A Dual Stable Isotope Approach Unravels Common Climate Signals and Species-Specific Responses to Environmental Change Stored in Multi-Century Tree-Ring Series from the Tibetan Plateau



 , and
, and 
Abstract
1. Introduction
- (i)
- deciphering and explaining common and contrasting long-term isotopic variations of δ13C and δ18O between two tree species from high-altitude, climate-sensitive tree-stands;
- (ii)
- assessing the added value for an analysis of δ13C and δ18O from two local tree species; and
- (iii)
- testing if common/differing signals of the proxy series from one site can be used to better assess, decipher, and explain diverging responses of tree species to recent environmental change.
2. Materials and Methods
2.1. Study Site
2.2. Sampling, Dating and Sample Preparation
2.3. Evaluation of δ18OTRC and δ13CTRC
2.4. Examination of Isotope–Climate Relationships
2.5. δ13CTRC Detrending and iWUE Calculation
3. Results and Discussion
3.1. Species Specific Characteristics of δ18OTRC
3.2. Species Specific Characteristics of δ13CTRC
3.3. Climate–δ18OTRC Relationships
3.4. Climate–δ13CTRC Relationships
3.5. Implications of Species-Specific Proxy–Climate Responses of Multi-Centennial Isotope Series
3.6. Long-Term Trends in Carbon Isotope Ratios and their Implications for Recording Climate Signals
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Yao, T.; Thompson, L.; Yang, W.; Yu, W.; Gao, Y.; Guo, X.; Yang, X.; Duan, K.; Zhao, H.; Xu, B.; et al. Different glacier status with atmospheric circulations in Tibetan plateau and surroundings. Nat. Clim. Chang. 2012, 2, 663–667. [Google Scholar] [CrossRef]
- Immerzeel, W.; van Beek, L.; Bierkens, M. Climate Change will affect the Asia Water Towers. Science 2010, 328, 1382–1385. [Google Scholar] [CrossRef] [PubMed]
- Royden, L.H.; Burchfiel, B.C.; van der Hilst, R.R. The geological evolution of the Tibetan plateau. Science 2008, 321, 1054–1058. [Google Scholar] [CrossRef]
- Böhner, J. General climatic controls and topoclimatic variations in Central and High Asia. Boreas 2006, 35, 279–295. [Google Scholar] [CrossRef]
- Yang, K.; Wu, H.; Qin, J.; Lin, C.; Tang, W.; Chen, Y. Recent climate changes over the Tibetan plateau and their impacts on energy and water cycle: A review. Glob. Planet. Chang. 2014, 112, 79–91. [Google Scholar] [CrossRef]
- Yang, K.; Ye, B.; Zhou, D.; Wu, B.; Foken, T.; Qin, J.; Zhou, Z. Response of the hydrological cycle to recent climate changes in the Tibetan plateau. Clim. Chang. 2011, 109, 517–534. [Google Scholar] [CrossRef]
- Cheng, G.; Wu, T. Responses of permafrost to climate change and their environmental significance, Qinghai-Tibet plateau. J. Geophys. Res. 2007, 112, F02S03. [Google Scholar] [CrossRef]
- Kang, S.; Xu, Y.; You, Q.; Flügel, W.A.; Pepin, N.; Yao, T. Review of climate and cryospheric change in the Tibetan plateau. Environ. Res. Lett. 2010, 5, 01510. [Google Scholar] [CrossRef]
- Yang, B.; He, M.; Shishov, V.; Tychkov, I.; Vaganov, E.; Rossi, S.; Ljungqvist, F.C.; Bräuning, A.; Grießinger, J. New perspective on spring vegetation phenology and global climate change based on Tibetan Plateau tree-ring data. Proc. Natl. Acad. Sci. USA 2017, 114, 6966–6971. [Google Scholar] [CrossRef] [PubMed]
- Shi, C.; Daux, V.; Zhang, Q.-B.; Risi, C.; Hou, S.-G.; Stievenard, M.; Pierre, M.; Li, Z.; Masson-Delmotte, V. Reconstruction of southeast Tibetan plateau summer climate using tree ring 18O: Moisture variability over the past two centuries. Clim. Past 2012, 8, 205–213. [Google Scholar] [CrossRef]
- Grießinger, J.; Bräuning, A.; Helle, G.; Thomas, A.; Schleser, G.H. Late Holocene Asian summer monsoon variability reflected by δ18O in tree-rings from Tibetan junipers. Geophys. Res. Lett. 2011, 38, L03701. [Google Scholar] [CrossRef]
- Grießinger, J.; Bräuning, A.; Helle, G.; Hochreuther, P.; Schleser, G.H. Late Holocene relative humidity history on the southeastern Tibetan plateau inferred from a tree-ring δ18O record: Recent decrease and conditions during the last 1500 years. Quat. Int. 2017, 430, 52–59. [Google Scholar] [CrossRef]
- Wernicke, J.; Hochreuther, P.; Grießinger, J.; Zhu, H.; Wang, L.; Bräuning, A. Multi-century humidity reconstructions from the southeastern Tibetan Plateau inferred from tree-ring δ18O. Glob. Planet. Chang. 2016, 149. [Google Scholar] [CrossRef]
- Wernicke, J.; Grießinger, J.; Hochreutherm, P.; Bräuning, A. Variability of summer humidity during the past 800 years on the eastern Tibetan Plateau inferred from δ18O of tree-ring cellulose. Clim. Past 2015, 11, 327–337. [Google Scholar] [CrossRef]
- Hochreuther, P.; Wernicke, J.; Grießinger, J.; Mölg, T.; Zhu, H.; Wang, L.; Bräuning, A. Influence of the Indian Ocean Dipole on tree-ring δ18O of monsoonal Southeast Tibet. Clim. Chang. 2016, 137, 217–230. [Google Scholar] [CrossRef]
- Xu, G.; Liu, X.; Trouet, V.; Treydte, K.; Wu, G.; Chen, T.; Sun, W.; An, W.; Wang, W.; Zeng, X.; et al. Regional shifts (1710–2019) in East Central Asia and linkages with atmospheric circulation recorded in tree-ring δ18O. Clim. Dyn. 2018. [Google Scholar] [CrossRef]
- Liu, X.; Xu, G.; Grießinger, J.; An, W.; Wang, W.; Zeng, X.; Wu, G.; Qin, D. A shift in cloud cover over the southeastern Tibetan plateau since 1600, evidence from regional tree-ring 18O and its linkages to tropical oceans. Quat. Sci. Rev. 2014, 88, 55–68. [Google Scholar] [CrossRef]
- McCarroll, D.; Gagen, M.H.; Loader, N.J.; Robertson, I.; Anchukaitis, K.J.; Los, S.; Young, G.H.F.; Jalkanen, R.; Kirchhefer, A.; Waterhouse, J.S. Correction of tree ring stable carbon isotope chronologies for changes in the carbon dioxide content of the atmosphere. Geochim. Cosmochim. Acta 2009, 73, 1539–1547. [Google Scholar] [CrossRef]
- Loader, N.J.; McCarroll, D.; Gagen, M.; Robertson, I.; Jalkanen, R. Extracting climatic informations from stable isotopes in tree rings. In Stable Isotopes as Indicators of Ecological Change; Dawson, T.E., Siegwolf, R.T.W., Eds.; Elsevier: Amsterdam, The Netherlands, 2007; pp. 27–48. [Google Scholar]
- Xu, G.; Liu, X.; Qin, D.; Chen, T.; Sun, W.; An, W.; Wang, W.; Wu, G.; Zeng, X.; Ren, J. Drought history inferred from tree ring 13C and 18O in the central Tianshan Mountains of China and linkage with the North Atlantic Oscillation. Theor. Appl. Climatol. 2014, 116, 385–401. [Google Scholar] [CrossRef]
- Xu, G.; Liu, X.; Belmecheri, S.; Chen, T.; Wu, G.; Wang, B.; Zeng, X.; Wang, W. Disentangling Contributions of CO2 Concentration and Climate to Changes in intrinsic water-use efficiency in the arid boreal forest in China’s Altay Mountains. Forests 2018, 9, 642. [Google Scholar] [CrossRef]
- Xu, G.; Liu, X.; Sun, W.; Chen, T.; Zhang, X.; Zeng, X.; Wu, G.; Wang, W.; Qin, D. Application and verification of simultaneous determination of cellulose δ13C and δ18O in Picea shrenkiana tree rings from northwestern China using the high-temperature pyrolysis method. J. Arid Land 2018, 10, 864–876. [Google Scholar] [CrossRef]
- An, W.; Liu, X.; Leavitt, S.W.; Ren, J.; Sun, W.; Wang, W.; Wang, Y.; Xu, G.; Chen, T.; Qin, D. Specific climatic signals recorded in earlywood and latewood 18O of tree rings in southwestern China. Tellus B 2012, 64, 18703. [Google Scholar] [CrossRef]
- Hartl-Meier, C.; Zang, C.; Büntgen, U.; Esper, J.; Rothe, A.; Göttlein, A.; Dirnböck, T.; Treydte, K. Uniform climate sensitivity in tree-ring stable isotopes across species and sites in a mid-latitude temperate forest. Tree Physiol. 2014, 35, 4–15. [Google Scholar] [CrossRef] [PubMed]
- Fu, P.; Grießinger, J.; Gebrekirstos, A.; Fan, Z.-X.; Bräuning, A. Earlywood and Latewood Stable Carbon and Oxygen Isotope Variations in Two Pine Species in Southwestern China during the Recent Decades. Front. Plant Sci. 2017, 7, 2050. [Google Scholar] [CrossRef] [PubMed]
- Zeng, X.; Liu, X.; Treydte, K.; Evans, M.N.; Wang, W.; An, W.; Sun, W.; Xu, G.; Wu, G.; Zhang, X. Climate signals in tree-ring δ18O and δ13C from southeastern Tibet: Insights from observations and forward modelling of intra- to interdecadal variability. New Phytol. 2017, 216, 1104–1118. [Google Scholar] [CrossRef] [PubMed]
- He, M.; Yang, B.; Bräuning, A.; Rossi, S.; Ljungqvist, F.C.; Shishov, V.; Grießinger, J.; Wang, J.; Liu, J.; Qin, C. Recent advances in dendroclimatology in China. Earth-Sci. Rev. 2019. [Google Scholar] [CrossRef]
- Huang, R.; Zhu, H.; Liu, X.; Liang, E.; Grießinger, J.; Wu, G.; Li, X.; Bräuning, A. Does increasing intrinsic water use efficiency (iWUE) stimulate tree growth at natural timberline on the southeastern Tibetan plateau? Glob. Planet. Chang. 2017, 148, 217–226. [Google Scholar] [CrossRef]
- Bräuning, A. Climate history of the Tibetan plateau during the last 1000 years derived from a network of Juniper chronologies. Dendrochronologia 2001, 19, 127–137. [Google Scholar]
- Zhu, H.; Shao, X.; Yin, J.; Huang, L. Early summer temperature reconstruction in the eastern Tibetan plateau since AD 1440 using tree-ring width of Sabina tibetica. Theor. Appl. Clim. 2011, 106, 45–53. [Google Scholar] [CrossRef]
- Miehe, G.; Miehe, S.; Vogel, J.; La, D. Highest tree-line in the northern hemisphere found in southern Tibet. Mt. Res. Dev. 2007, 27, 169–173. [Google Scholar] [CrossRef]
- Leavitt, S. Tree-ring C-H-O isotope variability and sampling. Sci. Total Environ. 2010, 408, 5244–5253. [Google Scholar] [CrossRef]
- McCarroll, D.; Loader, N.J. Stable isotopes in tree rings. Quat. Sci. Rev. 2004, 23, 771–801. [Google Scholar] [CrossRef]
- Wieloch, T.; Helle, G.; Heinrich, I.; Voight, M.; Schyma, P. A novel device for batch-wise isolation of a-cellulose from small-amount wholewood samples. Dendrochronologia 2011, 29, 115–117. [Google Scholar] [CrossRef]
- Laumer, W.; Andreau, L.; Helle, G.; Schleser, G.H.; Wieloch, T.; Wissel, H. A novel approach for the homogenization of cellulose to use micro-amounts for stable isotope analyses. Rapid Commun. Mass Spectrom. 2009, 23, 1934–1940. [Google Scholar] [CrossRef] [PubMed]
- Harris, I.; Jones, P.D.; Osborn, T.J.; Lister, D.H. Updated high-resolution grids of monthly climatic observations—The CRU TS 3.10 dataset. J. Clim. 2014, 34, 623–642. [Google Scholar] [CrossRef]
- R Core Team. A Language and Environment for Statistical Computing. R Foundation for Statistical Computing. 2013. Available online: www.R-project.org/ (accessed on 1 March 2019).
- MacFarling Meure, C.; Etheridge, D.; Trudinger, C.; Steele, P.; Langenfelds, R.; Van Ommen, T.; Smith, A.; Elkins, J. Law Dome CO2, CH4 and N2O ice core records extended to 2000 years BP. Geophys. Res. Lett. 2006, 33, L14810. [Google Scholar] [CrossRef]
- Ehleringer, J.R.; Hall, A.E.; Farquhar, G.D. Stable Isotopes and Plant Water Relations; Elsevier: Dordrecht, The Netherlands, 1993. [Google Scholar]
- Farquhar, G.D.; O’Leary, M.; Berry, J. On the relationship between carbon isotope discrimination and the intercellular carbon dioxide concentration in leaves. Funct. Plant Biol. 1982, 9, 121–137. [Google Scholar] [CrossRef]
- Dansgaard, W. Stable isotopes in precipitation. Tellus 1964, 4, 436–468. [Google Scholar]
- Barbour, M. Stable oxygen isotope composition of pant tissue: A review. Funct. Plant Biol. 2007, 34, 83–94. [Google Scholar] [CrossRef]
- Gessler, A.; Ferrio, J.P.; Hommel, R.; Treydte, K.; Werner, R.A.; Monson, R.K. Stable isotopes in tree rings e toward a mechanistic understanding of isotope fractionation and mixing processes from the leaves to the wood. Tree Physiol. 2014, 34, 796–818. [Google Scholar] [CrossRef] [PubMed]
- Huang, R.; Zhu, H.; Liang, E.; Grießinger, J.; Wernicke, J.; Yu, W.; Hochreuther, P.; Risi, C.; Zeng, Y.; Fremme, A.; et al. Temperature signals in tree-ring oxygen isotope series from the northern slope of the Himalaya. Earth Planet. Sci. Lett. 2018, 506, 455–465. [Google Scholar] [CrossRef]
- Wang, W.; Liu, X.; Shao, X.; Leavitt, S.; Xu, G.; An, W.; Qin, D. A 200 year temperature record from tree ring δ13C at the Qaidam Basin of the Tibetan Plateau after identifying the optimum method to correct for changing atmospheric CO2 and δ13C. J. Geophys. Res. Biogeosci. 2011, 116, G04022. [Google Scholar] [CrossRef]
- Liu, Y.; Wang, Y.; Li, Q.; Song, H.; Linderholm, H.W.; Leavitt, S.W.; Wang, R.; An, Z. Tree-ring stable carbon isotope-based May–July temperature reconstruction over Nanwutai, China, for the past century and its record of 20th century warming. Quat. Sci. Rev. 2014, 93, 67–76. [Google Scholar] [CrossRef]
- Liu, Y.; Ta, W.; Li, Q.; Song, H.; Sun, C.; Cai, Q.; Liu, H.; Wang, L.; Hu, S.; Sun, J.; et al. Tree-ring stable carbon isotope-based April–June relative humidity reconstruction since AD 1648 in Mt. Tianmu, China. Clim. Dyn. 2017, 50, 1733–1745. [Google Scholar] [CrossRef]
- Liu, Y.; Ma, L.; Leavitt, S.W.; Cai, Q.; Liu, W. A preliminary seasonal precipitation reconstruction from tree-ring stable carbon isotopes at Mt. Helan, China, since AD 1804. Glob. Planet. Chang. 2004, 41, 229–239. [Google Scholar] [CrossRef]
- Körner, C.; Paulsen, J. A world wide study of high altitude treeline temperatures. J. Biogeogr. 2004, 31, 713–732. [Google Scholar] [CrossRef]
- Hyvönen, R.; Agren, G.I.; Linder, S.; Persson, T.; Cotrufo, M.F.; Ekblad, A.; Freeman, M.; Grelle, A.; Janssens, I.A.; Jarvis, P.G.; et al. The likely impact of elevated CO2, nitrogen deposition, increased temperature and management on carbon sequestration in temperate and boreal forest ecosystems: A literature review. New Phytol. 2007, 173, 463–480. [Google Scholar] [CrossRef] [PubMed]
- Drake, B.G.; Gonzalez-Meier, M.A.; Long, S.P. More efficient plants: A consequence of rising atmospheric CO2? Annu. Rev. Plant Physiol. Plant Mol. Biol. 1997, 48, 609–639. [Google Scholar] [CrossRef]
- Battipaglia, G.; Saurer, M.; Cherubini, P.; Calfapietra, C.; McCarthy, H.R.; Norby, R.J.; Cotrufo, M.F. Elevated CO2 increases tree-level intrinsic water-use efficiency. Insights from carbon and oxygen isotopes analyses in tree rings across three forest FACE sites. New Phytol. 2013, 197, 544–554. [Google Scholar] [CrossRef] [PubMed]
- Giammarchi, F.; Cherubini, P.; Pretzsch, H.; Tonon, G. The increase of atmospheric CO2 affects growth potential and intrinsic water-use efficiency of Norway spruce forests: Insights from a multiple-stable isotope analysis in tree rings of two Alpine chronosequences. Trees 2017, 31, 503–515. [Google Scholar] [CrossRef]
- Wullschleger, S.D.; Tschaplinski, T.J.; Norby, R.J. Plant water relations at elevated CO2—Implications for water-limited environments. Plant Cell Environ. 2002, 25, 319–331. [Google Scholar] [CrossRef] [PubMed]
- Cao, L.; Yan, Z.; Zhao, P.; Zhu, Y.; Yu, Y.; Tang, G.; Jones, P. Climatic warming in China during 1901–2015 based on an extended dataset of instrumental temperature records. Environ. Res. Lett. 2012, 12, 064005. [Google Scholar] [CrossRef]
- Qin, J.; Yang, K.; Liang, S.; Guo, X. The altitudinal dependence of recent rapid warming over the Tibetan plateau. Clim. Chang. 2009, 97, 321–327. [Google Scholar] [CrossRef]
- Altieri, S.; Mereu, S.; Cherubini, P.; Castaldi, S.; Sirignano, C.; Lubritto, C.; Battipaglia, G. Tree-ring carbon and oxygen isotopes indicate different water-use strategies in three Mediterranean shrubs at Capo caccia (Sardinia, Italy). Trees 2015, 29, 1593–1603. [Google Scholar] [CrossRef]
- Xu, G.; Liu, X.; Qin, D.; Chen, T.; An, W.; Wang, W.; Wu, G.; Zeng, X.; Ren, J. Climate warming and increasing atmospheric CO2 have contributed to increased water use efficiency on the northeastern Tibetan plateau since 1850. Trees Struct. Funct. 2013, 27, 465–475. [Google Scholar] [CrossRef]
- Hietz, P.; Wanek, W.; Duenisch, O. Long-term trends in cellulose delta C-13 and water-use efficiency of tropical Cedrela and Swietenia from Brazil. Tree Physiol. 2005, 25, 745–752. [Google Scholar] [CrossRef] [PubMed]
- Wils, T.H.G.; Roberston, I.; Woodborne, S.; Hall, G.; Kaprowski, M.; Eshetu, Z. Anthropogenic forcing increases the water-use efficiency of African trees. J. Quat. Sci. 2016, 31, 386–390. [Google Scholar] [CrossRef]
- Urrutia-Jalabert, R.; Malhi, Y.; Barichivivh, J.; Lara, A.; Delgado-Huertas, A.; Rodríguez, C.G.; Cuq, E. Increased water use efficiency but contrasting tree growth patterns in Fitzroya cupressoides forests of southern Chile during recent decades. J. Geophys. Res. Biogeosciences 2015, 120, 2505–2524. [Google Scholar] [CrossRef]
- Keenan, T.F.; Hollinger, D.Y.; Bohrer, G.; Dragoni, D.; Munger, J.W.; Peter, H. Richardson, Increase in forest water-use efficiency as atmospheric carbon dioxide concentrations rise. Nature 2013, 499, 324–327. [Google Scholar] [CrossRef]
- Feng, X. Trends in intrinsic water-use efficiency of natural trees for the past 100-200 years: A response to atmospheric CO2 concentration. Geochim. Cosmochim. Acta 1999, 63, 1891–1903. [Google Scholar] [CrossRef]
- Penuelas, J.; Hunt, J.M.; Ogaya, R.; Jump, A.S. Twentieth century changes of tree ringδ13C at the southern range-edge of Fagus sylvatica: Increasing water use efficiency does not avoid the growth decline induced by warming at low altitudes. Glob. Chang. Biol. 2008, 14, 1076–1088. [Google Scholar] [CrossRef]
- Frank, D.; Poulter, B.; Saurer, M.; Esper, J.; Huntingford, C.; Helle, G.; Treydte, K.; Zimmermann, N.E.; Schleser, G.H.; Ahlström, A.; et al. Water-use eciency and transpiration across European forests during the Anthropocene. Nat. Clim. Chang. 2015, 5, 579–583. [Google Scholar] [CrossRef]
- Silva, L.C.R.; Anand, M.; Oliveira, J.M.; Pillar, V.D. Past century changes in Araucaria angustifolia (Bertol.) Kuntze water use efficiency and growth in forest and grassland ecosystems of southern Brazil: Implications for forest expansion. Glob. Chang. Biol. 2009, 15, 2387–2396. [Google Scholar] [CrossRef]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
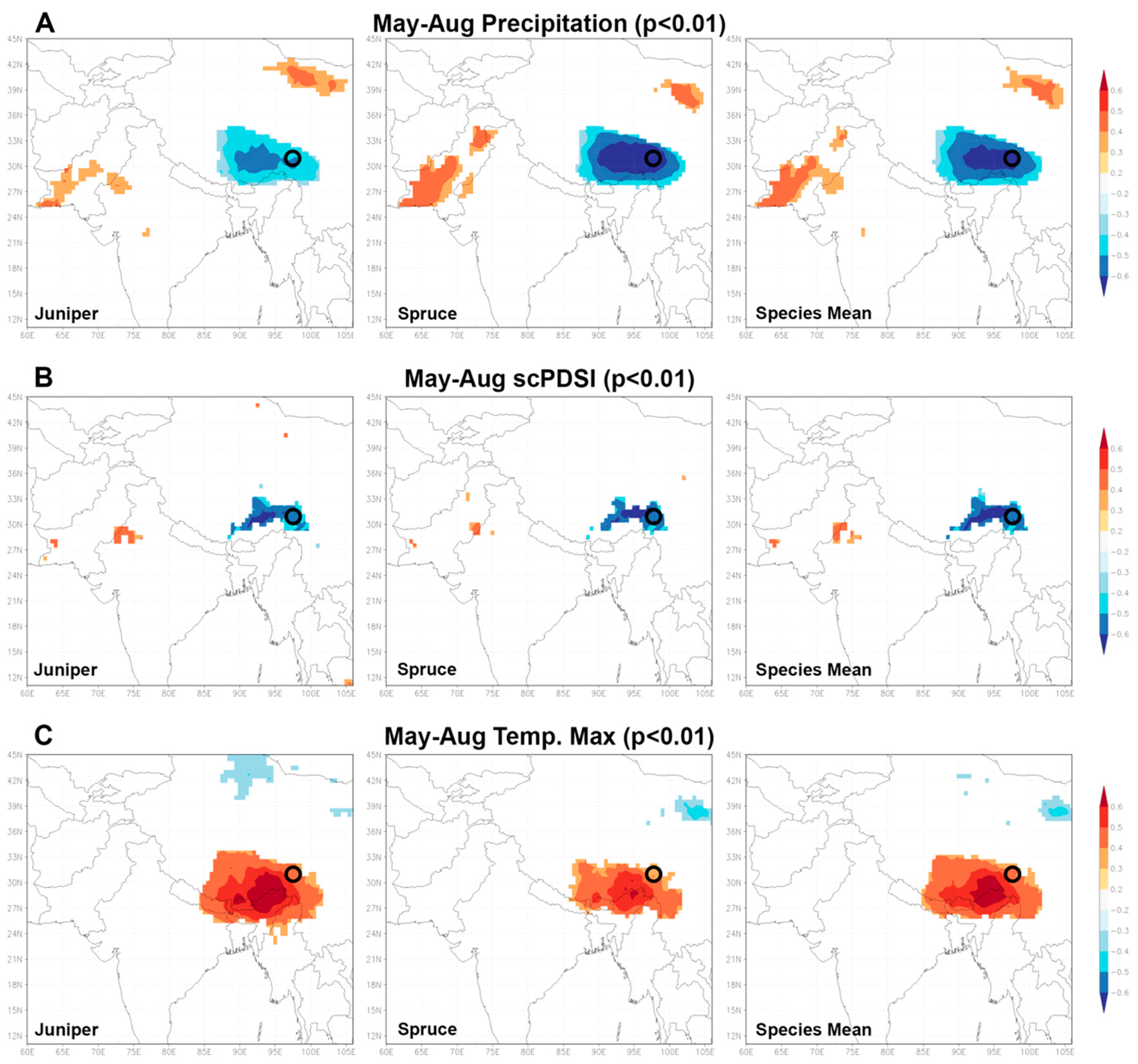{kind=link}
{kind=link}
| Species | Altitude (m asl.) | Chronology Time Span | Mean δ18OTRC | Mean δ13CTRC Corrected | SD | AC1 |
|---|---|---|---|---|---|---|
| Juniperus tibetica | 4600 m | 489–2011 AD | 23.33‰ | −20.46‰ | 1.2 (δ18O) | 0.61 (δ18O) |
| 0.5 (δ13C corr.) | 0.74 (δ13C corr.) | |||||
| Picea balfouriana | 4400 m | 1685–2007 AD | 22.98‰ | −20.67‰ | 1.4 (δ18O) | 0.26 (δ18O) |
| 0.9 (δ13C corr.) | 0.81 (δ13C corr.) |
| Stable Isotope Series | Annual | Low-Freq. (11-year FFT) | Low-Freq. (21-year FFT) |
|---|---|---|---|
| δ18OTRC | r = 0.60 | r = 0.73 | r = 0.75 |
| GLK = 70% | GLK = 91% | GLK = 78% | |
| δ13CTRC raw | r = 0.86 | r = 0.95 | r = 0.96 |
| GLK = 51% | GLK = 63% | GLK = 63% | |
| δ13CTRC CO2-corr. | r = 0.44 | r = 0.64 | r = 0.69 |
| GLK = 52% | GLK = 64% | GLK = 65% |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Grießinger, J.; Bräuning, A.; Helle, G.; Schleser, G.H.; Hochreuther, P.; Meier, W.J.-H.; Zhu, H. A Dual Stable Isotope Approach Unravels Common Climate Signals and Species-Specific Responses to Environmental Change Stored in Multi-Century Tree-Ring Series from the Tibetan Plateau. Geosciences 2019, 9, 151. https://doi.org/10.3390/geosciences9040151
Grießinger J, Bräuning A, Helle G, Schleser GH, Hochreuther P, Meier WJ-H, Zhu H. A Dual Stable Isotope Approach Unravels Common Climate Signals and Species-Specific Responses to Environmental Change Stored in Multi-Century Tree-Ring Series from the Tibetan Plateau. Geosciences. 2019; 9(4):151. https://doi.org/10.3390/geosciences9040151
Chicago/Turabian StyleGrießinger, Jussi, Achim Bräuning, Gerhard Helle, Gerhard Hans Schleser, Philipp Hochreuther, Wolfgang Jens-Henrik Meier, and Haifeng Zhu. 2019. "A Dual Stable Isotope Approach Unravels Common Climate Signals and Species-Specific Responses to Environmental Change Stored in Multi-Century Tree-Ring Series from the Tibetan Plateau" Geosciences 9, no. 4: 151. https://doi.org/10.3390/geosciences9040151
APA StyleGrießinger, J., Bräuning, A., Helle, G., Schleser, G. H., Hochreuther, P., Meier, W. J.-H., & Zhu, H. (2019). A Dual Stable Isotope Approach Unravels Common Climate Signals and Species-Specific Responses to Environmental Change Stored in Multi-Century Tree-Ring Series from the Tibetan Plateau. Geosciences, 9(4), 151. https://doi.org/10.3390/geosciences9040151