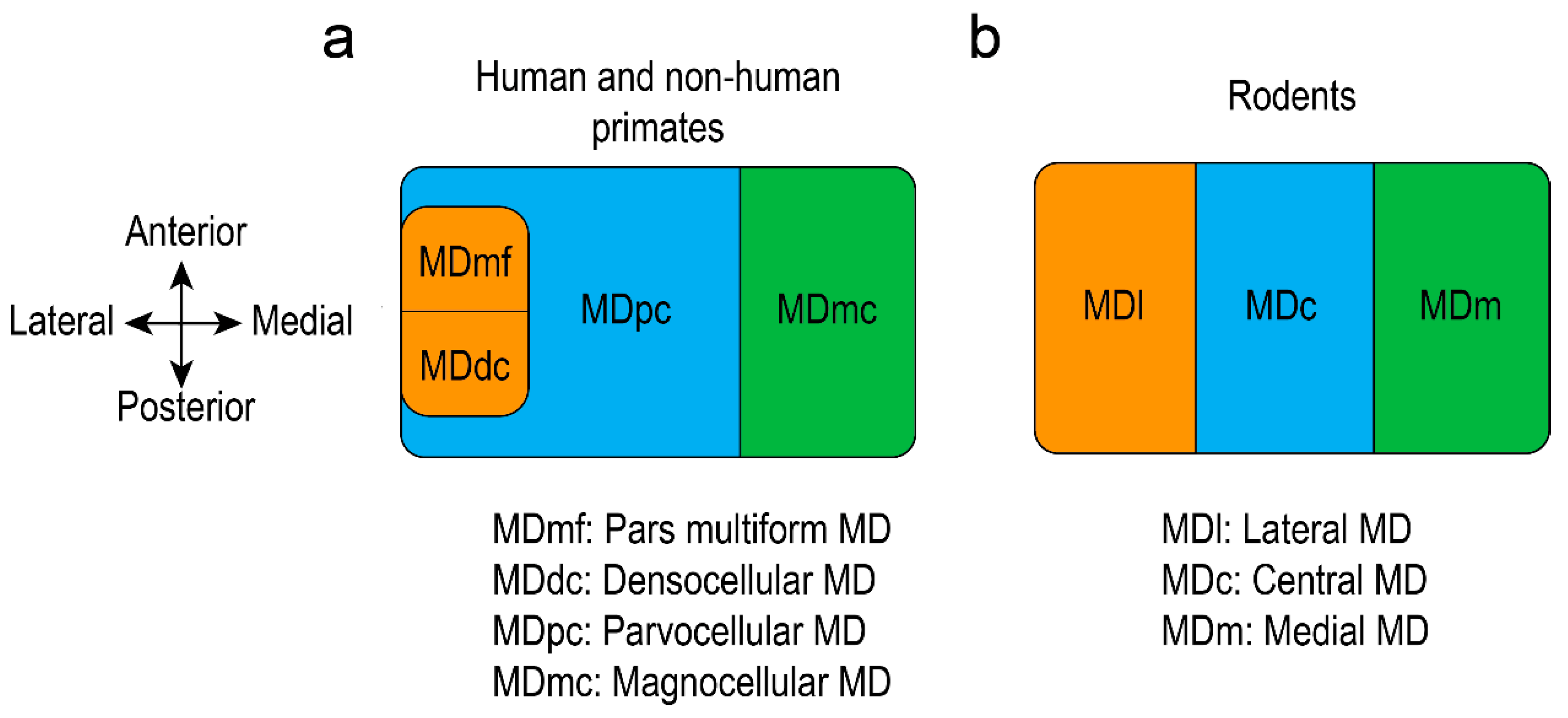
2. Thalamus Architecture
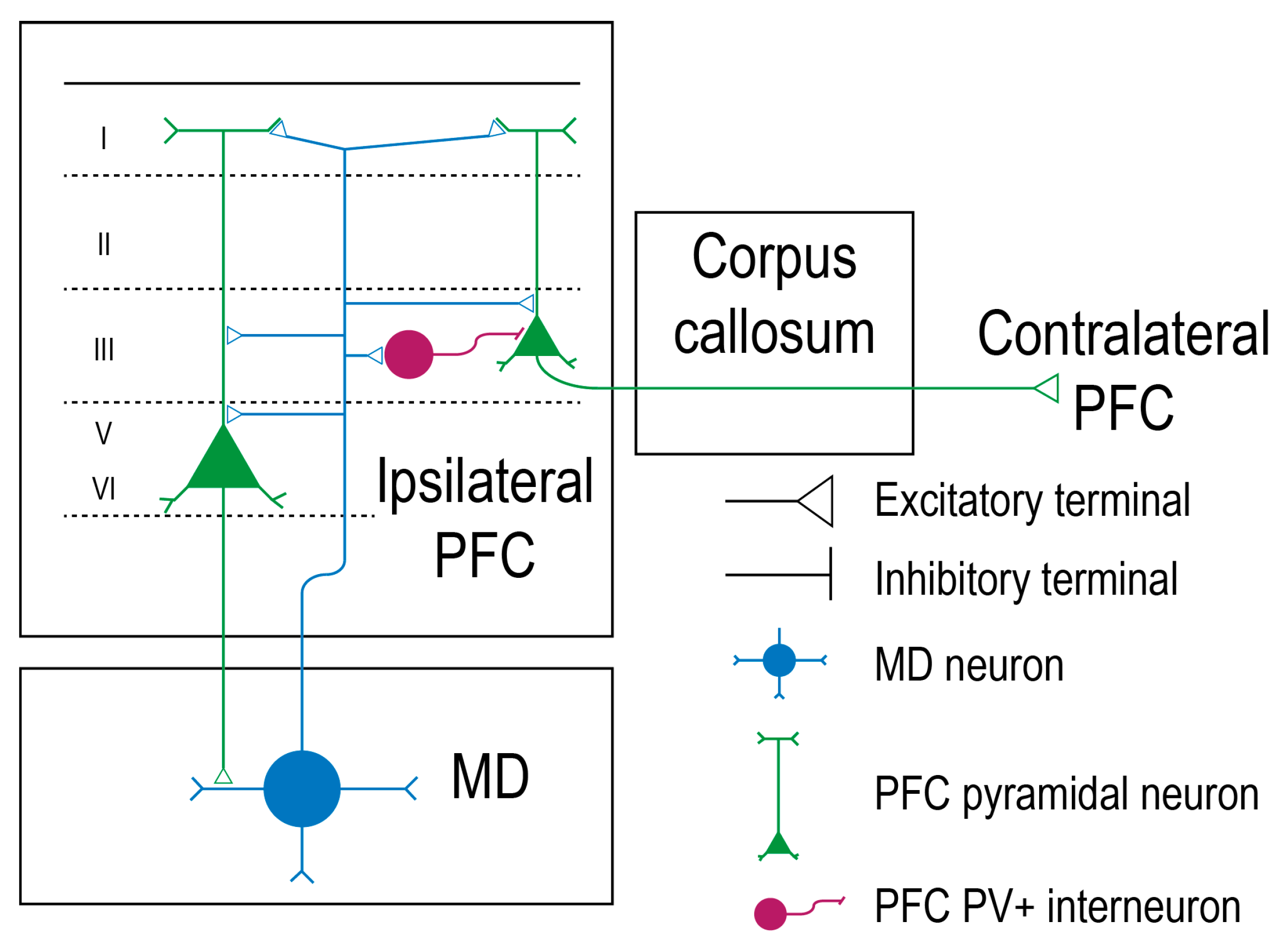
3. Mediodorsal Thalamic Relationship with the Prefrontal Cortex
4. Mediodorsal Thalamic Relationship with the Medial Temporal Regions
5. Mediodorsal Thalamic Relationship with Subcortical Structures
6. Mediodorsal Thalamic Relationship with the Cerebellum
7. Mediodorsal Thalamus Cytoarchitecture
8. Mediodorsal Thalamus Electrophysiological Responses in Relationship with Limbic-Related Areas
9. Mediodorsal Thalamus Neuromodulation
10. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Robert, P.V.; Stephanie, B.L.; Walter, B.H. Limbic circuitry of the midline thalamus. Neurosci. Biobehav. Rev. 2015, 54, 89–107. [Google Scholar] [CrossRef]
- Herrero, M.T.; Insausti, R.; Estrada, C. Thalamus: Anatomy; Elsevier Inc.: Amsterdam, The Netherlands, 2015; ISBN 9780123970251. [Google Scholar]
- Wolff, M.; Alcaraz, F.; Marchand, A.R.; Coutureau, E. Functional heterogeneity of the limbic thalamus: From hippocampal to cortical functions. Neurosci. Biobehav. Rev. 2015, 54, 120–130. [Google Scholar] [CrossRef]
- Block, A.E.; Dhanji, H.; Thompson-Tardif, S.F.; Floresco, S.B. Thalamic-prefrontal cortical-ventral striatal circuitry mediates dissociable components of strategy set shifting. Cereb. Cortex 2007, 17, 1625–1636. [Google Scholar] [CrossRef] [PubMed]
- Ouhaz, Z.; Fleming, H.; Mitchell, A.S. Cognitive functions and neurodevelopmental disorders involving the prefrontal cortex and mediodorsal thalamus. Front. Neurosci. 2018, 12, 1–18. [Google Scholar] [CrossRef] [PubMed]
- Mitchell, A.S.; Chakraborty, S. What does the mediodorsal thalamus do? Front. Syst. Neurosci. 2013, 7, 1–19. [Google Scholar] [CrossRef] [PubMed]
- Mitchell, A.S.; Gaffan, D. The magnocellular mediodorsal thalamus is necessary for memory acquisition, but not retrieval. J. Neurosci. 2008, 28, 258–263. [Google Scholar] [CrossRef] [PubMed]
- Harper, J.W.; Heath, R.G. Anatomic connections of the fastigial nucleus to the rostral forebrain in the cat. Exp. Neurol. 1973, 39, 285–292. [Google Scholar] [CrossRef]
- Saalmann, Y.B. Intralaminar and medial thalamic influence on cortical synchrony, information transmission and cognition. Front. Syst. Neurosci. 2014, 8, 1–8. [Google Scholar] [CrossRef]
- Grossman, E.J.; Inglese, M. The role of thalamic damage in mild traumatic brain injury. J. Neurotrauma 2016, 167, 163–167. [Google Scholar] [CrossRef]
- McFarland, N.R.; Haber, S.N. Thalamic relay nuclei of the basal ganglia form both reciprocal and nonreciprocal cortical connections, linking multiple frontal cortical areas. J. Neurosci. 2002, 22, 8117–8132. [Google Scholar] [CrossRef]
- Deecke, L.; Schwarz, D.W.F.; Fredrickson, J.M. Nucleus ventroposterior inferior (VPI) as the vestibular thalamic belay in the rhesus monkey i. field potential investigation. Exp. Brain Res. 1974, 20, 88–100. [Google Scholar] [CrossRef] [PubMed]
- Hirai, T.; Jones, E.G. A new parcellation of the human thalamus on the basis of histochemical staining. Brain Res. Rev. 1989, 14, 1–34. [Google Scholar] [CrossRef]
- Kosif, R. The Thalamus: A review of its functional anatomy. Med. Res. Arch. 2016, 4. Available online: https://journals.ke-i.org/index.php/mra/article/view/740 (accessed on 8 September 2020).
- Krauth, A.; Blanc, R.; Poveda, A.; Jeanmonod, D.; Morel, A.; Székely, G. NeuroImage A mean three-dimensional atlas of the human thalamus: Generation from multiple histological data. Neuroimage 2010, 49, 2053–2062. [Google Scholar] [CrossRef] [PubMed]
- Jones, E.G. The Thalamus; Springer: New York, NY, USA; Plenum Press: New York, NY, USA, 1985; ISBN 9781461357049. [Google Scholar] [CrossRef]
- Bay, H.H.; Çavdar, S. Regional connections of the mediodorsal thalamic nucleus in the rat. J. Integr. Neurosci. 2013, 12, 201–219. [Google Scholar] [CrossRef]
- Paxinos, G.; Franklin, K.B.J. The Mouse Brain in Stereotaxic Coordinates, 2nd ed.; Academic Press Inc.: New York, NY, USA, 2001; ISBN 0125476361. [Google Scholar]
- Bennett, C.; Gale, S.D.; Garrett, M.E.; Newton, M.L.; Callaway, E.M.; Murphy, G.J.; Olsen, S.R. Higher-order thalamic circuits channel parallel streams of visual information in mice. Neuron 2019, 102, 477–492.e5. [Google Scholar] [CrossRef]
- Brodmann, K.; Garey, J.L. Brodmann’s Localisation in the Cerebral Cortex; Springer US: New York, NY, USA, 2006; ISBN 9780387269177. [Google Scholar] [CrossRef]
- Groenewegen, H.J.; Wright, C.I.; Uylings, H.B.M. The anatomical relationships of the prefrontal cortex with limbic structures and the basal ganglia. J. Psychopharmacol. 1997, 11, 99–106. [Google Scholar] [CrossRef]
- Goldman-rakic, P.S.; Porrino, L.J. The primate mediodorsal (MD) nucleus and its projection to the frontal lobe. J. Comp. Neurol. 1985, 560, 535–560. [Google Scholar] [CrossRef]
- Murray Sherman, S.; Guillery, R.W. The role of the thalamus in the flow of information to the cortex. Philos. Trans. R. Soc. London. Series B Biol. Sci. 2002, 1695–1708. [Google Scholar] [CrossRef]
- Kuroda, M.; Yokofujita, J.; Murakami, K. An ultrastructural study of the neural circuit between the prefrontal cortex and the mediodorsal nucleus of the thalamus. Prog. Neurobiol. 1998, 54, 417–458. [Google Scholar] [CrossRef]
- Jones, E.G. Thalamic circuitry and thalamocortical synchrony. Philos. Trans. R. Soc. London. Series B Biol. Sci. 2002, 357, 1659–1673. [Google Scholar] [CrossRef]
- Miller, E.K.; Cohen, J.D. An integrate theory of PFC function. Annu. Rev. Neurosci. 2001, 24, 167–202. [Google Scholar] [CrossRef] [PubMed]
- Myers-Schulz, B.; Koenigs, M. Implications for mood and anxiety disorders. Mol. Psychiatry 2012, 17, 132–141. [Google Scholar] [CrossRef] [PubMed]
- Ongü, D.; Ferry, A.T.; Price, J.L. Architectonic subdivision of the human orbital and medial prefrontal cortex. J. Comp. Neurol. 2003, 449, 425–449. [Google Scholar] [CrossRef]
- Carmichael, S.T.; Price, J.L. Architectonic subdivision of the orbital and medial prefrontal cortex in the macaque monkey. J. Comp. Neurol. 1994, 346, 66–402. [Google Scholar] [CrossRef] [PubMed]
- Barbas, H.; Pandya, D.N. Architecture and intrinsic connections of the prefrontal cortex i n the rhesus monkey. J. Comp. Neurol. 1989, 286, 353–375. [Google Scholar] [CrossRef]
- Bachevalier, J.; Meunier, M.; Lu, M.X.; Ungerleider, L.G. Thalamic and temporal cortex input to medial prefrontal cortex in rhesus monkeys. Exp. Brain Res. 1997, 115, 430–444. [Google Scholar] [CrossRef]
- Morecraft, R.J.; Geula, C.; Mesulam, M. Cytoarchitecture and neural afferents of orbitofrontal cortex in the brain of the monkey. J. Comp. Neurol. 1992, 358, 341–358. [Google Scholar] [CrossRef]
- Ray, J.P.; Price, J.L. The organization of projections from the mediodorsal nucleus of the thalamus to orbital and medial prefrontal cortex in macaque monkeys. J. Comp. Neurol. 1993, 337, 1–31. [Google Scholar] [CrossRef]
- Erickson, S.L.; Lewis, D.A. Cortical connections of the lateral mediodorsal thalamus in cynomolgus monkeys. J. Comp. Neurol. 2004, 473, 107–127. [Google Scholar] [CrossRef]
- Preuss, T.M.; Goldman-Rakic, P.S. Crossed corticothalamic and thalamocortical connections of macaque prefrontal cortex. J. Comp. Neurol. 1987, 257, 269–281. [Google Scholar] [CrossRef]
- Russchen, F.T.; Amaral, D.G.; Price, J.L. The afferent input to the magnocellular division of the mediodorsal thalamic nucleus in the monkey, Macaca fascicularis. J. Comp. Neurol. 1987, 256, 175–210. [Google Scholar] [CrossRef] [PubMed]
- Barbas, H.; Henion, T.H.H.; Dermon, C.R. Diverse thalamic projections to the prefrontal cortex in the rhesus monkey. J. Comp. Neurol. 1991, 313, 65–94. [Google Scholar] [CrossRef] [PubMed]
- Giguere, M.; Goldman-Racik, P.S. Mediodorsal nucleus: Areal, laminar, and tangential distribution of afferents and efferents in the frontal lobe of rhesus monkeys. J. Comp. Neurol. 1988, 227, 195–213. [Google Scholar] [CrossRef] [PubMed]
- Kievit, J.; Kuypers, H.G.J.M. Brain Organization of the Thalamo-Cortical Connexions to the Frontal Lobe in the Rhesus Monkey. Exp. Brain Res. 1977, 322, 299–322. [Google Scholar]
- Ray, J.P.; Price, J.L. The organization of the thalamocortical connections of the mediodorsal thalamic nucleus in the rat, related to the ventral forebrain - prefrontal cortex topography. J. Comp. Neurol. 1992, 323, 167–197. [Google Scholar] [CrossRef]
- Burwell, R.; Witter, M.; Amarall, D. Perirhinal and Postrhinal Cortices of the Rat: A Review of the Neuroanatomical Literature and Comparison with Findings from the Monkey Brain. Hippocampus 1995, 5, 390–408. [Google Scholar] [CrossRef]
- Saunders, R.C.; Mishkin, Æ.M.; Aggleton, J.P. Projections from the entorhinal cortex, perirhinal cortex, presubiculum, and parasubiculum to the medial thalamus in macaque monkeys: Iden-tifying different pathways using disconnection techniques. Exp. Brain Res. 2005, 167, 1–16. [Google Scholar] [CrossRef]
- Insausti, R.; Amaral, D.G.; Cowan, W.M. The entorhinal cortex of the monkey. J. Comp. Neurol. 1987, 264, 356–395. [Google Scholar] [CrossRef]
- Gower, E.C. Efferent Projections from Limbic Cortex of the Temporal Pole to the Magnocellular Medial Dorsal Nucleus in the Rhesus Monkey. J. Comp. Neurol. 1989, 280, 343–358. [Google Scholar] [CrossRef]
- Baleydier, C.; Mauguiere, F. The duality of the cingulate gyrus in monkey neuroanatomical study and functional hypothesis. Brain J. Neurol. 1980, 103, 525–554. [Google Scholar] [CrossRef]
- Vogt, B.; Pandya, D. Cingulate Cortex of the Rhesus Monkey: 11. Cortical Merents. J. Comp. Neurol. 1987, 262, 271–289. [Google Scholar] [CrossRef] [PubMed]
- Barbas, H. Anatomic Organization of Basoventral and Mediodorsal Visual Recipient Prefrontal Regions in the Rhesus Monkey. J. Comp. Neurol. 1988, 276, 313–342. [Google Scholar] [CrossRef] [PubMed]
- Van Hoesen, G.W.; Pandya, D.N.; Butters, N. Some connections of the entorhinal (area 28) and perirhinal (area 35) cortices of the rhesus monkey 11 frontal lobe afferents. Brain Res. 1975, 95, 25–38. [Google Scholar] [CrossRef]
- Ungerleider, G.; Gaffan, D.; Pelak, S. Experimental brain research 9. Exp. Brain Res. 1989, 76, 473–484. [Google Scholar] [CrossRef]
- Suzuki, W.A.; Amaral, D.G. Perirhinal and Parahippocampal Cortices of the Macaque Monkey: Cortical Afferents. J. Comp. Neurol. 1994, 350, 497–533. [Google Scholar] [CrossRef]
- Webster, M.J.; Bachevalier, J.; Leslie, G. Connections of Inferior Temporal Areas TEO and TE with Parietal and Frontal Cortex in Macaque Monkeys. Cereb. Cortex 1994, 4, 470–483. [Google Scholar] [CrossRef]
- Arikuni, T.; Sako, H.; Murata, A. Neuroscience research. Neurosci. Res. 1994, 21, 19–39. [Google Scholar] [CrossRef]
- Baylis, L.L.; Rolls, E.T.; Baylis, G.C. Afferent connections of the caudolateral orbitofrontal cortex taste area of the primate. Neuroscience 1995, 64, 801–812. [Google Scholar] [CrossRef]
- Aggleton, J.P.; Desimone, R.; Mishkin, M. The Origin, Course, and Termination of the Hippocampothalamic Projections in the Macaque. J. Comp. Neurol. 1986, 243, 409–421. [Google Scholar] [CrossRef]
- Haber, S.N.; Calzavara, R. The cortico-basal ganglia integrative network: The role of the thalamus. Brain Res. Bull. 2009, 78, 69–74. [Google Scholar] [CrossRef]
- Aggleton, J.P.; Mishkin, M. Projections of the amygdala to the thalamus in the cynomolgus monkey. J. Comp. Neurol. 1984, 222, 56–68. [Google Scholar] [CrossRef] [PubMed]
- Krettek, J.E.; Price, J.L. The Cortical Projectioris of the Mediodorsal Nucleus and Adjacent Thalamic Nuclei in the Rat. J. Comp. Neurol. 1977, 171, 157–191. [Google Scholar] [CrossRef] [PubMed]
- Krettek, E.; Price, L. Projections from the Amygdaloid Complex to the Cerebral Cortex and Thalamus in the Rat and Cat 1. J. Comp. Neurol. 1977, 172, 687–722. [Google Scholar] [CrossRef] [PubMed]
- Porrino, L.; Crane, A.; Goldman-Rakic, P. Direct and Indirect Pathways from the Amygdala to the Frontal Lobe in Rhesus Monkeys. J. Comp. Neurol. 1981, 198, 121–136. [Google Scholar] [CrossRef] [PubMed]
- Sapolsky, R.M.; Eichenbaum, H. Thalamocortical Mechanisms in Odor-Guided Behavior. Brain Behav. Evol. 1980, 17, 276–290. [Google Scholar] [CrossRef] [PubMed]
- Turner, B.H.; Mortimer, M.; Knapp, M. Organization of the amygdalopetal projections from modality-specific cortical association areas in the monkey. J. Comp. Neurol. 1980, 191, 515–543. [Google Scholar] [CrossRef]
- Turner, B.H.; Gupta, K.C.; Mishkin, M. The Locus and Cytoarcvvhitecture of the Projection Areas of the Olfactory Bulb in Macaca mulatta. J. Comp. Neurol. 1978, 177, 381–396. [Google Scholar] [CrossRef]
- Berendse, H.W.; Groenewegen, H.J. Organization of the thalamostriatal projections in the rat, with special emphasis on the ventral striatum. J. Comp. Neurol. 1990, 299, 187–228. [Google Scholar] [CrossRef]
- Groenewegen, H.J.; Graaf, Y.G.; Smeets, W.J.A.J. Integration and segregation of limbic cortico-striatal loops at the thalamic level: An experimental tracing study in rats. J. Chem. Neuroanat. 1999, 16, 167–185. [Google Scholar] [CrossRef]
- Giménez-Amaya, J.M.; McFarland, N.R.; De Las Heras, S.; Haber, S.N. Organization of thalamic projections to the ventral striatum in the primate. J. Comp. Neurol. 1995, 354, 127–149. [Google Scholar] [CrossRef]
- Haber, S.; McFarland, R.N. The place of the thalamus in frontal cortical-basal ganglia circuits. Neuroscience 2001, 7, 315–324. [Google Scholar] [CrossRef] [PubMed]
- Groenewegen, H.J. Organization of the afferent connections of the mediodorsal thalamic nucleus in the rat, related to the mediodorsal-prefrontal topography. Neuroscience 1988, 24, 379–431. [Google Scholar] [CrossRef]
- Hallanger, A.E.; Levey, A.I.; Lee, H.J.; Rye, D.B.; Wainer, B.H. The origins of cholinergic and other nonspecific afferents to the thalamus in the rat. J. Comp. Neurol. 1987, 262, 105–124. [Google Scholar] [CrossRef] [PubMed]
- Parent, A. Extrinsic connections of the basal ganglia. Trends Neurosci. 1990, 13, 254–258. [Google Scholar] [CrossRef]
- Parent, A.; Hazrati, L.-N. The cortico-basal ganglia-thalamo-corticalloop. Abstr. Brain Res. Rev. 1995, 20, 91–127. [Google Scholar] [CrossRef]
- Yamamoto, T.; Yoshida, K.; Yoshikawa, H.; Kishimoto, Y.; Oka, H. The medial dorsal nucleus is one of the thalamic relays of the cerebellocerebral responses to the frontal association cortex in the monkey: Horseradish peroxidase and fluorescent dye double staining study. Brain Res. 1992, 579, 315–320. [Google Scholar] [CrossRef]
- Hintzen, A.; Pelzer, E.A.; Tittgemeyer, M. Thalamic interactions of cerebellum and basal ganglia. Brain Struct. Funct. 2017, 223, 569–587. [Google Scholar] [CrossRef]
- Tanibuchi, I.; Kitano, H.; Jinnai, K. Substantia nigra output to prefrontal cortex via thalamus in monkeys. i. electrophysiological identification of thalamic relay neurons. J. Neurophysiol. 2009, 102, 2933–2945. [Google Scholar] [CrossRef]
- Middleton, F.A.; Strick, P.L. Cerebellar Projections to the Prefrontal Cortex of the Primate. J. Neurosci. 2001, 21, 700–712. [Google Scholar] [CrossRef]
- Ristanović, D.; Milošević, N.T.; Stefanović, B.D.; Marić, D.L.; Rajković, K. Morphology and classification of large neurons in the adult human dentate nucleus: A qualitative and quantitative analysis of 2D images. Neurosci. Res. 2010, 67, 1–7. [Google Scholar] [CrossRef]
- Gallay, M.N.; Jeanmonod, D.; Liu, J.; Morel, A. Human pallidothalamic and cerebellothalamic tracts: Anatomical basis for functional stereotactic neurosurgery. Brain Struct. Funct. 2008, 212, 443–463. [Google Scholar] [CrossRef]
- Pelzer, E.A.; Melzer, C.; Timmermann, L.; von Cramon, D.Y.; Tittgemeyer, M. Basal ganglia and cerebellar interconnectivity within the human thalamus. Brain Struct. Funct. 2017, 222, 381–392. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.-Y.; Wang, J.-J.; Zhu, J.-N. Cerebellar fastigial nucleus: From anatomic construction to physiological functions. Cerebellum Ataxias 2016, 3, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Fujita, H.; Kodama, T.; du Lac, S. Modular output circuits of the fastigial nucleus for diverse motor and nonmotor functions of the cerebellar vermis. bioRxiv 2020, 9. [Google Scholar] [CrossRef]
- Anticevic, A.; Yang, G.; Savic, A.; Murray, J.D.; Cole, M.W.; Repovs, G.; Pearlson, G.D.; Glahn, D.C. Mediodorsal and visual thalamic connectivity differ in schizophrenia and bipolar disorder with and without psychosis history. Schizophr. Bull. 2014, 40, 1227–1243. [Google Scholar] [CrossRef]
- Parker, K.L.; Narayanan, N.S.; Andreasen, N.C. The therapeutic potential of the cerebellum in schizophrenia. Front. Syst. Neurosci. 2014, 8, 1–11. [Google Scholar] [CrossRef]
- Peng, H.J.; Zheng, H.R.; Ning, Y.P.; Zhang, Y.; Shan, B.C.; Zhang, L.; Yang, H.C.; Liu, J.; Li, Z.X.; Zhou, J.S.; et al. Abnormalities of cortical-limbic-cerebellar white matter networks may contribute to treatment-resistant depression: A diffusion tensor imaging study. BMC Psychiatry 2013, 13, 1–8. [Google Scholar] [CrossRef]
- Ito, M. Control of mental activities by internal models in the cerebellum. Nat. Rev. Neurosci. 2008, 9, 304–313. [Google Scholar] [CrossRef]
- Schutter, D.J.L.G.; Van Honk, J. The cerebellum on the rise in human emotion. Cerebellum 2005, 4, 290–294. [Google Scholar] [CrossRef]
- Strata, P. The Emotional Cerebellum. Cerebellum 2015, 14, 570–577. [Google Scholar] [CrossRef]
- Szalóki, N.; Krieger, J.W.; Komáromi, I.; Tóth, K.; Vámosi, G. Evidence for homodimerization of the c-Fos transcription factor in live cells revealed by fluorescence microscopy and computer modeling. Mol. Cell. Biol. 2015, 35, 3785–3798. [Google Scholar] [CrossRef]
- Moers-Hornikx, V.M.P.; Sesia, T.; Basar, K.; Lim, L.W.; Hoogland, G.; Steinbusch, H.W.M.; Gavilanes, D.A.W.D.; Temel, Y.; Vles, J.S.H. Cerebellar nuclei are involved in impulsive behaviour. Behav. Brain Res. 2009, 203, 256–263. [Google Scholar] [CrossRef] [PubMed]
- Rogers, T.D.; Price, E.D.; Detlef, H.H.; Dan, G.; Guy, M.; Charles, D.B. Connecting the dots of the cerebro-cerebellar role in cognitive function: Neuronal pathways for cerebellar modulation of dopamine release in the prefrontal cortex. Synapse 2011, 65, 1–14. [Google Scholar] [CrossRef] [PubMed]
- Strata, P.; Scelfo, B.; Sacchetti, B. Involvement of cerebellum in emotional behavior. Physiol. Res. 2011, 60, S39–S48. [Google Scholar] [CrossRef]
- Kuroda, M.; López-Mascaraque, L.; Price, J.L. Neuronal and synaptic composition of the mediodorsal thalamic nucleus in the rat: A light and electron microscopic golgi study. J. Comp. Neurol. 1992, 326, 61–81. [Google Scholar] [CrossRef]
- Leonard, C.M. The Connections of the Dorsomedial Nuclei. Brain Behav. Evol. 1972, 6, 524–541. [Google Scholar] [CrossRef]
- Kuroda, M.; Price, J.L. Synaptic organization of projections from basal forebrain structures to the mediodorsal thalamic nucleus of the rat. J. Comp. Neurol. 1991, 303, 513–533. [Google Scholar] [CrossRef]
- Jones, E.G. Synchrony in the interconnected circuitry of the thalamus and cerebral cortex. Ann. N. Y. Acad. Sci. 2009, 1157, 10–23. [Google Scholar] [CrossRef]
- Sherman, S.M. Tonic and burst firing: Dual modes of thalamocortical relay. Trends Neurosci. 2001, 24, 122–126. [Google Scholar] [CrossRef]
- Lavin, A.; Grace, A.A. Dopamine modulates the responsivity of mediodorsal thalamic cells recorded in vitro. J. Neurosci. 1998, 18, 10566–10578. [Google Scholar] [CrossRef]
- Radhakrishnan, V.; Tsoukatos, J.; Davis, K.D.; Tasker, R.R.; Lozano, A.M.; Dostrovsky, J.O. A comparison of the burst activity of lateral thalamic neurons in chronic pain and non-pain patients. Pain 1999, 80, 567–575. [Google Scholar] [CrossRef]
- Magnin, M.; Morel, A.; Jeanmonod, D. Single-unit analysis of the pallidum, thalamus and subthalamic nucleus in parkinsonian patients. Neuroscience 2000, 96, 549–564. [Google Scholar] [CrossRef]
- Parnaudeau, S.; Bolkan, S.S.; Kellendonk, C. The Mediodorsal Thalamus: An Essential Partner of the Prefrontal Cortex for Cognition. Biol. Psychiatry 2018, 83, 648–656. [Google Scholar] [CrossRef] [PubMed]
- Parnaudeau, S.; O’Neill, P.K.; Bolkan, S.; Ward, R.D.; Abbas, A.I.; Roth, B.L.; Balsam, P.; Gordon, J.A.; Kellendonk, C. Inhibition of medio-dorsal thalamus disrupts thalamo-frontal connectivity and cognition. Neuron 2013, 77, 1151–1162. [Google Scholar] [CrossRef] [PubMed]
- Bolkan, S.S.; Stujenske, J.M.; Parnaudeau, S.; Spellman, T.J.; Rauffenbart, C.; Abbas, A.I.; Harris, A.Z.; Gordon, J.A.; Kellendonk, C. Thalamic projections sustain prefrontal activity during working memory maintenance. Nat. Neurosci. 2017, 20, 987–996. [Google Scholar] [CrossRef] [PubMed]
- Floresco, S.B.; Braaksma, D.N.; Phillips, A.G. Thalamic–Cortical–Striatal Circuitry Subserves Working Memory during Delayed Responding on a Radial Arm Maze. J. Neurosci. 1999, 19, 11061–11071. [Google Scholar] [CrossRef]
- Salmeron, J.; Escames, G.; PerezJaranay, J.; Vives, F. Electrophysiological response of neurons of the mediodorsal nucleus of the thalamus to the stimulation of the nucleus accumbens in the rat. Neurosci. Res. Commun. 1996, 19, 109–114. [Google Scholar] [CrossRef]
- Floresco, S.B. Grace A Gating of hippocampal-evoked activity in prefrontal cortical neurons by inputs from the mediodorsal thalamus and ventral tegmental area. J. Neurosci. 2003, 23, 3930–3943. [Google Scholar] [CrossRef]
- Beierlein, M.; Connors, B.W. Short-term dynamics of thalamocortical and intracortical synapses onto layer 6 neurons in neocortex. J. Neurophysiol. 2002, 88, 1924–1932. [Google Scholar] [CrossRef]
- Zoppelt, D.; Koch, B.; Schwarz, M.; Daum, I. Involvement of the mediodorsal thalamic nucleus in mediating recollection and familiarity. Neuropsychologia 2003, 41, 1160–1170. [Google Scholar] [CrossRef]
- Van Der Werf, Y.D.; Witter, M.P.; Uylings, H.B.M.; Jolles, J. Neuropsychology of infarctions in the thalamus: A review. Neuropsychologia 2000, 38, 613–627. [Google Scholar] [CrossRef]
- Nakajima, M.; Halassa, M.M. Thalamic control of functional cortical connectivity. Curr. Opin. Neurobiol. 2017, 44, 127–131. [Google Scholar] [CrossRef] [PubMed]
- Lee, S.; Shin, H.S. The role of mediodorsal thalamic nucleus in fear extinction. J. Anal. Sci. Technol. 2016, 7, 13. [Google Scholar] [CrossRef]
- Varela, C. Thalamic neuromodulation and its implications for executive networks. Front. Neural Circuits 2014, 8, 1–22. [Google Scholar] [CrossRef] [PubMed]
- Jahnsen, A.H.; Llinás, R.R. Electrophysiology of mammalian thalamic neurons in vitro. Nature 1982, 279, 406–408. [Google Scholar] [CrossRef]
- Lee, S.; Ahmed, T.; Lee, S.; Kim, H.; Choi, S.; Kim, D.S.; Kim, S.J.; Cho, J.; Shin, H.S. Bidirectional modulation of fear extinction by mediodorsal thalamic firing in mice. Nat. Neurosci. 2012, 15, 308–314. [Google Scholar] [CrossRef]
- McCormick, D.A.; von Krosigk, M. Corticothalamic activation modulates thalamic firing through glutamate “metabotropic” receptors. Proc. Natl. Acad. Sci. USA 1992, 89, 2774–2778. [Google Scholar] [CrossRef]
- Mitchell, A.S. The mediodorsal thalamus as a higher order thalamic relay nucleus important for learning and decision-making. Neurosci. Biobehav. Rev. 2015, 54, 76–88. [Google Scholar] [CrossRef]



© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Georgescu, I.A.; Popa, D.; Zagrean, L. The Anatomical and Functional Heterogeneity of the Mediodorsal Thalamus. Brain Sci. 2020, 10, 624. https://doi.org/10.3390/brainsci10090624
Georgescu IA, Popa D, Zagrean L. The Anatomical and Functional Heterogeneity of the Mediodorsal Thalamus. Brain Sciences. 2020; 10(9):624. https://doi.org/10.3390/brainsci10090624
Chicago/Turabian StyleGeorgescu, Ioana Antoaneta, Daniela Popa, and Leon Zagrean. 2020. "The Anatomical and Functional Heterogeneity of the Mediodorsal Thalamus" Brain Sciences 10, no. 9: 624. https://doi.org/10.3390/brainsci10090624
APA StyleGeorgescu, I. A., Popa, D., & Zagrean, L. (2020). The Anatomical and Functional Heterogeneity of the Mediodorsal Thalamus. Brain Sciences, 10(9), 624. https://doi.org/10.3390/brainsci10090624