

In Vitro and In Vivo Evaluation of Bacillus Strains as Prophylactic Agents Against Porcine Epidemic Diarrhea Virus

 , , and
, , and
Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Virus, Cell, and Bacterial Strains
2.2. Cytotoxicity Assay of Bacillus Strains
2.3. Assessment of the Prophylactic Effects of Bacillus Strains Against PEDV
2.4. Quantitative Reverse-Transcription (qRT-PCR) Analysis of PEDV-N, Type I IFN-Dependent Genes, and Proinflammatory Cytokines
2.5. Animal Feed Preparation and Piglet PEDV Infection
2.6. qRT-PCR Detection of Fecal Viral RNA
2.7. Metagenomic Analysis of Fecal Microbiota
2.8. Statistical Analysis
3. Results
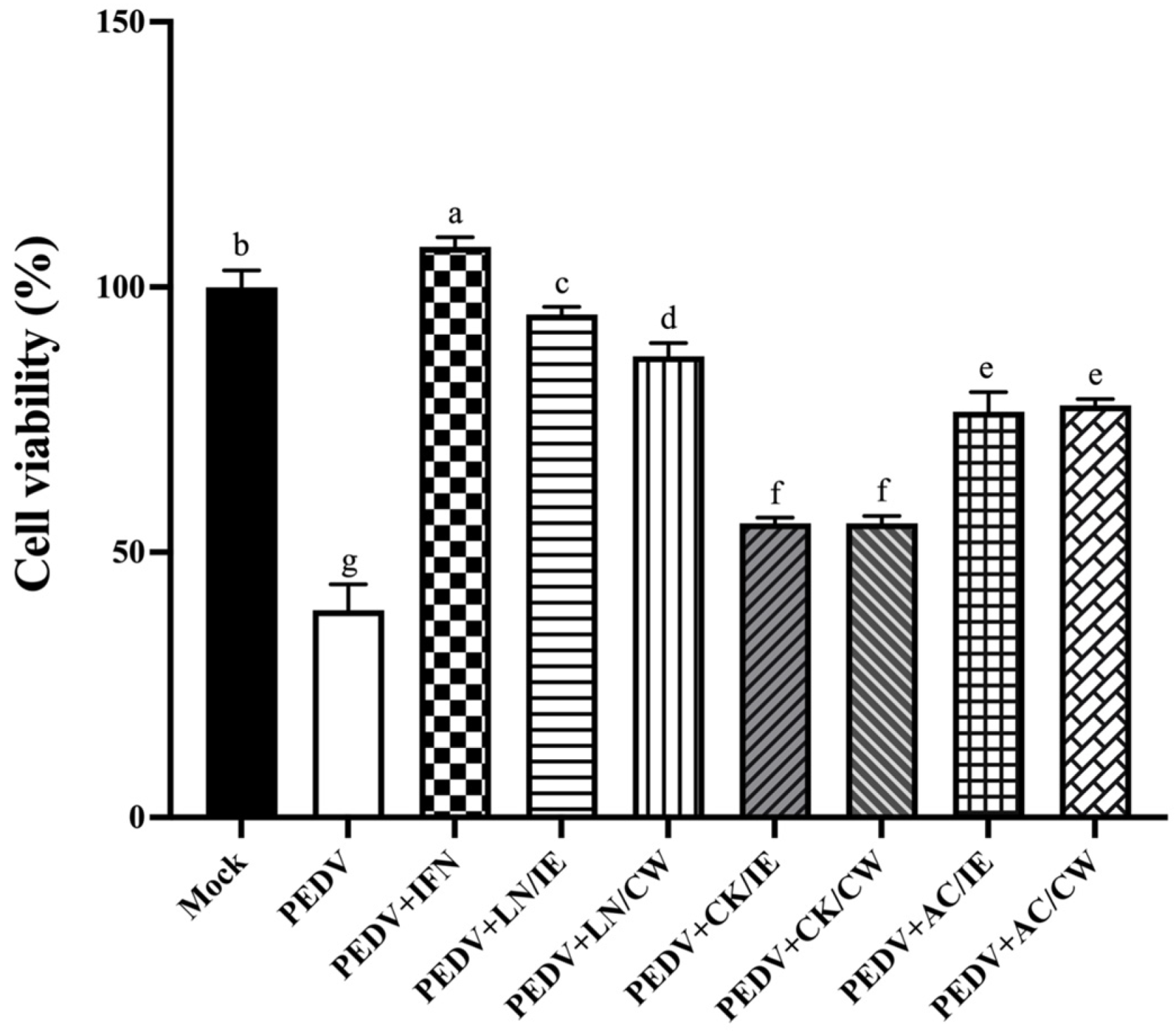
3.1. Cytotoxicity of Bacillus Strains on Vero Cells
3.2. Prophylactic Effects of Bacillus Strains Against PEDV
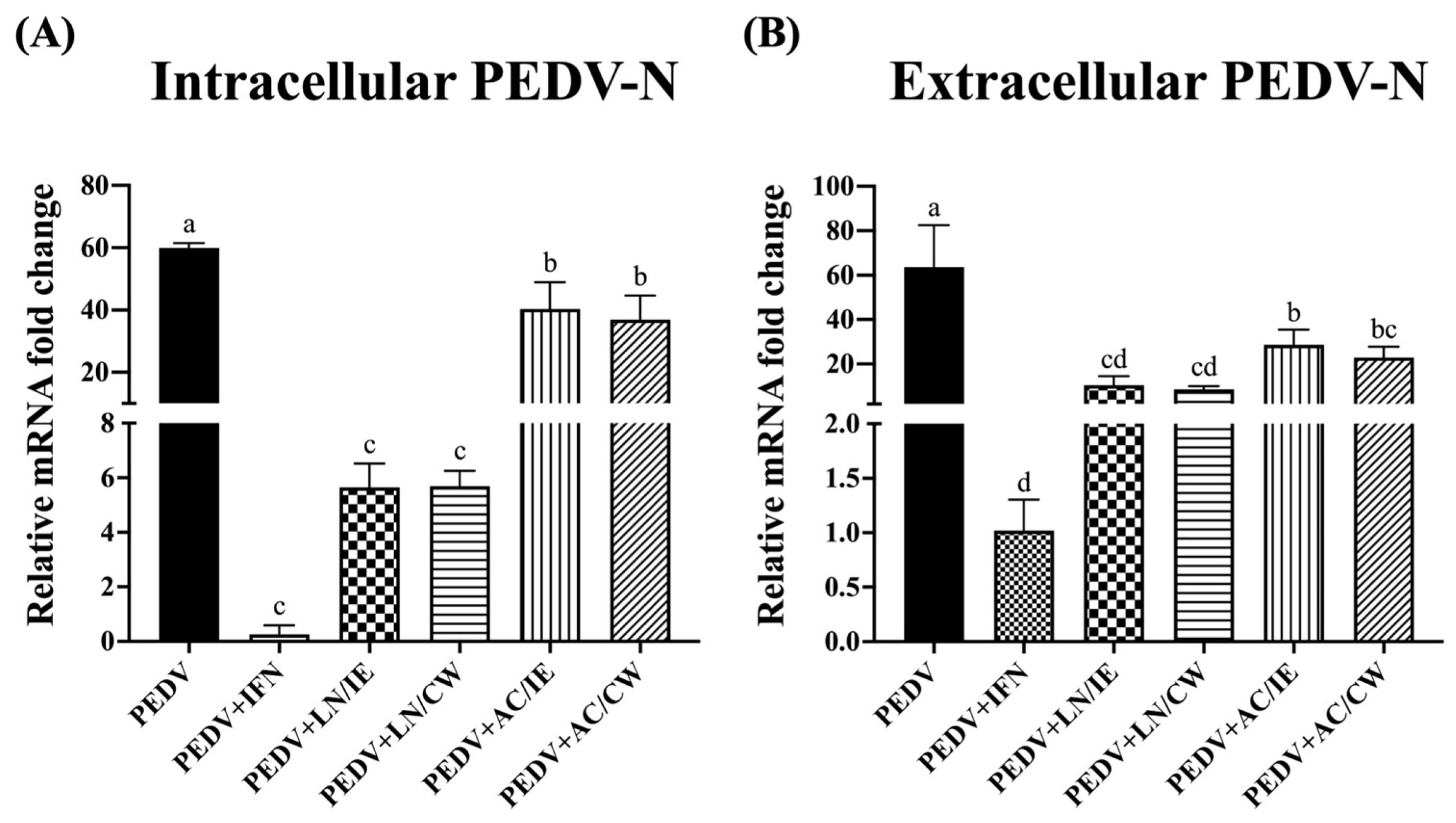
3.3. Effects on PEDV-N Protein Expression
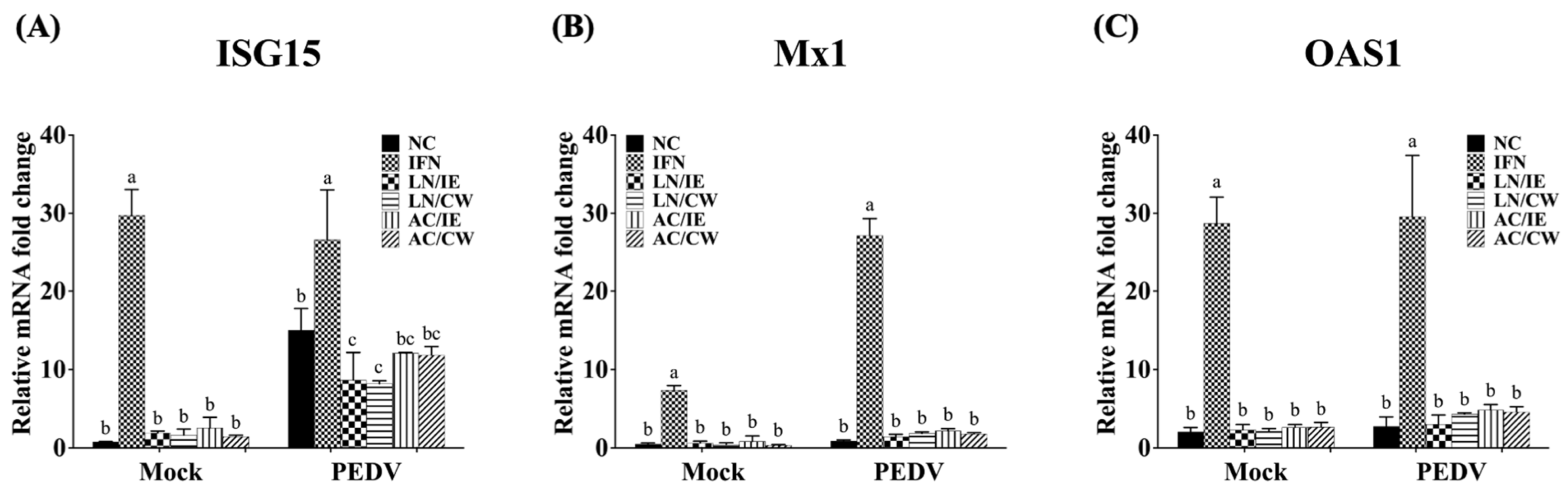
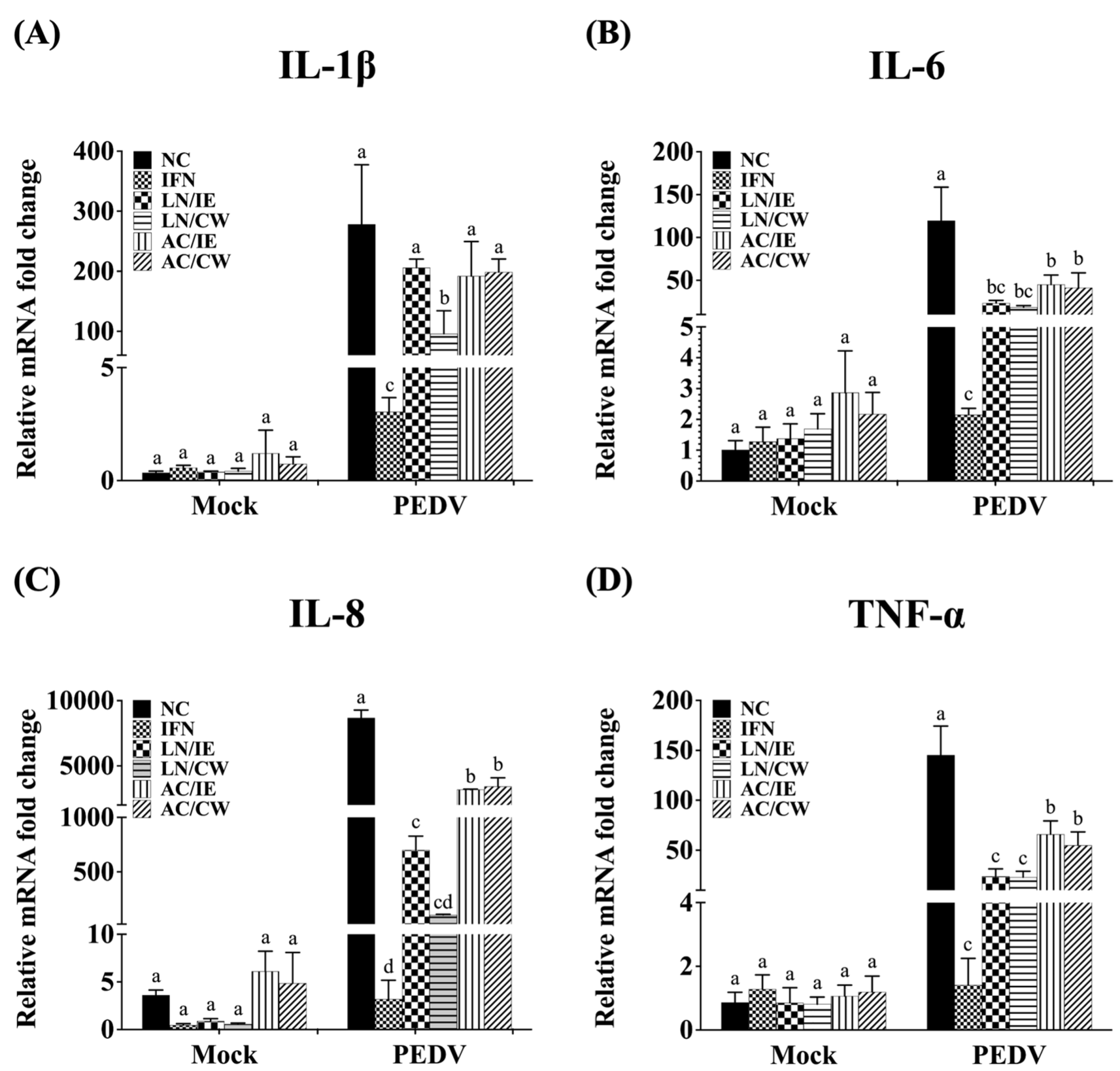
3.4. Effects on Type 1 IFN-Dependent Genes and Proinflammatory Cytokines
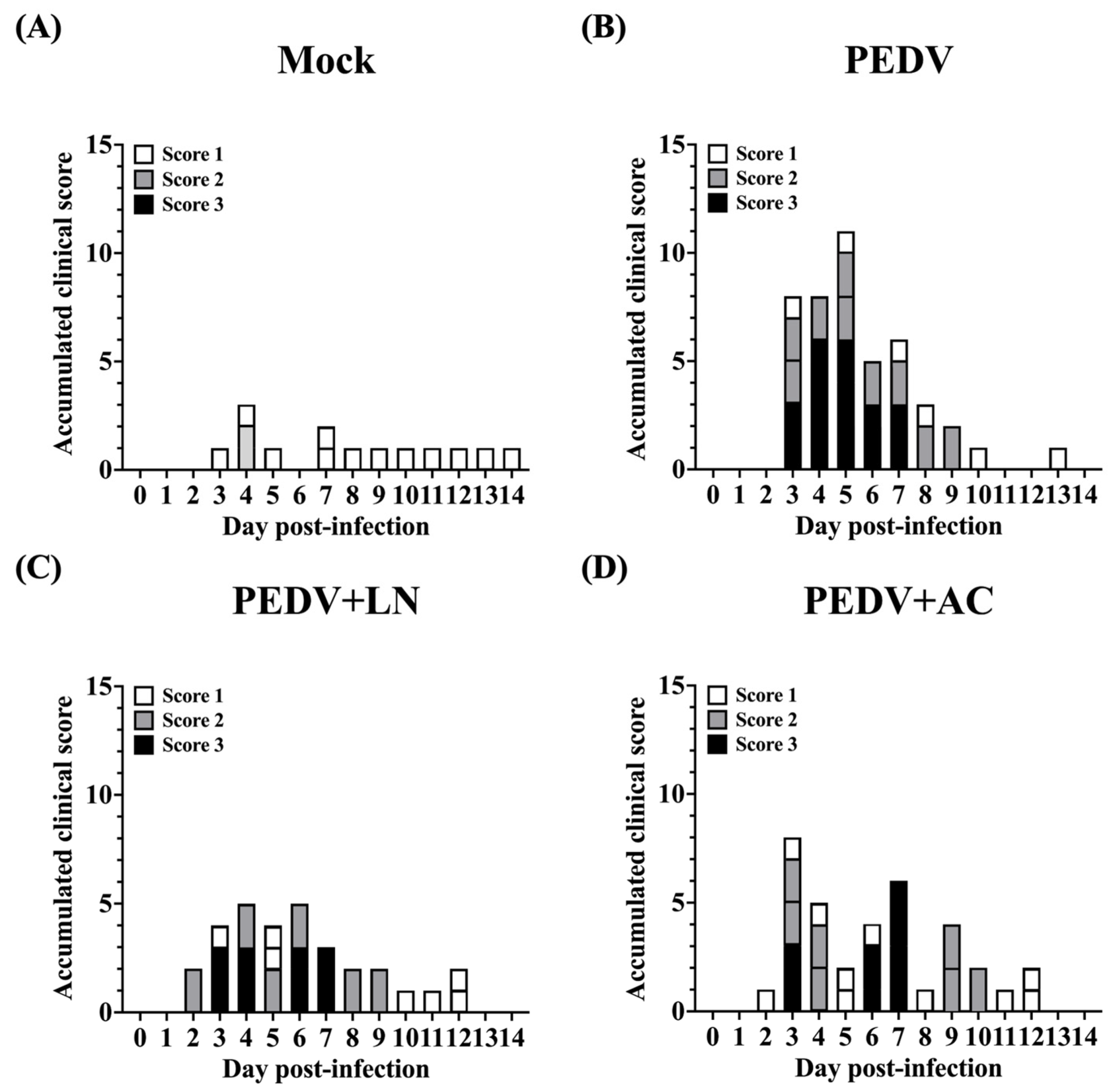
3.5. Effects of Bacillus Supplementation on Body Weight and Clinical Scoring of Fecal Consistency of Piglets
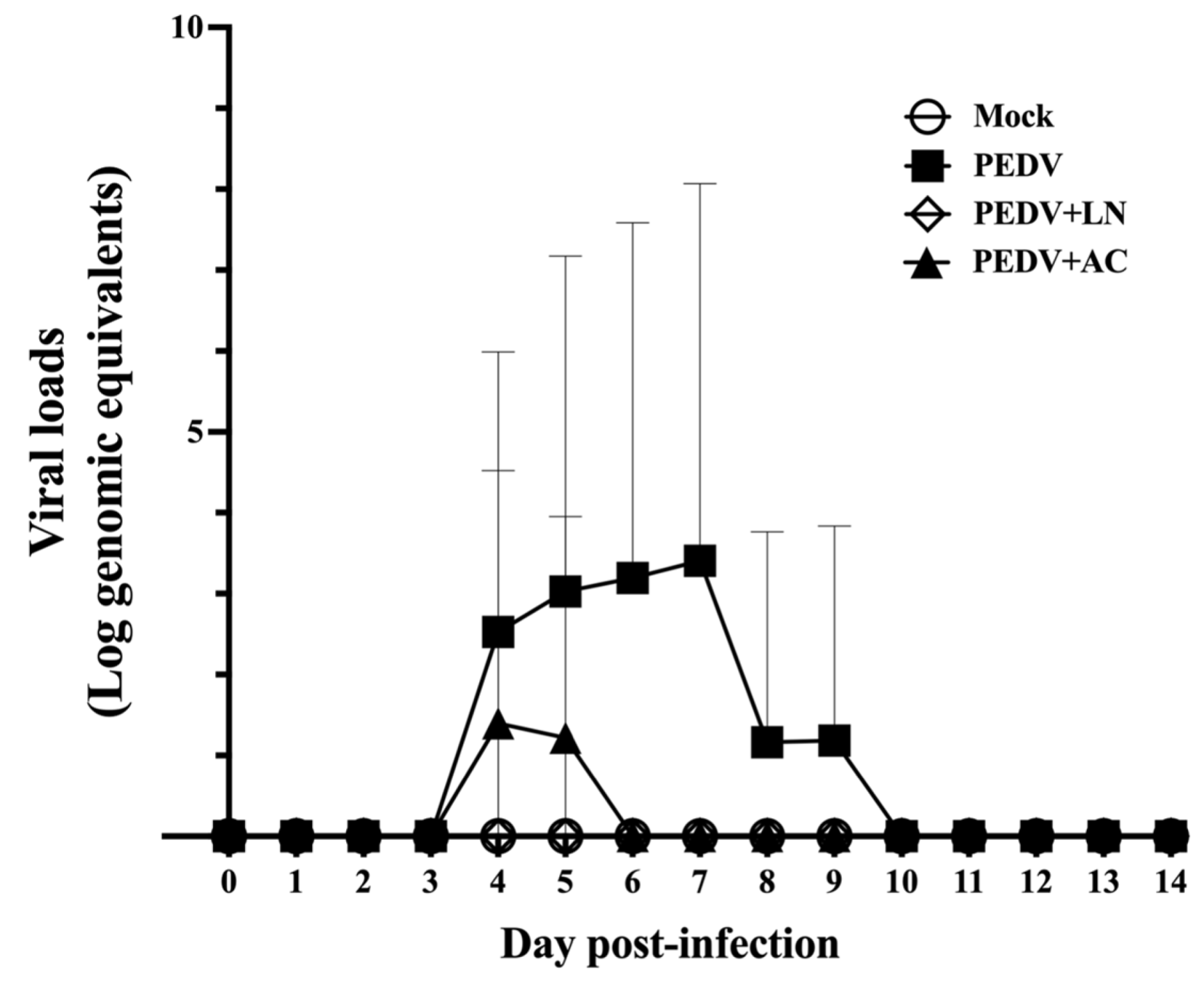
3.6. Fecal Viral RNA Shedding
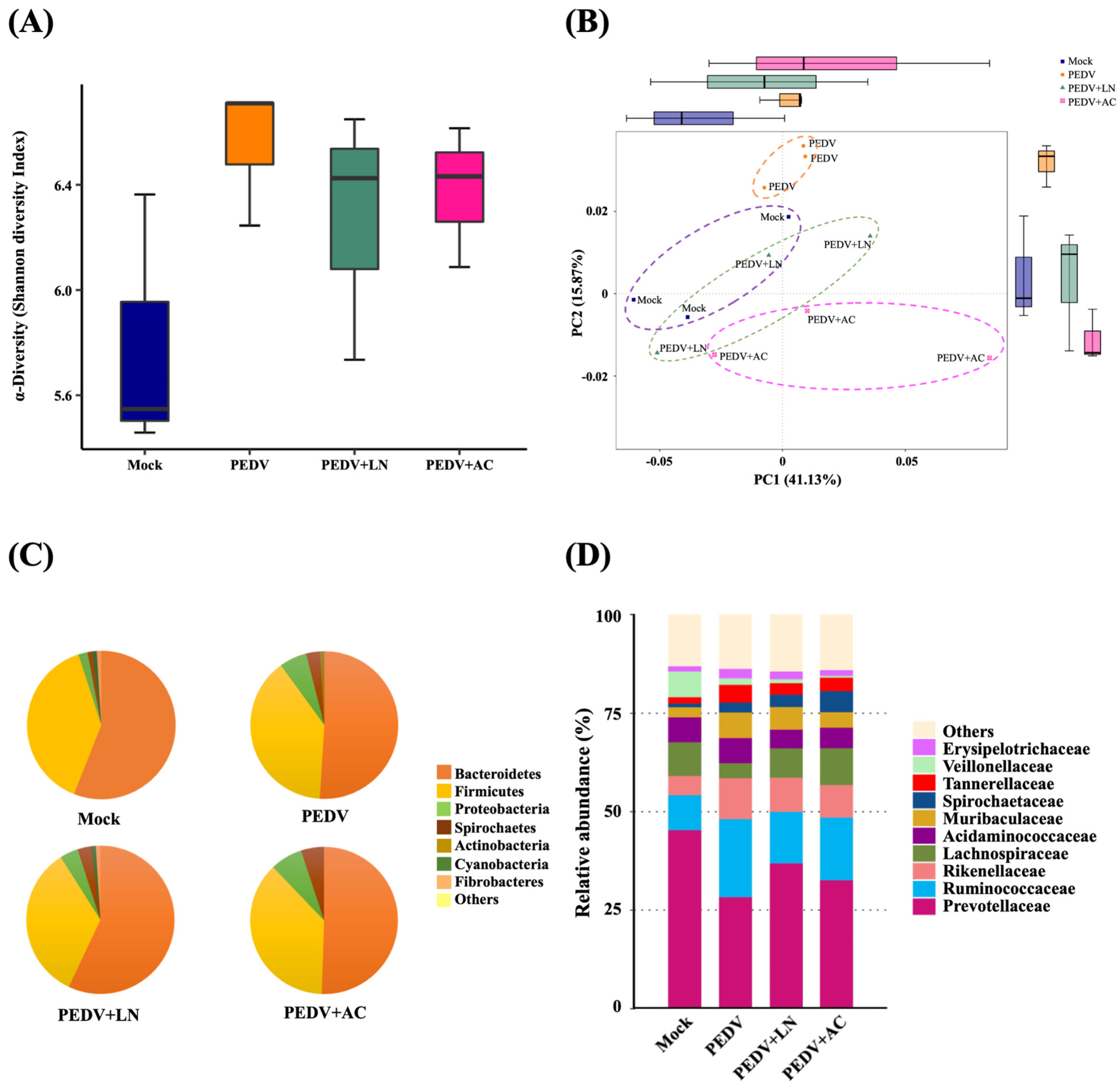
3.7. Fecal Microbiota Analysis
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Pensaert, M.B.; de Bouck, P. A New Coronavirus-like Particle Associated with Diarrhea in Swine. Arch. Virol. 1978, 58, 243–247. [Google Scholar] [CrossRef]
- Jung, K.; Saif, L.J.; Wang, Q. Porcine Epidemic Diarrhea Virus (PEDV): An Update on Etiology, Transmission, Pathogenesis, and Prevention and Control. Virus Res. 2020, 286, 198045. [Google Scholar] [CrossRef]
- Wood, E. An Apparently New Syndrome of Porcine Epidemic Diarrhoea. Vet. Rec. 1977, 100, 243. [Google Scholar] [CrossRef]
- Jang, G.; Lee, D.; Shin, S.; Lim, J.; Won, H.; Eo, Y.; Kim, C.-H.; Lee, C. Porcine Epidemic Diarrhea Virus: An Update Overview of Virus Epidemiology, Vaccines, and Control Strategies in South Korea. J. Vet. Sci. 2023, 24, e58. [Google Scholar] [CrossRef] [PubMed]
- Sun, R.-Q.; Cai, R.-J.; Chen, Y.-Q.; Liang, P.-S.; Chen, D.-K.; Song, C.-X. Outbreak of Porcine Epidemic Diarrhea in Suckling Piglets, China. Emerg. Infect. Dis. 2012, 18, 161–163. [Google Scholar] [CrossRef] [PubMed]
- Stevenson, G.W.; Hoang, H.; Schwartz, K.J.; Burrough, E.R.; Sun, D.; Madson, D.; Cooper, V.L.; Pillatzki, A.; Gauger, P.; Schmitt, B.J.; et al. Emergence of Porcine Epidemic Diarrhea Virus in the United States: Clinical Signs, Lesions, and Viral Genomic Sequences. J. Vet. Diagn. Investig. 2013, 25, 649–654. [Google Scholar] [CrossRef]
- Park, J.-E. Porcine Epidemic Diarrhea: Insights and Progress on Vaccines. Vaccines 2024, 12, 212. [Google Scholar] [CrossRef]
- Li, M.; Pan, Y.; Xi, Y.; Wang, M.; Zeng, Q. Insights and Progress on Epidemic Characteristics, Genotyping, and Preventive Measures of PEDV in China: A Review. Microb. Pathog. 2023, 181, 106185. [Google Scholar] [CrossRef]
- Jang, G.; Park, J.; Lee, C. Successful Eradication of Porcine Epidemic Diarrhea in an Enzootically Infected Farm: A Two-Year Follow-Up Study. Pathogens 2021, 10, 830. [Google Scholar] [CrossRef]
- Luise, D.; Bosi, P.; Raff, L.; Amatucci, L.; Virdis, S.; Trevisi, P. Bacillus Spp. Probiotic Strains as a Potential Tool for Limiting the Use of Antibiotics, and Improving the Growth and Health of Pigs and Chickens. Front. Microbiol. 2022, 13, 801827. [Google Scholar] [CrossRef]
- Peng, J.-Y.; Horng, Y.-B.; Wu, C.-H.; Chang, C.-Y.; Chang, Y.-C.; Tsai, P.-S.; Jeng, C.-R.; Cheng, Y.-H.; Chang, H.-W. Evaluation of Antiviral Activity of Bacillus licheniformis-Fermented Products against Porcine Epidemic Diarrhea Virus. AMB Express 2019, 9, 191. [Google Scholar] [CrossRef] [PubMed]
- Yuan, L.; Zhang, S.; Wang, Y.; Li, Y.; Wang, X.; Yang, Q. Surfactin Inhibits Membrane Fusion during Invasion of Epithelial Cells by Enveloped Viruses. J. Virol. 2018, 92, e00809-18. [Google Scholar] [CrossRef]
- Chen, Y.-M.; Limaye, A.; Chang, H.-W.; Liu, J.-R. Screening of Lactic Acid Bacterial Strains with Antiviral Activity Against Porcine Epidemic Diarrhea. Probiotics Antimicrob. Proteins 2022, 14, 546–559. [Google Scholar] [CrossRef]
- Chang-Liao, W.-P.; Lee, A.; Chiu, Y.-H.; Chang, H.-W.; Liu, J.-R. Isolation of a Leuconostoc mesenteroides Strain With Anti-Porcine Epidemic Diarrhea Virus Activities From Kefir Grains. Front. Microbiol. 2020, 11, 1578. [Google Scholar] [CrossRef]
- Sirichokchatchawan, W.; Temeeyasen, G.; Nilubol, D.; Prapasarakul, N. Protective Effects of Cell-Free Supernatant and Live Lactic Acid Bacteria Isolated from Thai Pigs Against a Pandemic Strain of Porcine Epidemic Diarrhea Virus. Probiotics Antimicro. Proteins 2018, 10, 383–390. [Google Scholar] [CrossRef]
- Payne, J.; Bellmer, D.; Jadeja, R.; Muriana, P. The Potential of Bacillus Species as Probiotics in the Food Industry: A Review. Foods 2024, 13, 2444. [Google Scholar] [CrossRef] [PubMed]
- Larsen, N.; Thorsen, L.; Kpikpi, E.N.; Stuer-Lauridsen, B.; Cantor, M.D.; Nielsen, B.; Brockmann, E.; Derkx, P.M.F.; Jespersen, L. Characterization of Bacillus Spp. Strains for Use as Probiotic Additives in Pig Feed. Appl. Microbiol. Biot. 2014, 98, 1105–1118. [Google Scholar] [CrossRef]
- Chang, Y.-C.; Kao, C.-F.; Chang, C.-Y.; Jeng, C.-R.; Tsai, P.-S.; Pang, V.F.; Chiou, H.-Y.; Peng, J.-Y.; Cheng, I.-C.; Chang, H.-W. Evaluation and Comparison of the Pathogenicity and Host Immune Responses Induced by a G2b Taiwan Porcine Epidemic Diarrhea Virus (Strain Pintung 52) and Its Highly Cell-Culture Passaged Strain in Conventional 5-Week-Old Pigs. Viruses 2017, 9, 121. [Google Scholar] [CrossRef]
- Hsu, T.-C.; Yi, P.-J.; Lee, T.-Y.; Liu, J.-R. Probiotic Characteristics and Zearalenone-Removal Ability of a Bacillus licheniformis Strain. PLoS ONE 2018, 13, e0194866. [Google Scholar] [CrossRef]
- Lee, A.; Cheng, K.-C.; Liu, J.-R. Isolation and Characterization of a Bacillus amyloliquefaciens Strain with Zearalenone Removal Ability and Its Probiotic Potential. PLoS ONE 2017, 12, e0182220. [Google Scholar] [CrossRef] [PubMed]
- Yi, P.-J.; Pai, C.-K.; Liu, J.-R. Isolation and Characterization of a Bacillus licheniformis Strain Capable of Degrading Zearalenone. World J. Microbiol. Biotechnol. 2011, 27, 1035–1043. [Google Scholar] [CrossRef]
- Mosmann, T. Rapid Colorimetric Assay for Cellular Growth and Survival: Application to Proliferation and Cytotoxicity Assays. J. Immunol. Methods 1983, 65, 55–63. [Google Scholar] [CrossRef]
- Shen, H.; Zhang, C.; Guo, P.; Liu, Z.; Sun, M.; Sun, J.; Li, L.; Dong, J.; Zhang, J. Short Communication: Antiviral Activity of Porcine IFN-λ3 against Porcine Epidemic Diarrhea Virus in Vitro. Virus Genes 2016, 52, 877–882. [Google Scholar] [CrossRef] [PubMed]
- Guo, X.; Zhang, M.; Zhang, X.; Tan, X.; Guo, H.; Zeng, W.; Yan, G.; Memon, A.M.; Li, Z.; Zhu, Y.; et al. Porcine Epidemic Diarrhea Virus Induces Autophagy to Benefit Its Replication. Viruses 2017, 9, 53. [Google Scholar] [CrossRef]
- Sun, P.; Wu, H.; Huang, J.; Xu, Y.; Yang, F.; Zhang, Q.; Xu, X. Porcine Epidemic Diarrhea Virus through P53-Dependent Pathway Causes Cell Cycle Arrest in the G0/G1 Phase. Virus Res. 2018, 253, 1–11. [Google Scholar] [CrossRef]
- Herlemann, D.P.R.; Labrenz, M.; Jürgens, K.; Bertilsson, S.; Waniek, J.J.; Andersson, A.F. Transitions in Bacterial Communities along the 2000 Km Salinity Gradient of the Baltic Sea. ISME J. 2011, 5, 1571–1579. [Google Scholar] [CrossRef] [PubMed]
- Jung, K.; Wang, Q.; Scheuer, K.A.; Lu, Z.; Zhang, Y.; Saif, L.J. Pathology of US Porcine Epidemic Diarrhea Virus Strain PC21A in Gnotobiotic Pigs—Volume 20, Number 4—April 2014—Emerging Infectious Diseases Journal—CDC. Emerg. Infect. Dis. 2014, 20, 668–671. [Google Scholar] [CrossRef]
- Caporaso, J.G.; Kuczynski, J.; Stombaugh, J.; Bittinger, K.; Bushman, F.D.; Costello, E.K.; Fierer, N.; Peña, A.G.; Goodrich, J.K.; Gordon, J.I.; et al. QIIME Allows Analysis of High-Throughput Community Sequencing Data. Nat. Methods 2010, 7, 335–336. [Google Scholar] [CrossRef] [PubMed]
- Edgar, R.C. UPARSE: Highly Accurate OTU Sequences from Microbial Amplicon Reads. Nat. Methods 2013, 10, 996–998. [Google Scholar] [CrossRef]
- Edgar, R.C. Search and Clustering Orders of Magnitude Faster than BLAST. Bioinformatics 2010, 26, 2460–2461. [Google Scholar] [CrossRef]
- Wang, Q.; Garrity, G.M.; Tiedje, J.M.; Cole, J.R. Naive Bayesian Classifier for Rapid Assignment of RRNA Sequences into the New Bacterial Taxonomy. Appl. Environ. Microbiol. 2007, 73, 5261–5267. [Google Scholar] [CrossRef]
- Quast, C.; Pruesse, E.; Yilmaz, P.; Gerken, J.; Schweer, T.; Yarza, P.; Peplies, J.; Glöckner, F.O. The SILVA Ribosomal RNA Gene Database Project: Improved Data Processing and Web-Based Tools. Nucleic Acids Res. 2012, 41, D590–D596. [Google Scholar] [CrossRef] [PubMed]
- Shannon, C.E. A Mathematical Theory of Communication. Bell Syst. Tech. J. 1948, 27, 379–423. [Google Scholar] [CrossRef]
- Jiang, X.-T.; Peng, X.; Deng, G.-H.; Sheng, H.-F.; Wang, Y.; Zhou, H.-W.; Tam, N.F.-Y. Illumina Sequencing of 16S RRNA Tag Revealed Spatial Variations of Bacterial Communities in a Mangrove Wetland. Microb. Ecol. 2013, 66, 96–104. [Google Scholar] [CrossRef]
- Koonpaew, S.; Teeravechyan, S.; Frantz, P.N.; Chailangkarn, T.; Jongkaewwattana, A. PEDV and PDCoV Pathogenesis: The Interplay Between Host Innate Immune Responses and Porcine Enteric Coronaviruses. Front. Vet. Sci. 2019, 6, 34. [Google Scholar] [CrossRef]
- Hao, Z.; Fu, F.; Cao, L.; Guo, L.; Liu, J.; Xue, M.; Feng, L. Tumor Suppressor P53 Inhibits Porcine Epidemic Diarrhea Virus Infection via Interferon-Mediated Antiviral Immunity. Mol. Immunol. 2019, 108, 68–74. [Google Scholar] [CrossRef]
- Simmons, J.D.; Wollish, A.C.; Heise, M.T. A Determinant of Sindbis Virus Neurovirulence Enables Efficient Disruption of Jak/STAT Signaling. J. Virol. 2010, 84, 11429–11439. [Google Scholar] [CrossRef] [PubMed]
- Huan, C.; Wang, H.; Sheng, X.; Wang, R.; Wang, X.; Liao, Y.; Liu, Q.; Tong, G.; Ding, C.; Fan, H.; et al. Porcine Epidemic Diarrhea Virus Nucleoprotein Contributes to HMGB1 Transcription and Release by Interacting with C/EBP-β. Oncotarget 2016, 7, 75064–75080. [Google Scholar] [CrossRef]
- Yu, H.; Chen, G.; Zhang, T.; Huang, X.; Lu, Y.; Li, M.; Li, S.; Wang, C.; Li, B.; Zhang, Y.; et al. PEDV Promotes the Differentiation of CD4+T Cells towards Th1, Tfh, and Treg Cells via CD103+DCs. Virology 2023, 587, 109880. [Google Scholar] [CrossRef] [PubMed]
- Du, J.; Luo, J.; Yu, J.; Mao, X.; Luo, Y.; Zheng, P.; He, J.; Yu, B.; Chen, D. Manipulation of Intestinal Antiviral Innate Immunity and Immune Evasion Strategies of Porcine Epidemic Diarrhea Virus. Biomed. Res. Int. 2019, 2019, 1862531. [Google Scholar] [CrossRef]
- Yang, S.; Liu, G.; Savelkoul, H.F.J.; Jansen, C.A.; Li, B. Mini-Review: Microbiota Have Potential to Prevent PEDV Infection by Improved Intestinal Barrier. Front. Immunol. 2023, 14, 1230937. [Google Scholar] [CrossRef]
- Shin, N.-R.; Whon, T.W.; Bae, J.-W. Proteobacteria: Microbial Signature of Dysbiosis in Gut Microbiota. Trends Biotechnol. 2015, 33, 496–503. [Google Scholar] [CrossRef]
- Xu, Z.; Zhang, Q.; Wu, M.; Zhang, Y.; Li, Z.; Li, H.; Yu, C.; Zhang, X.; Zhao, D.; Wang, L.; et al. Lactobacillus rhamnosus GG Powder Supplementation Alleviates Intestinal Injury in Piglets Challenged by Porcine Epidemic Diarrhea Virus. Front. Cell. Infect. Microbiol. 2024, 14, 1371916. [Google Scholar] [CrossRef]
- Xing, J.-H.; Niu, T.-M.; Zou, B.-S.; Yang, G.-L.; Shi, C.-W.; Yan, Q.-S.; Sun, M.-J.; Yu, T.; Zhang, S.-M.; Feng, X.-Z.; et al. Gut Microbiota-Derived LCA Mediates the Protective Effect of PEDV Infection in Piglets. Microbiome 2024, 12, 20. [Google Scholar] [CrossRef]

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene | Primer Sequence (5′ to 3′) | Reference |
|---|---|---|
| ISG15 | Forward: GGGCAACGAGTTCCAGGT Reverse: CACCACCAGCAGGACCGT | [23] |
| MX1 | Forward: GCAGCCAGTACGAGGAGAAG Reverse: CTCCTGACAGTGCCTCCAAC | [23] |
| OAS1 | Forward: GGTTGTCTTCCTCAGTCCTC Reverse: AGCCTGGACCTCAAACTTCA | [23] |
| IL-1β | Forward: GCGGCAACGAGGATGACTT Reverse: TGGCTACAACAACTGACACGG | [24] |
| IL-6 | Forward: TGTGAAAGCAGCAAAGAG Reverse: AGTGTCCTCATTGAATCCA | [24] |
| IL-8 | Forward: GGAACCATCTCGCTCTGTGTAA Reverse: GGTCCACTCTCAATCACTCTCAG | [24] |
| TNF-α | Forward: GTGCTGGTGACTTTGGTGCTA Reverse: GAGAAGCCTCAGGTCCCAAT | [24] |
| PEDV-N | Forward: TGAGGGTGTTTTCTGGGTTG Reverse: TTGCCATTGCCACGACTC | [25] |
| GAPDH | Forward: AGCCAAAAGGGTCATCATCT Reverse: ATGAGTCCTTCCACGATACC | [23] |
| 16S rRNA | Forward: CCTACGGGNGGCWGCAG Reverse: GACTACHVGGGTATCTAATCC | [26] |
| Group | 0 DPI | 7 DPI | 14 DPI |
|---|---|---|---|
| Mock 1 | 6.00 ± 2.36 | 9.10 ± 1.91 | 12.57 ± 3.06 |
| PEDV | 6.51 ± 1.30 | 10.17 ± 2.75 | 12.43 ± 2.02 |
| PEDV+LN | 7.00 ± 2.17 | 9.73 ± 2.63 | 11.93 ± 2.89 |
| PEDV+AC | 6.64 ± 1.23 | 11.00 ± 1.32 | 12.77 ± 1.89 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Chen, Y.-J.; Tsai, C.-F.; Hsu, C.-W.; Chang, H.-W.; Liu, J.-R. In Vitro and In Vivo Evaluation of Bacillus Strains as Prophylactic Agents Against Porcine Epidemic Diarrhea Virus. Animals 2025, 15, 470. https://doi.org/10.3390/ani15040470
Chen Y-J, Tsai C-F, Hsu C-W, Chang H-W, Liu J-R. In Vitro and In Vivo Evaluation of Bacillus Strains as Prophylactic Agents Against Porcine Epidemic Diarrhea Virus. Animals. 2025; 15(4):470. https://doi.org/10.3390/ani15040470
Chicago/Turabian StyleChen, You-Jia, Chia-Fang Tsai, Chin-Wei Hsu, Hui-Wen Chang, and Je-Ruei Liu. 2025. "In Vitro and In Vivo Evaluation of Bacillus Strains as Prophylactic Agents Against Porcine Epidemic Diarrhea Virus" Animals 15, no. 4: 470. https://doi.org/10.3390/ani15040470
APA StyleChen, Y.-J., Tsai, C.-F., Hsu, C.-W., Chang, H.-W., & Liu, J.-R. (2025). In Vitro and In Vivo Evaluation of Bacillus Strains as Prophylactic Agents Against Porcine Epidemic Diarrhea Virus. Animals, 15(4), 470. https://doi.org/10.3390/ani15040470