Abstract
Human β-defensin 3 (hBD3) is an antimicrobial peptide and a chemoattractant molecule expressed in human keratinocytes. Lysates prepared from Lactoplantibacillus plantarum KG (KG lysates), a fermented lactobacillus strain with Ginseng extracts, significantly induced hBD3 in human keratinocytes. hBD3 induced by KG lysates affected the internalization of Staphylococcus aureus by human keratinocytes. The intracellular pathway for the induction of hBD3 by KG lysates was examined using inhibitors. Briefly, KG lysates stimulated the late signaling pathway including JAK1/2, which might be induced by the EGFR- or IFN-γ-mediated pathway, and affected hBD3 induction. Through this study, we confirmed that KG lysates have the ability to control bacterial infections, especially S. aureus, in human keratinocytes. Thus, KG lysates can be provided in various formulations for the development of food and pharmaceutical materials to control bacterial infection.
1. Introduction
Human beta defensin-3 (hBD3) is a novel defensin peptide composed of a 45-amino-acid protein. hBD3 acts as an anti-microbial peptide as well as a chemoattractant. The N-terminus of hBD3 has antimicrobial properties against Staphylococcus aureus and Candida albicans [1]. As a chemoattractant, hBD3 induces inflammatory cytokines such as IL-6, IL-8, IL-1β, and TNF-α in monocytes. In a mouse model, hBD3 recruited neutrophils and macrophages [2]. In addition, hBD3 appears to be involved in skin wound healing. hBD3 gene expression was increased in primary keratinocytes by platelet-related growth factor via the EGFR [3]. Many epithelia express hBD3 upon cytokine and microbial stimuli [4,5]. In keratinocytes as well as in monocytes, epidermal growth factor receptor (EGFR) ligands such as transforming growth factor alpha (TGF-α), epidermal growth factor (EGF), amphiregulin, and heparin-binding EGF-like growth factor (HB-EGF) induce hBD3. However, individual blocking of these ligands did not inhibit hBD3 expression in keratinocytes, suggesting that several EGFR ligands may participate in the regulation of hBD3 [6].
Probiotic Escherichia coli Nissle 1917 induces beta-defensin in intestinal epithelial cells through the activation of nuclear factor kappa B (NF-kB) and activator protein 1 (AP-1) [7]. Caco-2 cells exposed to Lactoplantibacillus plantarum induce beta-defensin expression through the Toll-like receptor 2 (TLR2) signaling pathway and secrete it out of the cells in a dose- and time-dependent manner [8]. Similarly, Lactobacillus helveticus SBT2171 (LH2171) induced the expression of beta-defensin in Caco-2 cells using surface layer protein (SLP) of LH2171, which stimulates beta-defensin expression by TLR2-mediated activation of c-Jun N-terminal kinase (JNK) signaling [9]. The expression of beta-defensin affects the gut microbial community and modulates responses to pathogenic infections. Human skin keratinocytes also express beta-defensin including hBD2 and hBD3, which may play important antibacterial roles in inflamed epidermis for host defense against skin pathogens [10].
L. plantarum is an emerging probiotic in skin and intestinal health, improving skin conditions such as itching, aging, and moisture retention [11,12,13]. Not only food companies but also global pharmaceutical companies are aggressively launching probiotic products with L. plantarum through active research and are targeting consumers. Both L. plantarum K8 and L. plantarum KG were isolated from the traditional Korean fermented food Kimchi. We previously demonstrated the effects of L. plantarum K8 and its cell wall components on immune regulation and skin health [14,15,16,17,18]. Here, we examine the immune regulatory effect of L. plantarum KG lysates with a focus on hBD3.
2. Materials and Methods
2.1. Cell Line
HaCaT cells, immortalized human keratinocytes, were maintained in Dulbecco’s modified Eagle’s medium (DMEM, WELGENE, Gyeongsan, Korea). Cells were supplemented with 10% (v/v) heat-inactivated and filtered fetal bovine serum (FBS, WELGENE), 100 U/mL penicillin and 100 μg/mL streptomycin (P/S, WELGENE). These cells were maintained at 37 °C with 5% CO2 in a humidified incubator.
2.2. Bacterial Lysate Preparation
L. plantarum KG (KCTC 14102BP) was cultured in 1 L of MRS broth (BD Biosciences, San Jose, CA, USA) containing ginseng extracts at 37 °C overnight, and then cells were harvested by centrifugation at 8000 rpm for 10 min. Cells were re-suspended in sterilized water and disrupted by microfluidization at 15,000 psi, 5 times. Disrupted L. plantarum KG lysates (KG lysates) were freeze-dried and cells were re-suspended in nuclease-free water (Promega, Madison, WI, USA) and concentrated to 1011 colony-forming units (CFU)/mL for the experiments. L. plantarum K8 (KCTC 10887BP) lysates (K8 lysates) were also prepared according to the above protocol.
2.3. Cell Viability Assay
HaCaT cells were seeded onto 96-well culture plates and incubated overnight. After stabilization, cells were treated with KG lysates (107, 108, 109 CFU/mL) for 24, 48, 72 h. Cell viability was determined using an EZ-Cytox cell viability assay kit (Daeil Lab Service, Cheongwon, Korea) according to the manufacturer’s instructions. Absorbance was measured using a microplate reader (Eppendorf BioPhotometer, Hamburg, Germany) at a test wavelength of 450 nm and a reference wavelength of 590 nm.
2.4. Real-Time PCR
Total RNA was isolated from HaCaT cells using RNAiso Plus (TaKaRa Bio, Shiga, Japan). Total RNA served as a template in a reverse transcription reaction using PrimeScriptTM RT Master Mix (TaKaRa Bio). A real-time polymerase chain reaction (PCR) was performed using CFX ConnectTM Real-Time PCR Detection System (Bio-Rad, Hercules, CA, USA) and SYBR® Premix Ex TaqTM II (TaKaRa Bio). The following sequences of forward and reverse primer pairs were used: 5′-TCCATTATCTTCTGTTTGCTTTGC-3′ and 5′-TTCTGTAATGTGTTTATGATTCCTCCAT-3′ for hBD3 [19], 5′-AGCAATTGGTGGTGGATG-3′ and 5′-ACTCTTTGCAAAAGTTGTC-3′ for hEGF, 5′-GAGAGGAGAACTGCCAGAA-3′ and 5′-GTAGCATTTATGGAGAGTG-3′ for hEGFR, and 5′-AAGGTCGGAGTCAACGGATT-3′ and 5′-GCAGTGAGGGTCTCTCTCCT-3′ for Glyceraldehyde-3-phosphate dehydrogenase (GAPDH). GAPDH was used to normalize the detected gene expression and the fold change of experimental samples was estimated when untreated or control samples were set to 1.
2.5. Western Blot Analysis
Cells were lysed with 2X Laemmli sample buffer and boiled at 100 °C for 5 min. The supernatants were collected with centrifugation (12,000 rpm, 10 min) at 4 °C. Samples were separated on 10% (w/v) or 12% (w/v) sodium dodecyl sulfate-polyacrylamide gel electrophoresis (SDS-PAGE) in Tris-Glycine buffer (25 mM Tris, 250 mM Glycine, 0.1% SDS) or 16%/6 M urea separating gel in Tricine-SDS-PAGE buffer. Proteins were transferred onto polyvinylidene fluoride (PVDF) membranes (Millipore, Burlington, MA, USA) overnight (40 V and 4 °C). The membranes were treated with blocking buffer (20 mM Tris-HCl, 150 mM NaCl, 0.05% Tween-20, 5% BSA) for 2 h at room temperature (RT) and washed three times with TBS-T (20 mM Tris-HCl, 150 mM NaCl, 0.05% Tween-20). Then, the membranes were treated with primary antibodies such as anti-phospho-ERK1/2 (#9101), antiphospho-p38 (#9211), anti-phospho-SPAK/JNK (#9251), anti-phospho-AKT (#9271), antiphospho-JAK1 (#3331), anti-phospho-JAK2 (#3771) and anti-phospho-NF-κB p65 (#3033) antibodies (Cell Signaling Technology, Danvers, MA, USA), β-actin (sc-47778), or hBD 3 (sc-59495) antibodies (Santa Cruz Biotechnology, Dallas, TX, USA) for 2 h at RT. After washing three times with TBS-T buffer, the membranes were treated with secondary antibodies such as HRP-conjugated anti-rabbit (sc-2357), anti-goat (sc-2354) (Santa Cruz Biotechnology) or anti-mouse (#31430) (Invitrogen, Waltham, CA, USA) for 2 h at RT. After washing three times with TBS-T buffer, the membranes were treated with enhanced chemiluminescence (ECL) solutions (Thermo Fisher Scientific, Waltham, MA, USA) and exposed to X-ray film. The expression of β-actin was used as a loading control.
2.6. Enzyme-Linked Immunosorbent Assay (ELISA)
The quantities of IFN-γ were measured in the supernatant of HaCaT cells treated with KG lysates for indicated times. The quantity of IFN-γ was determined by Human IFN-γ DuoSet ELISA (R&D Systems, Minneapolis, MN, USA) according to the manufacturer’s instructions. Absorbance was measured using a microplate reader (Eppendorf BioPhotometer, Hamburg, Germany) at a test wavelength of 450 nm and a reference wavelength of 550 nm.
2.7. Preparation of S. aureus
S. aureus (ATCC 25923) was streaked onto brain heart infusion medium (BHI; HiMedia Laboratories, India) and grown overnight at 37 °C. A single colony of S. aureus was inoculated into 10 mL of BHI medium for overnight shaking cultivation at 37 °C. The logarithmic phase of S. aureus was obtained after 3 h subculture from the overnight culture. Bacterial concentrations were estimated by measuring the absorbance at 600 nm. Cells were centrifuged three times at 5000 rpm for 8 min, and the bacterial pellets were re-suspended in Dulbecco’s phosphate-buffered saline (DPBS), and diluted in DPBS to a concentration of 109 CFU/mL and conveniently diluted to different working concentrations. The bacterial count was confirmed by serial dilution and plating of the suspension.
2.8. Internalization Assay
HaCaT cells were seeded onto 6-well plates and cultured overnight. After stabilization, cells were washed three times with 1 mL DPBS and the medium was replaced with antibiotic-free DMEM, and samples were pretreated for the indicated times. S. aureus was added and treated for 6 h. Then, cells were washed three times with 1 mL DPBS and replaced with fresh DMEM medium containing 50 μg/mL gentamicin (WELGENE) to remove extracellular S. aureus. After 24 h incubation, cells were washed three times with DPBS and detached from the plate with trypsin-EDTA. The cells were then lysed with a hypertonic buffer (20 mM Tris; pH 7.5, with 5 mM MgCl2, 5 mM CaCl2, 1 mM DTT, 1 mM EDTA, and 0.1% Triton X-100) and serial dilutions were plated on a BHI agar plate. Plates were incubated overnight at 37 °C and CFU counted.
2.9. Preparation of Conditioned Media (CM)
HaCaT cells were seeded onto 6-well plates and cultured overnight. After stabilization, HaCaT cells were washed three times with 1 mL DPBS and the medium was replaced with antibiotic-free DMEM. The CM was harvested after treating cells with KG lysates for 72 h. The 1 mL KG lysates treated-CM (KG CM) were incubated with 10 μg of hBD 3 antibody (AF4435, R&D Systems) overnight at 4 °C with rotation. SureBeads™ Protein G Magnetic Beads (Bio-Rad) (20 μL) were added and further incubated for 3 h at 4 °C with rotation. Magnetic beads were centrifuged 2500 rpm for 1 min, and supernatants collected for hBD3 excluded CM.
2.10. Statistical Analysis
All of the experiments were performed at least three times. The data shown were the representative results of the means ± SD. Statistical analyses were conducted with an unpaired two-tailed t test, one-way ANOVA followed by Tukey’s honestly significant difference (HSD) post hoc test * p < 0.05, ** p < 0.01, *** p < 0.001, **** p < 0.0001 were interpreted as statistically significant. GraphPad Prism 5 software was used for the analysis (GraphPad Software Inc., San Diego, CA, USA).
3. Results
3.1. L. plantarum KG Lysates Significantly Increased hBD3 Production in HaCaT Cells
KG lysates significantly increased hBD3 mRNA expression from HaCaT cells after 48 h treatment. It further increased by 460-fold at 72 h. On the other hand, K8 lysates increased hBD3 mRNA by 50-fold. Data shows that KG lysates were 9-fold more inducible in hBD3 mRNA expression than K8 lysates (Figure 1A). Protein level of hBD3 was increased by 24 h stimulation of KG lysates and more hBD3 protein was noted at 72 h (Figure 1B). Next, the expression pattern according to concentration was observed. Both KG lysates and K8 lysates significantly increased hBD3 with 109 CFU/mL bacterial lysates (Figure 1C). The protein level of hBD3 was also dramatically increased by 109 CFU/mL KG lysates (Figure 1D). Viability assay was performed to determine whether KG lysates affect HaCaT viability. When cells were treated with 109 CFU/mL KG lysates for 72 h, viability was slightly reduced compared to the control, but there were no significant differences (Figure 1E).
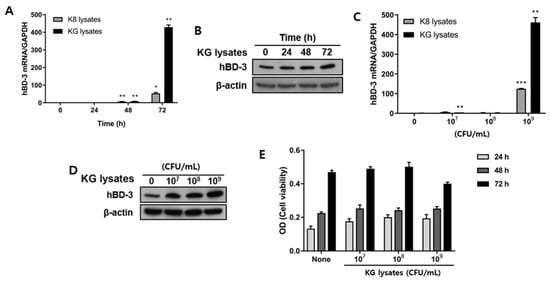
Figure 1.
L. plantarum KG lysates increased hBD3 expression in HaCaT cells. HaCaT cells were treated with the indicated doses of K8 lysates and KG lysates for the indicated times. (A,C) hBD3 mRNA level was examined by real-time PCR. (B,D) hBD3 protein level was examined by Western blot. (E) Cell viability assay was performed with HaCaT cells treated with KG lysates. * p < 0.05, ** p < 0.01, and *** p < 0.001.
3.2. L. plantarum KG Lysates Inhibited S. aureus Internalization into HaCaT Cells via hBD3 Induction
The effect of S. aureus on hBD3 expression was examined in HaCaT cells. Although it was less than in KG lysates, S. aureus induced hBD3 mRNA expression in HaCaT cells. Furthermore, S. aureus increased hBD3 mRNA expression more in cells pretreated with K8 lysates or KG lysates, suggesting that K8 lysates and KG lysates have a synergistic effect on the expression of hBD3 against pathogen infection (Figure 2A). Although K8 lysates also increased hBD3 mRNA expression, it did not effectively inhibit intracellular infection of S. aureus. On the other hand, KG lysates significantly inhibited intracellular infection of S. aureus (Figure 2B). When cells were treated with the indicated dose of KG lysates, intracellular S. aureus was decreased in a dose-dependent manner (Figure 2C). To examine whether the inhibition of S. aureus infection in KG lysate-treated cells was due to hBD3 induction, conditioned media (CM) excluding hBD3 was prepared. The treatment of normal CM containing hBD3 inhibited intracellular infection of S. aureus, but inhibition did not occur in cells treated with CM excluding hBD3, suggesting that hBD3 affected S. aureus infection (Figure 2D).
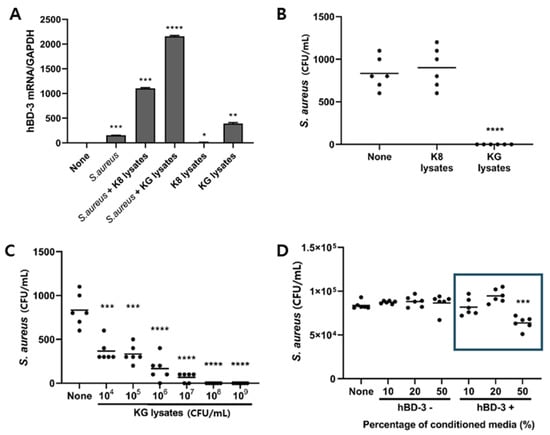
Figure 2.
L. plantarum KG lysates inhibited S. aureus internalization by HaCaT cells. (A,B) HaCaT cells were treated with 1 × 109 cells/mL bacterial lysates for 72 h and infected with 1 × 106 cells/mL S. aureus for 6 h. After removal of extracellular S. aureus, cells were further incubated for 24 h. (A) hBD3 mRNA level was examined by real-time PCR. (B) Intracellular S. aureus was counted by spreading of cell lysates. (C) HaCaT cells were treated with the indicated dose of KG lysates for 72 h. After S. aureus infection (as described above), intracellular S. aureus was counted. (D) HaCaT cells were treated with hBD3+ and hBD3− CM for 72 h and the number of S. aureus was measured through the above method. * p < 0.05, ** p < 0.01, *** p < 0.001, and **** p < 0.0001.
3.3. JAK-STAT Signaling Pathway Was Involved in hBD3 Induction in HaCaT Cells
When HaCaT cells were treated with KG lysates, no significant variation in signaling molecules was observed within 120 min. Rather, p38 and JNK showed a tendency to slightly decrease on phosphorylation after 60 min (Figure 3A). A change in signaling molecules was observed by extending the time to 72 h. Most signaling was down-regulated over time, but the phosphorylation of JAK 1 and JAK 2 was increased significantly (Figure 3B). In order to investigate the role of JAK1 and JAK2 in hBD3 expression, cells were pretreated with inhibitors and then KG lysates were treated. Ruxolitinib, an inhibitor of JAK1 and JAK2, decreased hBD3 mRNA expression in a dose-dependent manner (Figure 3C). Fludarabine, a STAT1 inhibitor, also decreased hBD3 mRNA level (Figure 3D). These data suggest that JAK1/2 and STAT1 are involved in the hBD3 expression in response to KG lysates in HaCaT cells. The JAK-STAT pathway is activated by IFN-γ. Thus, we examined the effect of IFN-γ on hBD3 production in HaCaT cells. When HaCaT cells were treated with IFN-γ, hBD3 mRNA was significantly increased (Figure 3E). We also observed that IFN-γ was increased by KG lysates in these cells (Figure 3F). These results suggest that IFN-γ produced in response to KG lysates stimulated the JAK-STAT pathway.
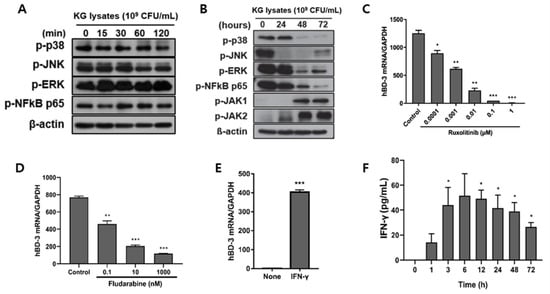
Figure 3.
The JAK-STAT1 pathway was involved in KG lysate-mediated hBD3 induction. (A) The phosphorylation of signaling molecules were detected by Western blot after HaCaT cells were treated with 1 × 109 cells/mL KG lysates for the indicated times. (B) HaCaT cells were treated with 1 × 109 cells/mL KG lysates for 24, 48, and 72 h and the variation of signaling molecules was examined by Western blot. HaCaT cells were treated with ruxolitinib (C) and fludarabine (D) prior to KG lysate treatment. hBD3 mRNA level was examined by real-time PCR. (E) HaCaT cells were treated with 10 ng/mL recombinant IFN-γ and hBD3 mRNA was examined by real-time PCR. (F) HaCaT cells were treated with 1 × 109 cells/mL KG lysates and IFN-γ production from culture supernatants was examined by ELISA. * p < 0.05, ** p < 0.01, and *** p < 0.001.
3.4. L. plantarum KG Lysates Increased EGF and EGFR Expression in HaCaT Cells
Although IFN-γ was detected in HaCaT cells treated with KG lysates, the protein level was too low to regulate hBD3 production. Thus, we suspected an alternative pathway and examined the state of EGF and EGFR mRNA levels in HaCaT cells. EFG mRNA was increased from 1 h after stimulation of KG lysates and peaked at 24 h (Figure 4A). EGFR increased significantly from 5 min of KG lysate treatment and maintained until 72 h (Figure 4B). To investigate the role of these genes in hBD3 expression, HaCaT cells were treated with EGFR inhibitor afatinib prior to KG lysates. hBD3 mRNA level was decreased in a dose-dependent manner (Figure 4C). These data suggest that KG lysates increased hBD3 expression by inducing EGF and EGFR expression.
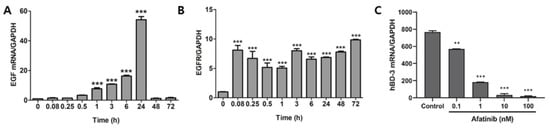
Figure 4.
EGFR is involved in KG lysate-mediated hBD3 induction. HaCaT cells were treated with 1 × 109 cells/mL KG lysates for the indicated times and EGF mRNA (A) and EGFR mRNA (B) was examined by real-time PCR. (C) HaCaT cells were pretreated with afatinib and re-treated with KG lysates. hBD3 mRNA level was examined by real-time PCR. ** p < 0.01 and *** p < 0.001.
4. Discussion
Keratinocytes, located in the outer layer of human skin, play an essential role in the skin defense against infections. In addition to creating a physical barrier between the environment and the inside of the body, keratinocytes exert a strong immune function against pathogenic infections. Skin keratinocytes express proinflammatory cytokines and antimicrobial peptides including hBD2, hBD3, and LL37 that affect skin immune system [20]. Because immune defenses of the skin can be increased by probiotics, the role of probiotics in disease management and immune regulation has recently received renewed attention from society. The skin immunoregulatory effect of L. plantarum KG was improved by fermentation with ginseng extracts. KG lysates suppressed S. aureus infection by increasing hBD3 expression in HaCaT cells. HBD3 expression in the skin affects skin inflammation, bacterial infections, and wound healing [21]. Indeed, hBD3 was significantly increased in HaCaT cells by KG lysates. Interestingly, hBD3 expression was more inducible in cells pretreated with KG lysates followed by infection of S. aureus. The KG lysates also showed a reduced S. aureus infection rate compared to the K8 lysates. Treatment with only K8 lysates did not affect, suggesting that it is a characteristic of KG lysates. It is unclear whether the inhibition of S. aureus intracellular infection was caused by antimicrobial activity of hBD3 or if it affects the mechanism used by S. aureus for intracellular infection. However, there was a study stating that hBD3 kills S. aureus [1]. In our research, S. aureus was cultured in CM prepared from HaCaT cells treated with KG lysates, but it did not significantly affect the viability of S. aureus (data not shown). S. aureus uses the Fibronectin α5β1 integrin pathway to enter host cells [22]. The interaction between S. aureus and its host cell receptors such as TLR2, CD36, and CD14 may also be important. Considering the anti-inflammatory and resistance-inducing efficacy of L. plantarum lysates, intracellular infection may be suppressed by reducing the expression of genes related to S. aureus infection.
The mechanism of inducing the expression of hBD3 in skin keratinocytes is not well known, but EGFR may be involved [23]. EGFR induces interferon regulatory factor 1 (IRF-1) expression through STAT1 activation [24,25]. We observed that KG lysates increased IRF-1 from 5 min treatment and peaked at 6 h (data not shown). IRF-1 acts as a transcriptional activator and regulates expression of type I interferon genes [26]. When cells were treated with EGFR inhibitor, IRF-1 expression was inhibited (data not shown). However, in this study, we have not yet confirmed directly that IRF-1 is involved in hBD3. Further studies on the relationship between IRF-1 and hBD3 expression are needed. The expression of hBD3 in HaCaT cells increased significantly at 72 h. This is thought to be because the activity of JAT-STAT1 occurs late. The expression of EGFR increased starting 5 min after treatment with KG lysates, but EGF increased from 1 h post-treatment and reached a peak at 24 h. In other words, we speculate that time is necessary for the signal initiated from EGF-EGFR to activate IFN-γ signaling to activate JAK-STAT1. In addition, the relationship between EGFR signaling and IFN-γ-mediated JAK/STAT activation will require further clarification.
The advantages of probiotics lysates are (i) higher immunity boosting efficacy than whole bacteria, (ii) commercialization through various formulations, and (iii) higher stability in the distribution process compared to live bacteria. Lysates of L. plantarum increased anti-inflammatory activity according to the number of crushing. L. plantarum lysates have anti-inflammatory activity, and suppresses transient immune function such as excessive expression of TNF-α induced by LPS in the immune cells. This anti-inflammatory effect tended to increase as the number of disruptions of L. plantarum increased [16]. In another experiment, molecular weight analysis of the index component and MS/MS analysis were performed to investigate the structure of the component having anti-inflammatory activity. The molecular weight of the index component was confirmed to be 1080.4 and 1094.5 through mass spectrometry, and it was identified as a type of LTA, a glycolipid containing a hexasaccharide through MS/MS analysis [27].
In conclusion, we isolated a new strain of L. plantarum KG from kimchi, and KG lysates strongly induced the expression of hBD3 in human keratinocytes after fermentation with ginseng extracts. hBD3 induced by KG lysates was shown to inhibit S. aureus infection into human keratinocytes, suggesting that KG lysates can improve control of the skin microbiome without the need of antibiotics. Since the KG lysate is in the form of parabiotics or postbiotics, it can be provided in various formulations for the development of food and pharmaceutical materials to control bacterial infection.
Author Contributions
H.K. and D.-K.C. conceived, planned the experiments; K.-O.J. and H.K. wrote the manuscript; K.-O.J., C.Y. and H.K. carried out the experiments; H.K. and D.-K.C. contributed to the interpretation of the results; D.-K.C. supervised the project. All authors have read and agreed to the published version of the manuscript.
Funding
This research was financially supported by the Ministry of Trade, Industry, and Energy (MOTIE), Korea, under the “Infrastructure Support Program for Industry Innovation” (reference number P114000049) supervised by the Korea Institute for Advancement of Technology (KIAT).
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
The data presented in this study are available in the article.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Hoover, D.M.; Wu, Z.; Tucker, K.; Lu, W.; Lubkowski, J. Antimicrobial characterization of human beta-defensin 3 derivatives. Antimicrob. Agents Chemother. 2003, 47, 2804–2809. [Google Scholar] [CrossRef] [PubMed]
- Tewary, P.; Li, Z.; de la Rosa, G.; Chen, Q.; Oppenheim, J.; Yang, D. Human Beta Defensin 3 (HBD3) induces migration and activation of antigen presenting cells and acts as an immune enhancer. FASEB J. 2008, 22, 673. [Google Scholar] [CrossRef]
- Bayer, A.; Lammel, J.; Tohidnezhad, M.; Lippross, S.; Behrendt, P.; Klüter, T.; Pufe, T.; Cremer, J.; Jahr, H.; Rademacher, F.; et al. The Antimicrobial Peptide Human Beta-Defensin-3 Is Induced by Platelet-Released Growth Factors in Primary Keratinocytes. Mediat. Inflamm. 2017, 2017, 6157491. [Google Scholar] [CrossRef]
- Sørensen, O.E.; Cowland, J.B.; Theilgaard-Mönch, K.; Liu, L.; Ganz, T.; Borregaard, N. Wound healing and expression of antimicrobial peptides/polypeptides in human keratinocytes, a consequence of common growth factors. J. Immunol. 2003, 170, 5583–5589. [Google Scholar] [CrossRef]
- Kiehne, K.; Fincke, A.; Brunke, G.; Lange, T.; Fölsch, U.R.; Herzig, K.H. Antimicrobial peptides in chronic anal fistula epithelium. Scand. J. Gastroenterol. 2007, 42, 1063–1069. [Google Scholar] [CrossRef] [PubMed]
- Sørensen, O.E.; Thapa, D.R.; Rosenthal, A.; Liu, L.; Roberts, A.A.; Ganz, T. Differential regulation of beta-defensin expression in human skin by microbial stimuli. J. Immunol. 2005, 174, 4870–4879. [Google Scholar] [CrossRef]
- Wehkamp, J.; Harder, J.; Wehkamp, K.; Wehkamp-von Meissner, B.; Schlee, M.; Enders, C.; Sonnenborn, U.; Nuding, S.; Bengmark, S.; Fellermann, K.; et al. NF-kappaB- and AP-1-mediated induction of human beta defensin-2 in intestinal epithelial cells by Escherichia coli Nissle 1917: A novel effect of a probiotic bacterium. Infect. Immun. 2004, 72, 5750–5758. [Google Scholar] [CrossRef]
- Paolillo, R.; Romano Carratelli, C.; Sorrentino, S.; Mazzola, N.; Rizzo, A. Immunomodulatory effects of Lactobacillus plantarum on human colon cancer cells. Int. Immunopharmacol. 2009, 9, 1265–1271. [Google Scholar] [CrossRef]
- Kobatake, E.; Kabuki, T. S-Layer Protein of Lactobacillus helveticus SBT2171 Promotes Human β-Defensin 2 Expression via TLR2-JNK Signaling. Front. Microbiol. 2019, 10, 2414. [Google Scholar] [CrossRef]
- Liu, A.Y.; Destoumieux, D.; Wong, A.V.; Park, C.H.; Valore, E.V.; Liu, L.; Ganz, T. Human beta-defensin-2 production in keratinocytes is regulated by interleukin-1, bacteria, and the state of differentiation. J. invest. Dermatol. 2002, 118, 275–281. [Google Scholar] [CrossRef]
- Lee, D.E.; Huh, C.S.; Ra, J.; Choi, I.D.; Jeong, J.W.; Kim, S.H.; Ryu, J.H.; Seo, Y.K.; Koh, J.S.; Lee, J.H.; et al. Clinical Evidence of Effects of Lactobacillus plantarum HY7714 on Skin Aging: A Randomized, Double Blind, Placebo-Controlled Study. J. Microbiol. Biotechnol. 2015, 25, 2160–2168. [Google Scholar] [CrossRef] [PubMed]
- Nam, B.; Kim, S.A.; Park, S.D.; Kim, H.J.; Kim, J.S.; Bae, C.H.; Kim, J.Y.; Nam, W.; Lee, J.L.; Sim, J.H. Regulatory effects of Lactobacillus plantarum HY7714 on skin health by improving intestinal condition. PLoS ONE 2020, 15, e0231268. [Google Scholar] [CrossRef] [PubMed]
- Tsai, W.H.; Chou, C.H.; Chiang, Y.J.; Lin, C.G.; Lee, C.H. Regulatory effects of Lactobacillus plantarum-GMNL6 on human skin health by improving skin microbiome. Int. J. Med. Sci. 2021, 18, 1114–1120. [Google Scholar] [CrossRef] [PubMed]
- Kim, J.Y.; Kim, H.; Jung, B.J.; Kim, N.R.; Park, J.E.; Chung, D.K. Lipoteichoic acid isolated from Lactobacillus plantarum suppresses LPS-mediated atherosclerotic plaque inflammation. Mol. Cells 2013, 35, 115–124. [Google Scholar] [CrossRef] [PubMed]
- Hong, Y.F.; Kim, H.; Kim, H.R.; Gim, M.G.; Chung, D.K. Different immune regulatory potential of Lactobacillus plantarum and Lactobacillus sakei isolated from Kimchi. J. Microbiol. Biotechnol. 2014, 24, 1629–1635. [Google Scholar] [CrossRef]
- Kim, H.; Kim, H.R.; Kim, N.R.; Jeong, B.J.; Lee, J.S.; Jang, S.; Chung, D.K. Oral administration of Lactobacillus plantarum lysates attenuates the development of atopic dermatitis lesions in mouse models. J. Microbiol. 2015, 53, 47–52. [Google Scholar] [CrossRef] [PubMed]
- Lee, Y.D.; Hong, Y.F.; Jeon, B.; Jung, B.J.; Chung, D.K.; Kim, H. Differential Cytokine Regulatory Effect of Three Lactobacillus Strains Isolated from Fermented Foods. J. Microbiol. Biotechnol. 2016, 26, 1517–1526. [Google Scholar] [CrossRef] [PubMed]
- Ahn, J.E.; Kim, H.; Chung, D.K. Lipoteichoic Acid Isolated from Lactobacillus plantarum Maintains Inflammatory Homeostasis through Regulation of Th1- and Th2-Induced Cytokines. J. Microbiol. Biotechnol. 2019, 29, 151–159. [Google Scholar] [CrossRef]
- Gambichler, T.; Skrygan, M.; Tomi, N.S.; Othlinghaus, N.; Brockmeyer, N.H.; Altmeyer, P.; Kreuter, A. Differential mRNA expression of antimicrobial peptides and proteins in atopic dermatitis as compared to psoriasis vulgaris and healthy skin. Int. Arch. Allergy Immunol. 2008, 147, 17–24. [Google Scholar] [CrossRef]
- Wang, J.N.; Li, M. The Immune Function of Keratinocytes in Anti-Pathogen Infection in the Skin. Int. J. Dermatol. Venereol. 2020, 3, 231–238. [Google Scholar] [CrossRef]
- Schittek, B.; Paulmann, M.; Senyürek, I.; Steffen, H. The role of antimicrobial peptides in human skin and in skin infectious diseases. Infect. Disord. Drug Targets 2008, 8, 135–143. [Google Scholar] [CrossRef] [PubMed]
- Josse, J.; Laurent, F.; Diot, A. Staphylococcal Adhesion and Host Cell Invasion: Fibronectin-Binding and Other Mechanisms. Front Microbiol. 2017, 8, 2433. [Google Scholar] [CrossRef] [PubMed]
- Shuyi, Y.; Feng, W.; Jing, T.; Hongzhang, H.; Haiyan, W.; Pingping, M.; Liwu, Z.; Zwahlen, R.A.; Hongyu, Y. Human beta-defensin-3 (hBD-3) upregulated by LPS via epidermal growth factor receptor (EGFR) signaling pathways to enhance lymphatic invasion of oral squamous cell carcinoma. Oral Surg. Oral Med. Oral Pathol. Oral Radiol. Endod. 2011, 112, 616–625. [Google Scholar] [CrossRef] [PubMed]
- Andersen, P.; Pedersen, M.W.; Woetmann, A.; Villingshøj, M.; Stockhausen, M.T.; Odum, N.; Poulsen, H.S. EGFR induces expression of IRF-1 via STAT1 and STAT3 activation leading to growth arrest of human cancer cells. Int. J. Cancer 2008, 122, 342–349. [Google Scholar] [CrossRef]
- Lulli, D.; Carbone, M.; Pastore, S. Epidermal growth factor receptor inhibitors trigger a type I interferon response in human skin. Oncotarget 2016, 7, 47777–47793. [Google Scholar] [CrossRef]
- Feng, H.; Zhang, Y.B.; Gui, J.F.; Lemon, S.M.; Yamane, D. Interferon regulatory factor 1 (IRF1) and anti-pathogen innate immune responses. PLoS Pathog. 2021, 17, e1009220. [Google Scholar] [CrossRef]
- Jang, K.S.; Baik, J.E.; Han, S.H.; Chung, D.K.; Kim, B.G. Multi-spectrometric analyses of lipoteichoic acids isolated from Lactobacillus plantarum. Biochem. Biophys. Res. Commun. 2011, 407, 823–830. [Google Scholar] [CrossRef]
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).