Small Ruminants: Farmers’ Hope in a World Threatened by Water Scarcity

 , and
, and
Simple Summary
Abstract
1. Introduction
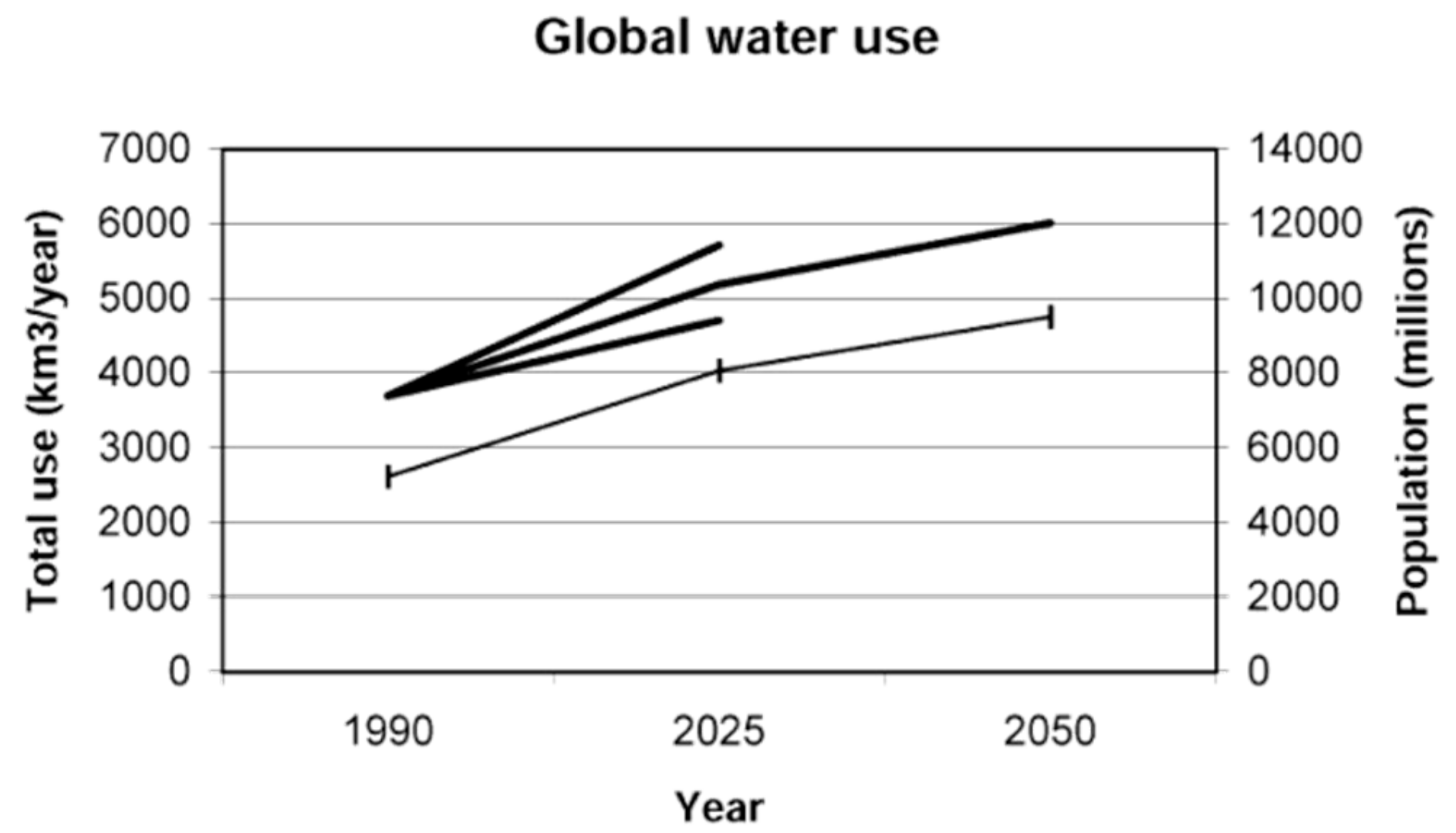
2. Global Water Resources and Use
3.Small Ruminant Distribution
4. Water Requirements and Metabolism
5. Adaptive Responses of Small Ruminants to Water Shortages and Deprivation
5.1. Effect on Metabolic, Rectal Temperature, Respiratory and Pulse Rate
5.2. Drinking Behaviour, Body Weight and Feed Intake
5.3. Effect on Nitrogen Balance and Digestibility
5.4. Effect on Blood Metabolites
5.5. Effect on Physiological Status
5.5.1. Effect on Reproductive Traits and Hormones
5.5.2. Lactation
5.6. Immune Response and Drug Pharmacokinetics
6. Modulating Genes in Small Ruminants Found in Dry Areas and Future Research Gap
7. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- United Nations. Sustainable Development Goals, Clean Water and Sanitation: Why It Matters; United Nations: San Francisco, CA, USA, 2015. [Google Scholar]
- Naqvi, S.M.K.; Kumar, D.; Kalyan, D.; Sejian, V. Climate change and water availability for livestock: Impact on both quality and quantity. In Climate Change Impact on Livestock: Adaptation and Mitigation; Sejian, V., Gaughan, J., Baumgard, L., Prasad, C.S., Eds.; Springer-Verlag GMbH Publisher: New Delhi, India, 2015; pp. 81–96. [Google Scholar]
- IPCC (Intergovernmental Panel on Climate Change). Synthesis Report. In Contribution of Working Groups I, II and II to the Fourth Assessment Report of the Intergovernmental Panel on Climate Change; Pachauri, R.K., Reisinger, A., Eds.; IPCC: Geneva, Switzerland, 2007; p. 104. [Google Scholar]
- Damerau, K.; van Vliet, O.P.R.; Patt, A.G. Direct impacts of alternative energy scenarios on water demand in the Middle East and North Africa Clim. Change 2015, 130, 171–183. [Google Scholar]
- Schlosser, C.A.; Strzepek, K.; Gao, X.; Fant, C.; Blanc, É.; Paltsev, S.; Jacoby, H.; Reilly, J.; Gueneau, A. The future of global water stress: An integrated assessment. Earth’s Future 2014, 2, 341–361. [Google Scholar] [CrossRef]
- United Nation. Sustainable Development Goals. Revision of World Population Prospects; United Nations: San Francisco, CA, USA, 2017. [Google Scholar]
- Nejad, G.J.; Lohakare, J.D.; West, J.W.; Sung, K.I. Effects of water restriction after feeding during heat stress on nutrient digestibility, nitrogen balance, blood profile and characteristics in Corriedale ewes. Anim. Feed Sci. Technol. 2014, 193, 1–8. [Google Scholar] [CrossRef]
- Abdelatif, A.M.; Elsayed, S.A.; Hassan, Y.M. Effect of state of hydration on body weight, blood constituents and urine excretion in Nubian Goats (Capra hircus). World J. Agric. Sci. 2010, 6, 178–188. [Google Scholar]
- Peacock, C. Goats-A pathway to poverty. Small Rum. Res. 2005, 20, 179–186. [Google Scholar] [CrossRef]
- Ben Salem, H.; Smith, T. Feeding strategies to increase small ruminant production in dry environment. Small Rum. Res. 2008, 77, 174–194. [Google Scholar] [CrossRef]
- Seo, S.N.; Mendelsohn, R. Climate Change Adaptation in Africa: A Microeconomic Analysis of Livestock Choice; CEEPA Discussion Paper, No. 19; Centre for Environmental Economics and Policy in Africa, University of Pretoria: Pretoria, South Africa, 2006. [Google Scholar]
- Silanikove, N. The physiological basis of adaptation in goats to harsh environments. Small Rum. Res. 2000, 35, 181–193. [Google Scholar] [CrossRef]
- Jaber, L.S.; Habre, A.; Rawda, N.; AbiSaid, M.; Barbour, E.K.; Hamadeh, S.K. The effect of water restriction on certain physiological parameters in Awassi sheep. Small Rum. Res. 2004, 54, 115–120. [Google Scholar] [CrossRef]
- Casamassima, D.; Pizzo, R.; Palazzo, M.; Alessandro, A.G.D.; Martemucci, G. Effect of water restriction on productive performance and blood parameters in comisana sheep reared under intensive condition. Small Rum. Res. 2008, 78, 169–175. [Google Scholar] [CrossRef]
- Doreau, M.; Corson, M.S.; Wiedemann, S.G. Water use by livestock: A global perspective for a regional issue. Anim. Front. 2012, 2, 9–16. [Google Scholar] [CrossRef]
- Jaber, L.; Chedid, M.; Hamadeh, S. Water stress in small ruminants. In Responses of Organisms to Water Stress; Akinci, S., Ed.; InTech: Rijeka, Croatia, 2013; pp. 115–149. [Google Scholar]
- Chedid, M.; Jaber, L.S.; Giger-Reverdin, S.; Duvaux-Ponter, C.; Hamadeh, S.K. Review: Water stress in sheep raised under arid conditions. Can. J. Anim. Sci. 2014, 94, 243–257. [Google Scholar] [CrossRef]
- Shkolnik, A.; Maltz, E.; Choshniak, I. The role of the ruminants digestive tract as a water reservoir. In Digestive Physiology and Metabolism in Ruminants; Rockebusch, Y., Thiven, P., Eds.; MTP Press: Lancaster, UK, 1980; pp. 731–741. [Google Scholar]
- Shiklomanov, I.A. Water in Crisis: A Guide to the World’s Fresh Water Resources; Gleick, P.H., Ed.; Oxford University Press: New York, NY, USA, 1993; pp. 13–24. [Google Scholar]
- Postel, S.L.; Daily, G.C.; Ehrlich, P.R. Human appropriation of renewable fresh water. Science 1996, 271, 785–788. [Google Scholar] [CrossRef]
- Kundzewicz, Z.W.; Döll, P. Will groundwater ease freshwater stress under climate change? Hydrol. Sci. J. J. Des. Sci. Hydrol. 2009, 54, 665–675. [Google Scholar] [CrossRef]
- Misra, A.K. Climate change and challenges of water and food scarcity. Int. J. Sustain. Built Dev. 2014, 3, 153–165. [Google Scholar] [CrossRef]
- Gosain, A.K.; Sandhya, R.; Debajit, B. Climate change impact assessment on the hydrology of Indian river basins. Curr. Sci. 2006, 90, 346–353. [Google Scholar]
- Fujimori, S.; Hanasaki, N.; Masui, T. Projections of industrial water withdrawal under shared socioeconomic pathways and climate mitigation scenarios. Sustain. Sci. 2017, 12, 275–292. [Google Scholar] [CrossRef] [PubMed]
- Arnell, N.W. Climate change and global water resources. Glob. Environ. Chang. 1999, 9, 31–49. [Google Scholar] [CrossRef]
- Schlink, A.C.; Nguyen, M.L.; Viljoen, G.J. Water requirements for livestock production: A global perspective. Rev. Sci. Tech. Off. Int. Epiz. 2010, 29, 603–619. [Google Scholar] [CrossRef]
- Marino, R.; Atzori, A.S.; D’Andrea, M.; Iovane, G.; Trabalza-Marinucci, M.; Rinaldi, L. Climate change: Production performance, health issues, greenhouse gas emissions and mitigation strategies in sheep and goat farming. Small Rum. Res. 2016, 135, 50–59. [Google Scholar] [CrossRef]
- Food and Agriculture Organization of the United Nations [FAO]. Statistical Year Book; Food and Agriculture Organization of the United Nations: Rome, Italy, 2016; Volume 1. [Google Scholar]
- Skapetas, B.; Bampidis, V. Goat production in the World: Present situation and trends. Livest. Res. Rural Dev. 2016, 28, 200. Available online: http://www.lrrd.org/lrrd28/11/skap28200.html (accessed on 17 December 2018).
- Foresight. The Future of Food and Farming; Final Project Report; The Government Office for Science: London, UK, 2011. Available online: http://www.bis.gov.uk/assets/foresight/docs/food-and-farming/11-546-future-of-food-and-farmingreport.Pdf (accessed on 15 January 2019).
- Scherf, B.D. (Ed.) World Watch List of Domestic Animal Diversity, 3rd ed.; Food and Agriculture Organization of the United Nations: Rome, Italy, 2000; p. 726. [Google Scholar]
- Iniguez, L. The challenges of research and development of small ruminant production in dry areas. Small Rum. Res. 2011, 98, 12–20. [Google Scholar] [CrossRef]
- National Research Council (NRC). Nutrient Requirements’ of Small Ruminants: Sheep, Goats, Cervids, and New World Camelids; NRC: Washington, DC, USA, 2007; 384p. [Google Scholar]
- Meyer, U.; Everinghoff, M.; Gadeken, D.; Flachowsky, G. Investigations on water intake of lactating dairy cows. Livest. Prod. Sci. 2004, 90, 117–121. [Google Scholar] [CrossRef]
- Beede, D.K. What will our ruminants drink? Anim. Front. 2012, 2, 36–43. [Google Scholar] [CrossRef]
- Esminger, M.E.; Oldfield, J.L.; Heinemann, J.J. Feeds and Nutrition, 2nd ed.; Esminger Publishing: Clovis, CA, USA, 1990; 1552p. [Google Scholar]
- Maynard, L.A.; Loosli, J.K.; Hintz, H.F.; Warner, R.G. Animal Nutrition; Tata McGraw-Hill Publishing: New Delhi, India, 1981. [Google Scholar]
- Araujo, G.G.L.; Voltolini, T.V.; Chizzotti, M.L.; Turco, S.H.N.; de Carvalho, F.F.R. Water and small ruminant production. Revista Brasileira de Zootecnia 2010, 39, 326–336. [Google Scholar] [CrossRef]
- Ratnakaran, A.P.; Sejian, V.; Jose, V.S.; Vaswani, S.; Bagath, M.; Krishnan, G.; Beena, P.; Devi, I.; Varma, G.; Bhatta, R. Behavioral Responses to Livestock Adaptation to Heat Stress Challenges. Asian J. Anim. Sci. 2017, 11, 1–13. [Google Scholar]
- Kerr, S.R. Livestock Heat Stress: Recognition, Response, and Prevention; Extension Fact Sheet FS157E, Washington, US Pullman; Washington State University: Pullman, WA, USA, 2015. [Google Scholar]
- Choshniak, I.; Ben-Kohav, N.; Taylor, C.R.; Robertshaw, D.; Barnes, R.J.; Dobson, A.; Belkin, V.; Shkolnik, A. Metabolic adaptations for desert survival in the Bedouin goat. Am. J. Physiol. 1995, 268, 1101–1110. [Google Scholar] [CrossRef]
- Rahardja, D.P.; Toleng, A.L.; Lestari, V.S. Thermoregulation and water balance in fat-tailed sheep and Kacang goat under sunlight exposure and water restriction in a hot and dry area. Animal 2011, 5, 1587–1593. [Google Scholar] [CrossRef]
- Nejad, J.G.; Sung, K. Behavioral and physiological changes during heat stress in Corriedale ewes exposed to water deprivation. J. Anim. Sci. Technol. 2017, 59, 13. [Google Scholar] [CrossRef]
- Davis, J.R.; DeNardo, D.F. The urinary bladder as a physiological reservoir that moderates dehydration in a large desert lizard, the Gila monster Helodermasuspectum. J. Exp. Biol. 2007, 210, 1472–1480. [Google Scholar] [CrossRef]
- Mckinley, M.J.; Martelli, D.; Pennington, G.L.; Trevaks, D.; McAllen, R.M. Integrating Competing Demands of Osmoregulatory and Thermoregulatory Homeostasis. Am. Phys. Soc. 2007, 33, 170–181. [Google Scholar] [CrossRef]
- Mitchell, D.; Maloney, S.K.; Jessen, C.; Laburn, H.P.; Kamerman, P.R.; Mitchell, G.; Fuller, A. Adaptive heterothermy and selective brain cooling in arid zone mammals. Comp. Biochem. Physiol. B 2002, 131, 571–585. [Google Scholar] [CrossRef]
- Jessen, C.; Dmi’el, R.; Choshniak, I.; Ezra, D.; Kuhnen, G. Effects of dehydration and rehydration on body temperatures in the black Bedouin goat. Pflüg. Arch. Eur. J. Physiol. 1998, 436, 659–666. [Google Scholar] [CrossRef] [PubMed]
- Hamadeh, S.K.; Rawda, N.; Jaber, L.S.; Habre, A.; Abi Said, M.; Barbour, E.K. Physiological responses to water restriction in dry and lactating Awassi ewes. Livest. Sci. 2006, 101, 101–109. [Google Scholar] [CrossRef]
- Casamassima, D.; Vizzarri, F.; Nardola, M.; Palazzo, M. Effect of water restriction on various physiological variables on intensively reared Lacauna ewes. Vet. Med. 2016, 61, 623–634. [Google Scholar] [CrossRef]
- Alamer, M. Effect of water restriction on thermoregulation and some biochemical constituents in lactating Aardi goats during hot weather conditions. J. Basic Appl. Sci. Res. 2010, 11, 189–205. [Google Scholar]
- Fuller, A.; Meyer, L.C.R.; Mitchell, D.; Maloney, S.K. Dehydration increases the magnitude of selective brain cooling independently of core temperature in sheep. Am. J. Physiol. Regul. Integr. Comp. Physiol. 2007, 293, R438–R446. [Google Scholar] [CrossRef] [PubMed]
- Alamer, M. Effect of water restriction on lactation performance of Aardi goats under heat stress conditions. Small Rum. Res. 2009, 84, 76–81. [Google Scholar] [CrossRef]
- Alamer, M. Physiological responses of Saudi Arabia indigenous goats to water deprivation. Small Rum Res. 2006, 63, 100–109. [Google Scholar] [CrossRef]
- Ismail, E.; Hassan, G.A.; Abo-Elezz, Z.; AbdElLatiff, H. Physiological responses of Barki and Suffolk sheep and their crossbreeds to dehydration. Egypt. J. Anim. Prod. 1996, 33, 89–101. [Google Scholar]
- Das, S.K.; Kumar, P.; Singh, D.; Singh, K. Effect of water restriction on thermoregulatory responses in bucks. Indian J. Anim. Sci. 1994, 64, 899–901. [Google Scholar]
- Mengistu, U.; Dahlborn, K.; Olsson, K. Effects of intermittent watering on water balance and feed intake in male Ethiopian Somali goats. Small Rum. Res. 2007, 67, 45–54. [Google Scholar] [CrossRef]
- Choshniak, I.; Wittenberg, C.; Rosenfeld, J.; Shkolnik, A. Rapid rehydration and kidney function in the black Bedouin goat. Physiol. Zool. 1984, 57, 573–579. [Google Scholar] [CrossRef]
- Abdelatif, A.M.; Ahmed, M.M.M. Water restriction, thermoregulation, blood constituents and endocrine responses in Sudanese desert sheep. J. Arid Environ. 1994, 26, 171–180. [Google Scholar] [CrossRef]
- Adogla-Bessa, T.; Aganga, A.A. Responses of Tswana goats to various lengths of water deprivation. S. Afr. J. Anim. Sci. 2000, 30, 87–91. [Google Scholar] [CrossRef]
- Li, B.T.; Christopherson, R.J.; Cosgrove, S.J. Effect of water restriction and environmental temperatures on metabolic rate and physiological parameters in sheep. Can. J. Anim. Sci. 2000, 80, 97–104. [Google Scholar] [CrossRef]
- Alamer, M.; Al-hozab, A. Effect of water deprivation and season on feed intake, body weight and thermoregulation in Awassi and Nadji sheep breeds in Saudi Arabia. J. Arid Environ. 2004, 59, 71–84. [Google Scholar] [CrossRef]
- El-Hadi, H.M. The effect of dehydration on Sudanese desert sheep and goats. J. Agric. Sci. 1986, 106, 17–20. [Google Scholar] [CrossRef]
- Parker, A.J.; Hamlin, G.P.; Coleman, C.J.; Fitzpatrick, L.A. Dehydration in stressed ruminants may be the result of a cortisol induced diuresis. J. Anim. Sci. 2003, 81, 512–519. [Google Scholar] [CrossRef]
- Kumar, D.; De, K.; Singh, A.K.; Kumar, K.; Sahoo, A.; Naqvi, S.M.K. Effect of water restriction on physiological responses and certain reproductive traits of Malpura ewes in a semiarid tropical environment. J. Vet. Behav. 2016, 12, 54–59. [Google Scholar] [CrossRef]
- Al-Ramamneh, D.; Riek, A.; Gerken, M. Effect of water restriction on drinking behaviour and water intake in German black-head mutton sheep and Boer goats. Animal 2012, 6, 173–178. [Google Scholar] [CrossRef]
- El Khashab, M.A.; Semaida, A.I.; Abd El-Ghany, M.A. Water restriction and its effect on blood hormones, minerals and metabolite in Baladii goats. Int. J. Curr. Microbiol. App. Sci. 2018, 7, 747–755. [Google Scholar] [CrossRef]
- Degen, A.A.; Kam, M. Body mass loss and body uid shifts during dehydration in Dorper sheep. J. Agric. Sci. 1992, 119, 419–422. [Google Scholar] [CrossRef]
- Choshniak, A.; Shkolnik, A. Rapid rehydration in the black Bedouin goat and role of rumen red blood cells fragility. Comp. Biochem. Physiol. 1997, 56A, 581–583. [Google Scholar] [CrossRef]
- Ikhatua, U.J.; Dede, T.I.; Apumami, A.S. Water and feed utilisation relationship in West African Dwarf (forest) goats of Southern Nigeria. Niger. J. Anim. Prod. 1992, 19, 120–124. [Google Scholar]
- Maloiy, G.M.O.; Kanui, T.I.; Towett, P.K.; Wambugu, S.N.; Miaron, J.O.; Wanyoike, M.M. Effects of dehydration and heat stress on food intake and dry matter digestibility in East African ruminants. Comp. Biochem. Physiol. 2008, 151, 185–190. [Google Scholar] [CrossRef] [PubMed]
- Mengistu, U.; Dahlborn, K.; Olsson, K. Effect of intermittent watering on dry matter intake and body weight of male Ethiopian Somali goats. J. Anim. Feed Sci. 2004, 13, 647–650. [Google Scholar] [CrossRef][Green Version]
- Hadjigeorgiou, I.; Dardamani, K.; Goulas, C.; Zervas, G. Effect of water availability on feed intake and digestibility in sheep. Small Rum. Res. 2000, 37, 147–150. [Google Scholar] [CrossRef]
- Hassan, G.E. Physiological responses of Anglo-Nubian and Baladi goats and their crossbreeds to water deprivation under subtropical conditions. Livest. Prod. Sci. 1989, 22, 295–304. [Google Scholar]
- El-Nouty, F.D.; Al-Haiderary, A.A.; Basmaeil, S.M. Physiological responses, feed intake, urine volume and serum osmolality of Aardi goats deprived of water during spring and summer. Aust. J. Anim. Sci. 1990, 3, 331–336. [Google Scholar] [CrossRef]
- Ayoub, M.A.; Saleh, A.A. A comparative physiological study between camels and goats during water deprivation. In Proceedings of the Third Annual Meeting for Animal Production under Arid Conditions; United Arab Emirates University: Al Ain, UAE, 1998; Volume 1, pp. 71–87. [Google Scholar]
- Mousa, H.M.; Ali, K.E.; Hume, I.D. Effect of water deprivation on urea metabolism in camels, desert sheep and desert goats fed dry desert grass. Comp. Biochem. Physiol. A Physiol. 1983, 74, 715–720. [Google Scholar] [CrossRef]
- Muna, M.M.A.; Ammar, I.E.S. Effects of water and feed restriction on body weight change and nitrogen balance in desert goats fed high and low quality forages. Small Rum. Res. 2001, 41, 19–27. [Google Scholar] [CrossRef]
- Osman, H.E.; Fadlalla, B. The effect of level of water intake on some aspects of digestion and nitrogen metabolism of desert sheep of the Sudan. J. Agric. Camb. 1974, 82, 61–69. [Google Scholar] [CrossRef]
- Silanikove, N. The struggle to maintain hydration and osmoregulation in animals experiencing severe dehydration and rapid rehydration: The story of ruminants. Exp. Physiol. 1994, 79, 281–300. [Google Scholar] [CrossRef] [PubMed]
- Thang, T.V.; Sunagawa, K.; Nagamine, I.; Kishi, T.; Ogura, G.A. Physiological stimulating factor of water intake during and after forage feeding in large-type goats. Asian Australas. J. Anim. Sci. 2012, 25, 502–514. [Google Scholar] [CrossRef] [PubMed]
- Macfarlane, W.V.; Howard, B. Comparative water and energy economy of wild and domestic mammals. Symp. Zool. Soc. Lond. 1972, 31, 261–296. [Google Scholar]
- Langhans, W.; Scharre, E.; Meyer, A.H. Changes in feeding behaviour and plasma vasopressin concentration during water deprivation in goats. J. Vet. Med. 1991, 38, 11–20. [Google Scholar] [CrossRef]
- Wittemberg, C.; Choshniak, I.; Shkolnik, A.; Thurau, K.; Rosenfeld, J. Effect of dehydration and rapid rehydration on renal function and plasma renin and aldosterone levels in the black Bedouin goat. Pflügers Arch. 1986, 406, 405–408. [Google Scholar] [CrossRef]
- Kramer, J.W. Normal haematology of cattle, sheep and goats. In Schalm’s Veterinary Haematology, 5th ed.; Feldman, B.F., Zinkl, J.G., Jain, N.C., Eds.; Lippincott Williams and Wilkins: Philadelphia, PA, USA, 2000; pp. 1075–1084. [Google Scholar]
- Igbokwe, I.O. Haemoconcentration in Yankasa sheep exposed to prolonged water deprivation. Small Rum. Res. 1993, 12, 99–105. [Google Scholar] [CrossRef]
- Baxmann, A.C.; Ahmed, M.S.; Marques, N.C.; Menon, V.B.; Pereira, A.B.; Kirsztajn, G.M.; Heilberg, I.P. Influence of Muscle Mass and Physical Activity on Serum and Urinary Creatinine and Serum Cystatin C. Clin. J. Am. Soc. Nephrol. 2008, 3, 348–355. [Google Scholar] [CrossRef]
- Ashour, G.; Benlemilh, S. Adaptation of Mediterranean breeds to heat stress and water deprivation. In Livestock Production and Climatic Uncertainty in the Mediterranean, Proceedings of the Joint ANPA-EAAPCIHEAM-FAO Symposium, Agadir, Morocco, 22–24 October 2000; Guessous, F., Rihani, N., Ilham, A., Eds.; EAAP Publication No. 94; Wageningen Pers: Wageningen, The Netherlands, 2000; pp. 63–74. [Google Scholar]
- Tasker, J.B. Fluids, electrolytes, and acid-base balance. In Clinical Biochemistry of Domestic Animals, 2nd ed.; Kaneko, J.J., Cornelius, C.E., Eds.; Academic Press Inc.: London, UK, 1971; pp. 65–75. [Google Scholar]
- Moeller, K.T.; Butler, M.W.; Denardo, D.F. The effect of hydration state and energy balance on innate immunity of a desert reptile. Front. Zool. 2013, 10, 23. [Google Scholar] [CrossRef]
- Schneider, E.G. In water deprivation osmolality become an important determination of aldosterone secretions. News. Physiol. Sci. 1990, 5, 197–201. [Google Scholar]
- Afsal, A.; Sejian, V.; Bagath, M.; Krishnan, G.; Devaraj, C.; Bhatta, R. Heat Stress and Livestock Adaptation: Neuro-endocrine Regulation. Int. J. Vet. Anim. Med. 2008, 1, 108. [Google Scholar]
- Naqvi, S.M.K.; Soren, N.M.; Karim, S.A. Effect of concentrate supplementation on performance, ovarian response, and some biochemical profile of Malpura ewes. Trop. Anim. Health Prod. 2011, 43, 905–913. [Google Scholar] [CrossRef]
- Koyuncu, M.; Canbolat, O. Effect of different dietary energy levels on the reproductive performance of Kivircik sheep under a semi-intensive system in the South-Marmara region of Turkey. J. Anim. Feed Sci. 2009, 18, 620–627. [Google Scholar] [CrossRef]
- Liker, S.; Gunes, S.; Murat, Y.; Funda, K.; Ahmet, C. The effects of body weight, body condition score, age, lactation, serum triglyceride, cholesterol and paraoxonase levels on pregnancy rate of Saanen goats in the breeding season. J. Anim. Vet. Adv. 2010, 9, 1848–1851. [Google Scholar]
- Sejian, V.; Maurya, V.P.; Naqvi, S.M.K. Effect of thermal, nutritional and combined (thermal and nutritional) stresses on growth and reproductive performance of Malpura ewes under semi-arid tropical environment. J. Anim. Physiol. Anim. Nutr. 2011, 95, 252–258. [Google Scholar] [CrossRef]
- Roth, Z.; Meidan, R.; Braw-Tal, R.; Wolfenson, D. Immediate and delayed effects of heat stress on follicular development and its association with plasma FSH and inhibin concentration in cows. J. Reprod. Fertil. 2000, 120, 83–90. [Google Scholar] [CrossRef]
- Kiyma, Z.; Alexander, B.M.; Van Kirk, E.A.; Murdoch, W.J.; Hallford, D.M.; Moss, G.E. Effect of feed restriction on reproductive and metabolic hormones in ewes. J. Anim. Sci. 2004, 82, 2548–2557. [Google Scholar] [CrossRef]
- Narayan, E.; Parisella, S. Influences of the stress endocrine system on the reproductive endocrine axis in sheep (Ovisaries). Ital. J. Anim. Sci. 2017, 16, 640–651. [Google Scholar] [CrossRef]
- Deck, C.A.; Honeycutt, J.L.; Cheung, E.; Reynolds, H.M.; Borski, R.J. Assessment of the functional role of leptin in energy homeostasis and the stress response in vertebrates. Front. Endocrinol. 2017, 8, 63. [Google Scholar] [CrossRef]
- Chilliard, Y.; Ferlay, A.; Faulconnier, Y.; Bonnet, M.; Rouel, J.; Bocquier, F. Adipose tissue metabolism and its role in adaptations to under-nutrition in ruminants. Proc. Nutr. Soc. 2000, 59, 127–134. [Google Scholar] [CrossRef]
- Kraut, J.A.; Madias, N.E. Lactic acidosis. N. Engl. J. Med. 2014, 371, 2309–2319. [Google Scholar] [CrossRef]
- Lozano, J.M.; Abecia, J.A.; Forcada, F.; Zarazaga, L.; Alfaro, B. Effect of undernutrition on the distribution of progesterone in the uterus of ewes during the luteal phase of the oestrous cycle. Theriogenology 1998, 49, 539–546. [Google Scholar] [CrossRef]
- Kaliber, M.; Koluman, N.; Silanikove, N. Physiological andbehavioural basis for the successful adaptation of goats to severe water restriction under hot environmental conditions. Animal 2015, 10, 82–88. [Google Scholar] [CrossRef]
- Roumelioti, M.E.; Glew, R.H.; Khitan, Z.J.; Rondon-Berrios, H.; Argyropoulos, C.P.; Malhotra, D.; Raj, D.S.; Agaba, E.I.; Rohrscheib, M.; Murata, G.H.; et al. Fluid balance concepts in medicine: Principles and practice. World J. Nephrol. 2018, 7, 1–28. [Google Scholar] [CrossRef]
- Aganga, A.A. Water utilization by sheep and goats in Northern Nigeria. World Anim. Rev. 1992, 73, 9–14. [Google Scholar]
- Alamer, M. The role of prolactin in thermoregulation and water balance during heat stress in domestic ruminants. Asian J. Anim. Vet. Adv. 2011, 6, 1153–1169. [Google Scholar] [CrossRef]
- Maltz, E.; Shkolnik, A. Milk composition and yield of black Bedouin goat during dehydration and rehydration. J. Dairy Res. 1984, 51, 23–27. [Google Scholar] [CrossRef]
- Silanikove, N.; Shamy, A.; Shinder, D.; Moran, A. Stress down regulates milk yield in cows by plasmin induced beta-casein product that blocks K+ channels on the apical membranes. Life Sci. 2000, 67, 2201–2212. [Google Scholar] [CrossRef]
- Hossaini-Hilali, J.; Benlamlih, S.; Dahlborn, K. Effects of dehydration, rehydration, and hyperhydration in the lactating and non-lactating black Moroccan goat. Comp. Biochem. Physiol. 1994, 109, 1017–1026. [Google Scholar] [CrossRef]
- El-Sherif, M.M.A.; Assad, F. Changes in some blood constituents in Barki ewes during pregnancy and lactation under semi-arid conditions. Small Rum. Res. 2001, 40, 269–277. [Google Scholar] [CrossRef]
- Abdelrahman, M.M.; Aljumaah, R.S. Metabolic profiles and milk composition of periparturient and early lactation periods in sheep. Asian J. Anim. Vet. Adv. 2012, 7, 1130–1138. [Google Scholar] [CrossRef]
- Dupoué, A.; Angelier, F.; Lourdais, O.; Bonnet, X.; Brischoux, F. Effect of water deprivation on baseline and stress-induced corticosterone levels in the Children’s python (Antaresiachildreni). Comp. Biochem. Physiol. A Mol. Integr. Physiol. 2014, 168, 11–16. [Google Scholar] [CrossRef]
- Maresh, C.M.; Gabaree-Boulant, C.L.; Armstrong, L.E.; Judelson, D.A.; Hoffman, J.R.; Castellani, J.W.; Kenefick, R.W.; Bergeron, M.F.; Casa, D.J. Effect of hydration status on thirst, drinking, and related hormonal responses during low-intensity exercise in the heat. J. Appl. Physiol. 2004, 97, 39–44. [Google Scholar] [CrossRef]
- Busch, D.S.; Hayward, L.S. Stress in a conservation context: A discussion of glucocorticoid actions and how levels change withconservation-relevantt variables. Biol. Conserv. 2009, 142, 2844–2853. [Google Scholar] [CrossRef]
- Sapolsky, R.M.; Romero, L.M.; Munck, A.U. How do glucocorticoids influence stress responses? Integrating permissive, suppressive, stimulatory, and preparative actions. Endocr. Rev. 2000, 21, 55–89. [Google Scholar]
- Davis, A.K.; Maney, D.L.; Maerz, J.C. The use of leukocyte profiles to measure stress in vertebrates: A review for ecologists. Rev. Funct. Ecol. 2008, 22, 760–772. [Google Scholar] [CrossRef]
- Ramirez, M.S.; Tolmasky, M.E. Aminoglycoside modifying enzymes. Drug Resist. Updat. 2010, 13, 151–171. [Google Scholar] [CrossRef]
- Sastrasinh, M.; Knauss, T.C.; Weinberg, J.M.; Humes, H.D. Identification of the aminoglycoside binding site in rat renal brus hborde rmembranes. J. Pharmacol. Exp. Ther. 1982, 222, 350–358. [Google Scholar]
- Elsheikh, H.A.; Intisar, O.M. Pharmacokinetics of ampicillin trihydrate following intravenous administratio no water-deprivedd Nubian goats. Res. Vet. Sci. 1998, 65, 111–114. [Google Scholar] [CrossRef]
- Elsheikh, H.A.; Intisar, O.M.; Abdullah, A. Pharmacokinetics of gentamicin administered intravenously to water-deprived goats. Small Rum. Res. 1999, 34, 15–20. [Google Scholar] [CrossRef]
- Oukessou, M.; Toutain, P.L. Effect of water deprivation on absorption (oral, intramuscular) and disposition of ampicillin in sheep. J. Vet. Pharmacol. Ther. 1992, 15, 421–432. [Google Scholar] [CrossRef]
- Hooper, L.; Bunn, D.K.; Abdelhamid, A.; Gillings, R.; Jennings, A.; Maas, K.; Millar, S.; Twomlow, E.; Hunter, P.R.; Shepstone, L.; et al. Water-loss (intracellular) dehydration assessed using urinary tests: How well do they work? Diagnostic accuracy in older people. Am. J. Clin. Nutr. 2016, 104, 121–131. [Google Scholar] [CrossRef]
- Singh, R.R.; Sajeesh, V.; Booth, L.C.; McArdle, Z.; May, C.N.; Head, G.A.; Moritz, K.M.; Schlaich, M.P.; Denton, K.M. Cather-Based Renal Denervation Exacerbates Blood Pressure Fall During Hemorrhage. J. Am. Coll. Cardiol. 2017, 69, 951–964. [Google Scholar] [CrossRef]
- Ali, M.A.; Adem, A.; Chandranath, I.S.; Benedict, S.; Pathan, J.Y.; Nagelkerke, N.; Nyberg, F.; Lewis, L.K.; Yandle, T.G.; Nicholls, G.M.; et al. Response to dehydration in the one-humped camel and effects of blocking therenin-angiotensinn system. PLoS ONE 2012, 7, e37299. [Google Scholar] [CrossRef]
- Mwacharo, J.M.; Kim, E.; Elbeltagy, A.; Aboul Naga, M.; Rischkowsky, B.; Rothschild, M.F. Genome-Widee Scans Reveal Multiple Selection Sweep Regions in Indigenous Sheep (Ovis Aries) from a Hot Arid Tropical Environment. In Proceedings of the Plant and Animal Genome Conference XXIV, San Diego, CA, USA, 9–13 January 2016; International Centre for Agricultural Research in the Dry Areas: Addis Ababa, Ethiopia, 2016. [Google Scholar]
- Elbeltagy, A.R.; Kim, E.; Rischkowsky, B.; Aboul-naga, A.M.; Mwacharo, J.M.; Rothschild, M.F. Genome-wide Analysis of Small Ruminant Tolerance to Grazing Stress Under Arid Desert. Anim. Ind. Rep. 2016, 662, R3106. [Google Scholar]
- Yang, J.; Li, W.; Lv, F.; He, S.; Tian, S. Whole-Genome Sequencing of Native Sheep Provides Insights into Rapid Adaptations to Extreme Environments. Mol. Biol. Evol. 2016, 33, 2576–2592. [Google Scholar] [CrossRef]


{kind=link}
| Ruminant Type/Distribution | Population Size (×103) | Breed Number | Population Share (%) | Breed Share (%) | Breeds per Million |
|---|---|---|---|---|---|
| Buffalo | |||||
| Africa | 0 | 3 | 0 | 3 | 0 |
| Asia + Pacific | 152,404 | 61 | 96 | 71 | 0.4 |
| Europe | 412 | 3 | 0 | 3 | 7.3 |
| LatinAmerica + Carribean | 1711 | 9 | 1 | 10 | 5.3 |
| Near East | 3998 | 10 | 1 | 12 | 2.5 |
| NorthAmerica | 0 | 0 | 0 | 0 | 0 |
| Total | 158,525 | 86 | |||
| Cattle | |||||
| Africa | 174,556 | 251 | 13 | 21 | 1.4 |
| Asia + Pacific | 461,197 | 236 | 35 | 19 | 0.5 |
| Europe | 162,119 | 482 | 12 | 39 | 3.0 |
| LatinAmerica + Carribean | 356,069 | 107 | 27 | 9 | 0.3 |
| Near East | 71,913 | 86 | 5 | 7 | 1.2 |
| NorthAmerica | 111,481 | 62 | 8 | 5 | 0.6 |
| Total | 1,337,335 | 1224 | |||
| Goat | |||||
| Africa | 137,104 | 89 | 19 | 16 | 0.6 |
| Asia + Pacific | 390,433 | 146 | 55 | 26 | 0.4 |
| Europe | 26,092 | 187 | 4 | 33 | 7.2 |
| LatinAmerica + Carribean | 40,752 | 34 | 6 | 6 | 0.8 |
| Near East | 114,572 | 94 | 16 | 16 | 0.8 |
| NorthAmerica | 1428 | 20 | 0 | 4 | 14.0 |
| Total | 710,381 | 570 | |||
| Sheep | |||||
| Africa | 127,440 | 147 | 12 | 11 | 1.2 |
| Asia + Pacific | 408,098 | 233 | 39 | 18 | 0.6 |
| Europe | 185,035 | 629 | 18 | 48 | 3.4 |
| LatinAmerica + Carribean | 89,372 | 42 | 8 | 3 | 0.5 |
| Near East | 242,770 | 201 | 23 | 15 | 0.8 |
| NorthAmerica | 7891 | 62 | 1 | 5 | 7.9 |
| Total | 1,060,606 | 1314 |
| Water Restriction | Specie | Average Tr (°C) | Age (Years) | Average Ambient Temperature (°C) | RH (%) | Ref. | |
|---|---|---|---|---|---|---|---|
| Water Restriction | Control | ||||||
| 3 days | Nubian goat | 37.90 | 39.90 s | 2–3 | 34.8 | 25.5 | [8] |
| 4 days | Sheep | 38.86 | 38.99 ns | Mature | 24.8 | NG | [60] |
| 4 days | Sheep | 39.78 | 39.37 ns | Mature | 0.4 | NG | [60] |
| 20%, 40% less ad lib and alternate day | Malpura ewes | 38.68, 38.80 and 38.74 respectively | 38.80 ns | 2–4 | 39.86 | 26.03 | [64] |
| 20%, 40% less ad lib and alternate day ad lib | Malpura ewes | 38.55, 38.46, and 38.47 respectively | 38.53 ns | 2–4 | 32.37 | 36.67 | [64] |
| Once every 3 days | Lactating Awassi ewes | 39.53 | 39.51 ns | mature | 27-31 | 61–85 | [48] |
| Once every 3 days | Dry Awassi ewes | 39.47 | 39.67 ns | mature | 27-31 | 61–85 | [48] |
| 3–15 h per day | German black-head mutton sheep | 38.7 | 39.0 ns | 1.8 | 12.7 | 73.5 | [65] |
| 3 h per day | German black-head mutton sheep | 38.6 | 38.7 ns | 1.8 | 12.7 | 73.5 | [65] |
| 6 h on the second day | German black-head mutton sheep | 38.3 | 38.8 ns | 1.8 | 12.7 | 73.5 | [65] |
| 3–15 h per day | Boer goats | 37.8 | 38.1 ns | 4.3 | 8.9 | 71.0 | [65] |
| 3 h per day | Boer goats | 37.8 | 38.1 ns | 4.3 | 8.9 | 71.0 | [65] |
| 6 h on the second day | Boer goats | 37.3 | 37.7 ns | 4.3 | 8.9 | 71.0 | [65] |
| 50% of ad lib | Baladi goat | 38.98 | 38.93 ns | 1.5-2 | NG | NG | [66] |
| Water Restriction | Specie | Average RR (Breath/Min) | Age (Years) | Ambient Temp. | RH (%) | Ref. | |
|---|---|---|---|---|---|---|---|
| Water Restriction | Control | ||||||
| 3–15 h per day | German black-head mutton sheep | 35.9 | 36.7 s | 1.8 | 12.7 | 73.5 | [65] |
| 3 h per day | German black-head mutton sheep | 34.1 | 36.1 s | 1.8 | 12.7 | 73.5 | [65] |
| 6 h on the second day | German black-head mutton sheep | 32.0 | 25.1 s | 1.8 | 12.7 | 73.5 | [65] |
| 3–15 h per day | Boer Goats | 20.5 | 20.1 s | 4.3 | 8.9 | 71.0 | [65] |
| 3 h per day | Boer Goats | 19.1 | 18.4 s | 4.3 | 8.9 | 71.0 | [65] |
| 6 h on the second day | Boer Goats | 18.7 | 17.6 s | 4.3 | 8.9 | 71.0 | [65] |
| 50% of ad lib | Baladi goat | 34.77 | 36.97 s | 1.5-2 | NG | NG | [66] |
| 80% and 60% ad lib | Lacauna ewes | 22.60 and 20.20 respectively | 26.20 s | mature | NG | NG | [49] |
| 20% and 40% ad lib and alternate day | Malpura ewes | 38.23, 40.40 and 37.03 respectively | 46.23 s | 2-4 | 32.37 | 36.67 | [64] |
| 20%, 40% ad lib and alternate day ad lib | Malpura ewes | 59.43, 62.87 and 60.69 | 67.26 ns | 2-4 | 39.86 | 26.03 | [64] |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Akinmoladun, O.F.; Muchenje, V.; Fon, F.N.; Mpendulo, C.T. Small Ruminants: Farmers’ Hope in a World Threatened by Water Scarcity. Animals 2019, 9, 456. https://doi.org/10.3390/ani9070456
Akinmoladun OF, Muchenje V, Fon FN, Mpendulo CT. Small Ruminants: Farmers’ Hope in a World Threatened by Water Scarcity. Animals. 2019; 9(7):456. https://doi.org/10.3390/ani9070456
Chicago/Turabian StyleAkinmoladun, Oluwakamisi F., Voster Muchenje, Fabian N. Fon, and Conference T. Mpendulo. 2019. "Small Ruminants: Farmers’ Hope in a World Threatened by Water Scarcity" Animals 9, no. 7: 456. https://doi.org/10.3390/ani9070456
APA StyleAkinmoladun, O. F., Muchenje, V., Fon, F. N., & Mpendulo, C. T. (2019). Small Ruminants: Farmers’ Hope in a World Threatened by Water Scarcity. Animals, 9(7), 456. https://doi.org/10.3390/ani9070456