1. Introduction
Coccidiosis is a potentially debilitating disease caused by the
Eimeria parasite, characterized by impaired growth performance and decreased feed conversion rates [
1]. It is estimated that the global economic burden of coccidiosis exceeded £ 12.9 billion in 2022 [
2]. Among the various
Eimeria species,
Eimeria necatrix is identified as a highly pathogenic protozoan, primarily targeting the intestinal tract of chickens and inducing acute intestinal coccidiosis in poultry [
3]. Currently, the primary strategies for controlling chicken coccidiosis rely on chemoprophylaxis and the administration of live attenuated vaccines. However, limitations in vaccine efficacy, the emergence of drug-resistant
Eimeria strains, and increasing regulatory restrictions on antibiotic usage have prompted a shift in control strategies [
4]. In this context, subunit vaccines derived from parasite-specific antigens or recombinant proteins expressed from cloned DNA represent a promising alternative to overcome these challenges [
5].
Immunoprotective genes from the
Eimeria genome have been identified and selected for cloning and expression, facilitating the development of highly immunogenic subunit vaccine candidates [
6]. To date, CoxAbic
®, the only commercially available subunit vaccine against chicken coccidiosis, comprises three major gametocyte-derived antigens from
Eimeria maxima: 230 kDa, 82 kDa, and 56 kDa proteins. Field trials have demonstrated that CoxAbic
® reduces oocyst shedding by approximately 50–80% [
7]. Inspired by these findings, researchers have focused on identifying additional gametocyte antigens, such as GAM22 [
8,
9,
10], GAM56 [
10,
11,
12,
13], and GAM59 [
14], which may further enhance vaccine efficacy.
Gam22 is the first identified multi-copy gene within
Eimeria species, characterized by an exceptionally high copy number and highly conserved sequences among its copies [
8]. EtGAM22 is localized to wall-forming bodies type 2 (WFB2), where it is involved in the formation of the inner oocyst wall and/or the Stieda body in
Eimeria tenella [
10]. In contrast, EnGAM22 is distributed within wall-forming bodies type 1 (WFB1) and plays a role in the formation of the outer oocyst wall in
E. necatrix [
15]. The
Gam56 gene is a single-copy, intronless gene initially identified in the gametocytes of
E. maxima [
11] and encodes the EmGAM56, which localizes to both WFB2 and the inner oocyst wall in
E. maxima. The
Gam59 gene (also referred to as Et
gam56 tmp2), encoding a GAM56-like protein (GAM59), was first identified in the
E. tenella genome and is positioned adjacent to the
gam56 cDNA (Et
gam56 tmp1) [
10]. In
E. necatrix, EnGAM59 similarly localizes to WFB2 and contributes to the formation of the inner oocyst wall [
15]. Previous studies in our group demonstrated that En
gam22 and En
gam59 are promising candidates for the development of recombinant subunit vaccines against coccidiosis [
9,
14]. Further investigation of the
gam56 gene is expected to yield valuable insights into the design of more effective vaccination strategies for the prevention and control of coccidiosis.
In this study, we successfully cloned and expressed the full-length Engam56 gene (Engam56-F) and subsequently evaluated its potential, along with that of a truncated version (Engam56-T), as candidate subunit immunogens for protection against E. necatrix infection. In addition, the protective efficacy of immunization with individual recombinant proteins (rEnGAM22, rEnGAM56, and rEnGAM59) and their combination was assessed in chickens.
2. Materials and Methods
2.1. Protein, Animals, and Parasites
Recombinant gametocyte proteins of
E. necatrix-rEnGAM22, rEnGAM59, and the truncated rEnGAM56-T-were successfully expressed in our laboratory using a prokaryotic expression system [
9,
14]. rEnGAM22 contains a region rich in histidine and proline [
9], while rEnGAM59 protein contains both a tyrosine-serine-rich region and a proline-methionine-rich region [
14]. rEnGAM56-T, a truncated form of the rEnGAM56-F protein, retains a domain enriched in tyrosine and serine [
9]. Preliminary evaluations have shown that both rEnGAM22 and rEnGAM59 are immunogenic and confer protective immunity against
E. necatrix infection.
One-day-old yellow-feathered broiler chickens were purchased from Jiangsu Jinghai Poultry Industry Group Co., Ltd. (Nantong, Jiangsu, China), reared in a coccidian-free environment, and provided with water and food ad libitum.
The
E. necatrix Yangzhou strain was previously isolated and is routinely maintained in our laboratory. A total of 20 chickens, aged 15 days, were orally inoculated with 20,000 sporulated oocysts. Feces samples were collected daily from 7 to 12 days post-infection (PI). Unsporulated oocysts were isolated and purified using a saturated salt solution floatation method, as described in previous reports to allow for sporulation [
16]. The resulting sporulated oocysts were then stored at 4 °C in 2.5% (
w/
v) potassium dichromate until further use. Gametocytes were isolated and purified from experimentally infected chickens using previously published methods [
15]. Briefly, mucosal tissues were scraped from the caecum of chickens infected with 30,000 sporulated oocysts, and the tissues were digested in SAC buffer (1 mM phenylmethanesulfonyl fluoride (PMSF), 1 mg/mL bovine serum albumin, 170 mM NaCl, 10 mM Tris-HCl (pH 7.0), 10 mM glucose, and 5 mM CaCl
2) containing 0.5 mg/mL hyaluronidase. The digested material was sequentially filtered through 17 μm and 10 μm polymer meshes, with gametocytes retained on the mesh being washed with cold SAC, collected by centrifugation at 1000×
g for 5 min, and stored at −80 °C until further use.
2.2. Plasmid Construction
The total RNA of gametocytes was extracted and transcribed into cDNA using an RNA Extraction Kit (TaKaRa, Tokyo, Japan) and Reverse Transcriptase Kit (TaKaRa), following the manufacturer’s instructions. As the amplicons were expected to have high GC contents, the full-length
gam56 gene (En
gam56-F) was amplified in segments using RT-PCR with PrimeSTAR
® GXL DNA Polymerase (TaKaRa). Three overlapping fragments were amplified with three pairs of primers designed based on the
gam56 sequence of the
E. necatrix Houghton strain (GenBank accession number: XM_013578447.1) (
Table 1). The PCR cycling conditions were as follows: 98 °C for 10 s, 58 °C for 30 s, and 68 °C for 1.5 min.
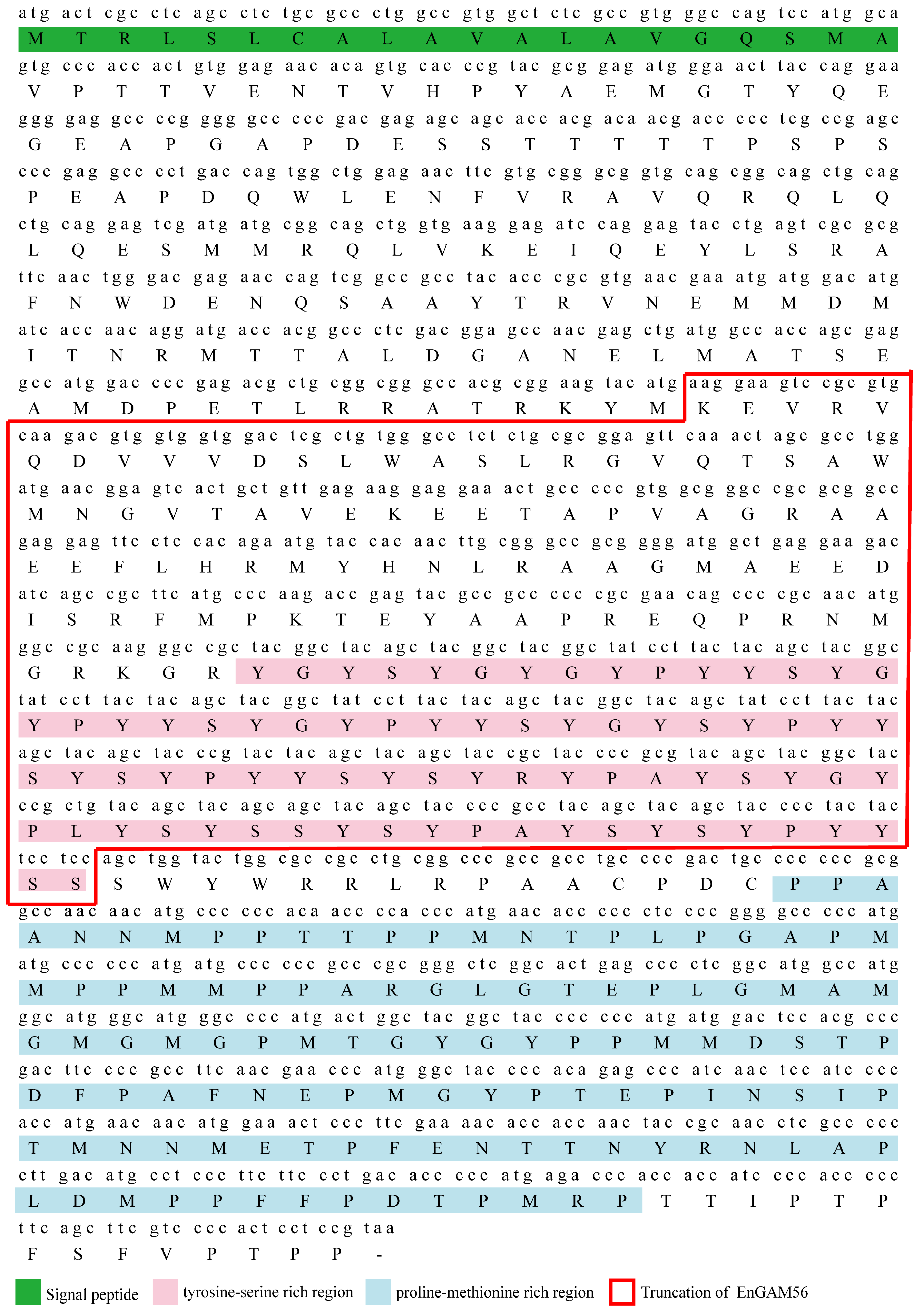
The fragments were subsequently subcloned into pGEM®-T-easy vector (Promega, Madison, WI, USA) and sequenced by Beijing Genomics Institute (BGI). The obtained sequences were spliced and analyzed using the DNAMAN 6.0 software. After removing the signal peptides (first 20 amino acids), the Engam56-F gene was further optimized for prokaryotic expression and synthesized by Genscript (Nanjing, China).
2.3. Expression and Purification of Recombinant Proteins
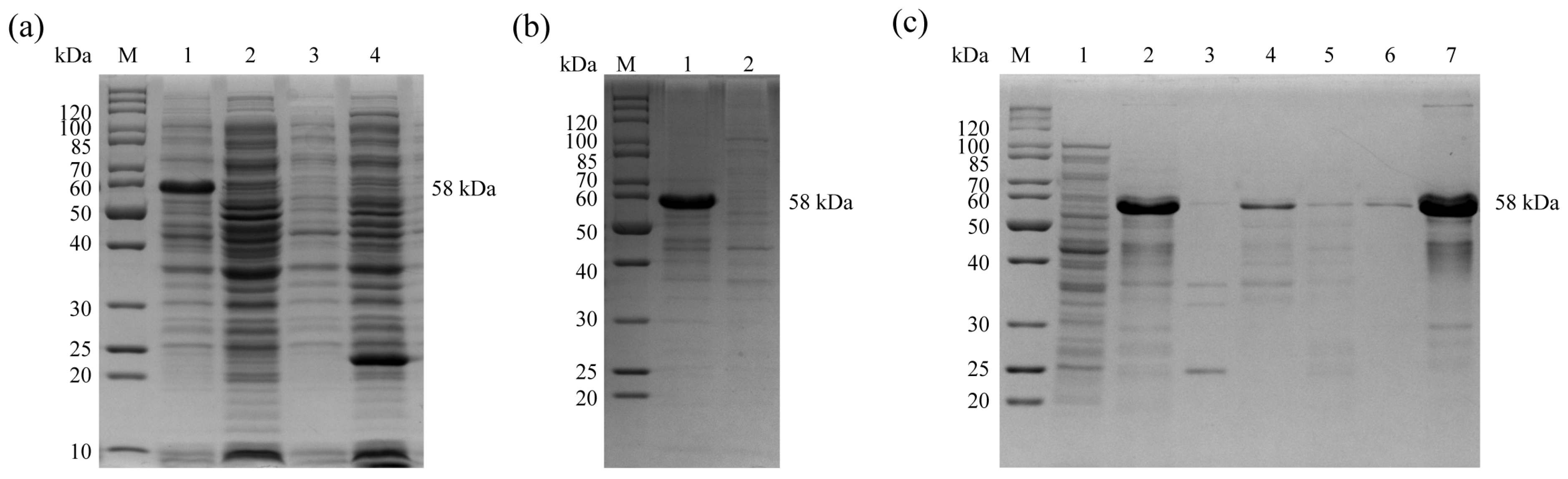
Recombinant expressing bacteria pET28a(+)-En
gam56-F/BL21 was cultured in LB medium at 37 °C to OD600 = 0.8, and a final concentration of 1.0 mM isopropyl β-D-1-thiogalactopyranoside (IPTG; Promega Corp., Madison, WI, USA) was added to induce recombinant protein (rEnGAM56-F) production. The rEnGAM56-F was purified using a Nickel-Nitrilotriacetic Acid (Ni-NTA) affinity chromatography column (GenScript, Nanjing, China) according to our previous study [
9], analyzed by 12% SDS-PAGE with Coomassie Brilliant Blue G-250 staining (Sigma-Aldrich, St Louis, MO, USA), and treated with the ToxinEraser endotoxin removal kit (Genscript).
2.4. Preparation of Immunoglobulin Serum
Mouse anti-rEnGAM56-F polyclonal antibody (anti-rEnGAM56-F pAb) was generated by immunizing six 6-week-old female BALB/c mice with rEnGAM56-F. Each mouse was intramuscularly injected with rEnGAM56-F (20 μg of protein in 50 μL of PBS), emulsified with 50 μL of QuickAntibody-Mouse3W adjuvant (Biodragon, Beijing, China). Booster immunizations were administered 14 days later, and blood was collected to isolate serum 21 days later.
Convalescent serum of chicken with E. necatrix infection had already been prepared and stored at −80 °C in our lab.
2.5. Western Blot Analysis
Total protein was extracted from gametocytes placed in 1.5 mL RNase-free microcentrifuge tubes containing 200 μL cell lysis buffer (Xinsaimei, Shanghai, China) and 2 μL protease inhibitor (Xinsaimei). The samples were incubated on ice for 30 min, followed by ultrasonic disruption in an ice bath for 5 min (30% power, 2 s on/3 s off cycles). The lysates were then centrifuged at 10,000× g for 10 min at 4 °C, and the resulting supernatants containing total protein were collected for further analysis.
The identification of EnGAM56-F protein and rEnGAM56-F was performed using the Wes Capillary Western Blot System (ProteinSimple, San Jose, CA, USA), with a 12–230 kDa Separation Module (#SM-W003). The antigen specificity of rEnGAM56-F was also analyzed using this system. Briefly, the mouse anti-6 × HIS tag monoclonal antibody (1:100 dilution, BIO BASIC, Markham, Canada), mouse negative serum (1:50 dilution), mouse anti-rEnGAM56-F pAb (1:50 dilution), chicken negative serum (1:20 dilution), and the convalescent serum of chicken infected with E. necatrix (1:20 dilution) were used as primary antibodies, respectively. The HRP-labeled rabbit anti-mouse IgG (1:1000 dilution, BIO BASIC) and rabbit anti-chicken IgG (1:1000 dilution, BIO BASIC) were used as secondary antibodies. All antibodies were diluted in antibody diluent (ProteinSimple). Positive signals were visualized using Compass for SW software (V4.0.0, Protein Simple).
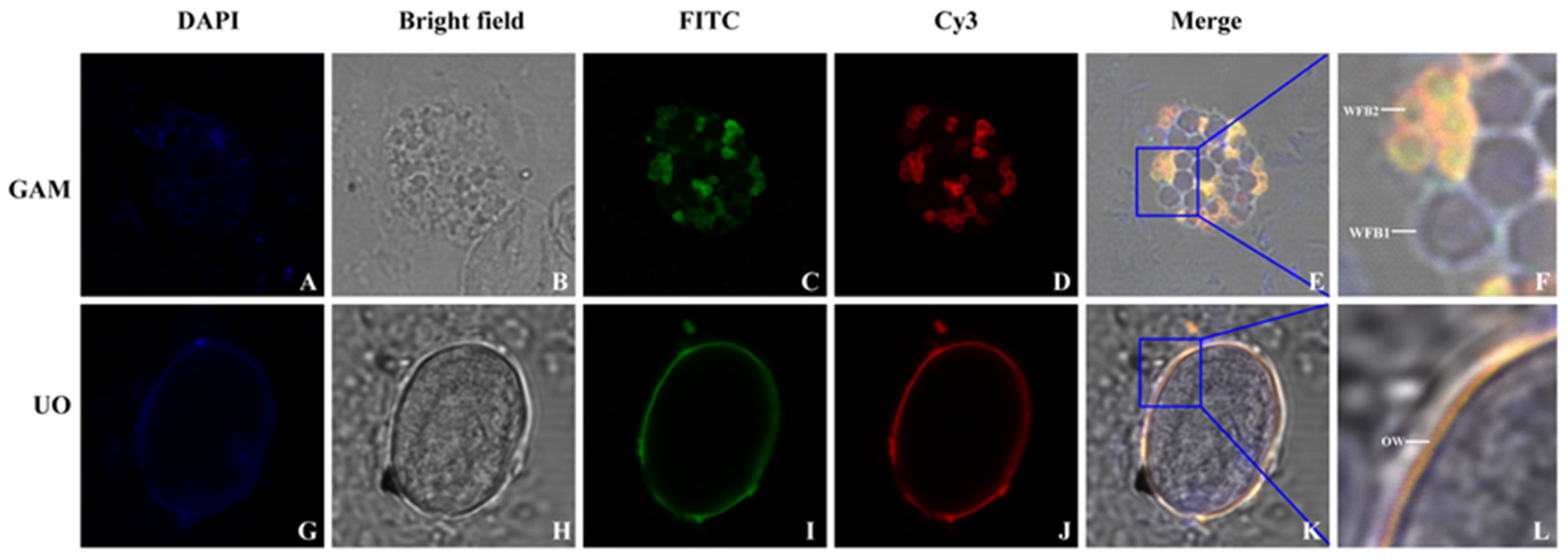
2.6. Indirect Immunofluorescence Localization
The rabbit anti-rEnGAM59 pAb specifically localized EnGAM59 to the WFB2 in gametocytes, serving as a marker to differentiate between WFB1 and WFB2 [
15].
Freshly isolated gametocytes and unsporulated oocysts were evenly spread onto glass slides and fixed with methanol at −20 °C for 10 min. The samples were then permeabilized with 0.1% Triton X-100 (Beyotime, Shanghai, China) for 10 min at room temperature. After being blocked with 3% BSA in PBS for 3 h at 37 °C, the slides were incubated overnight at 4 °C with anti-rEnGAM56-F pAb (1:200 dilution) or rabbit anti-rEnGAM59 pAb (1:100 dilution, prepared in our laboratory). The slides were washed three times with 0.03% Tween-20 in PBS (PBST) for 15 min each and then incubated for 1 h at 37 °C with either fluorescein isothiocyanate (FITC)-conjugated goat anti-rabbit IgG (1:100 dilution; MultiSciences, Hangzhou, China) or Cy3-conjugated goat anti-mouse IgG (1:100 dilution; MultiSciences).
Under the fluorescence microscope (Leica DM2500, Leica Microsystems GmbH, Wetzlar, Germany), sealing tables with anti-fluorescence quenching agents (Roche, Basel, Switzerland) were examined.
2.7. Vaccinations and Challenge Experiment
Two immunization trials were conducted. The first trial aimed to evaluate the protective efficacy of rEnGAM56-F and rEnGAM56-T, as well as to determine the optimal immunization dosage. The second trial was designed to assess the protective efficacy of a combined immunization strategy involving rEnGAM56, rEnGAM22, and rEnGAM59.
For immune dose screening, doses of 50 μg, 100 μg, and 200 μg were selected based on previous findings from our laboratory to evaluate the immune protective efficacy of different gametocyte proteins [
14,
17]. Accordingly, chickens immunized with rEnGAM56-F and rEnGAM56-T received doses of 50 μg, 100 μg, and 200 μg, respectively. For rEnGAM22, prior studies demonstrated that a 50 μg dose provided optimal immune protection [
9], while a 200 μg dose was most effective for rEnGAM59 [
14]. In the present study, the immunization dose of rEnGAM59 was reduced to 100 μg per chicken to minimize protein load, ensuring safety and preventing potential overstimulation of the immune system.
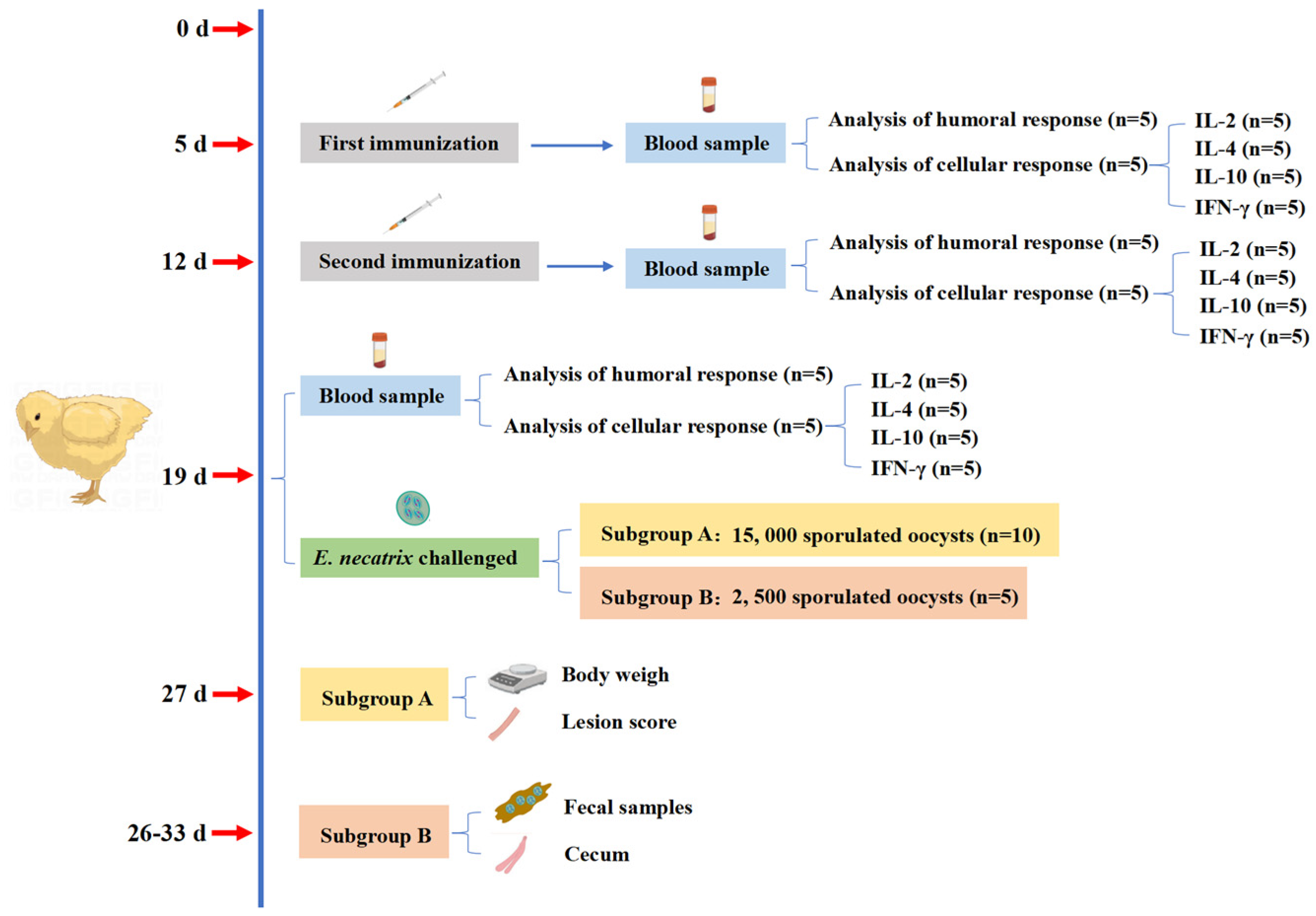
The experimental design and its immunization procedure were shown in
Figure 1. Chickens aged 5 days within a similar weight range were randomly divided into eight groups, each consisting of 15 chickens. Experimental groups contained immunized groups (IC): rEnGAM56-F immunized groups (rEnGAM56-F-H: 200 μg per chicken, rEnGAM56-F-M: 100 μg per chicken, and rEnGAM56-F-L: 50 μg per chicken) and rEnGAM56-T immunized groups (rEnGAM56-T-H: 200 μg per chicken, rEnGAM56-T-M: 100 μg per chicken, and rEnGAM56-T-L: 50 μg per chicken). Control groups included the unimmunized and challenged group (UC) and the unimmunized and unchallenged group (UU).
In a combined protocol, the rEnGAM22, rEnGAM56-T, and rEnGAM59 were combined to immunize chickens in different combinations. The immunized groups consisted of three single-immunized groups, namely rEnGAM22 (50 μg per chicken), rEnGAM56-T (50 μg per chicken), and rEnGAM59 (100 μg per chicken), as well as two co-immunization groups: One co-immunized with rEnGAM22 (50 μg per chicken) and rEnGAM56-T (50 μg per chicken), and the other co-immunized with rEnGAM22 (50 μg per chicken), rEnGAM56-T (50 μg per chicken), and rEnGAM59 (100 μg per chicken). Control groups included an unimmunized and challenged group (UC) and an unimmunized and unchallenged group (UU). Immunizations were administered via subcutaneous injection at one-week intervals. Complete Freund’s adjuvant (Sigma-Aldrich) was used for the first immunization, while incomplete Freund’s adjuvant (Sigma-Aldrich) was utilized for the second immunization. Recombinant protein was diluted in PBS and mixed with the corresponding adjuvant at a 1:1 volume ratio.
To evaluate vaccine efficacy, a low-dose vaccination strategy was employed to better simulate natural infection conditions and enable a more accurate assessment of immune protection. Seven days after the second immunization (day 19, D19), each group was randomly divided into two subgroups. Subgroup A (n = 10) was orally challenged with 15,000 sporulated oocysts, while subgroup B (n = 5) received 2,500 sporulated oocysts.
For subgroup A, the protective efficacy of the treatments was evaluated based on survival rate, body weight gain (BWG), relative weight gain (RWG), and lesion score. The survival rate (%) was calculated with the formula (number surviving per group/total number of each group) × 100. BWG1 was calculated for days 5–19 in the immunization period, BWG2 was calculated for days 19–27 in the challenge period, and RWG (%) was calculated with the formula BWG of IC or UC groups/BWG of UU group × 100. Lesion scores were assessed on day 8 PI (27 days of age) using a 0 to 4 scale, as described by Johnson and Reid [
18]. Two independent observers assessed the extent of intestinal bleeding, evaluated changes in the intestinal wall, and examined the condition of intestinal contents, assigning lesion scores accordingly. For subgroup B, the protective efficacy was evaluated based on oocyst reduction (OR). The OR (%) was calculated with the formula (OR of UC group—OR of IC groups)/OR of UC group) × 100. The oocyst shedding was measured between days 7 and 14 PI (26–33 days of age) using the McMaster counting method, as previously described [
19].
The anticoccidial index (ACI) was used to evaluate the anticoccidial efficacy. The ACI is calculated as ACI = (survival rate + RWG) × 100 − (lesion value + oocyst value). The lesion value is lesion score × 10. The oocyst value is assigned based on the oocyst ratio: 0 if the oocyst ratio is 0–1%, 5 if 1–25%, 10 if 26–50%, 20 if 51–75%, and 40 if 76–100%, where the oocyst ratio is defined as the oocyst outputs of IC groups divided by that of UC groups. The ACI values above 180 were deemed excellent, 160–180 as marked, 140–160 as moderate, 120–140 was classified as mild or slight, and below 120 as inefficacy [
19].
2.8. Detection of Serum Antibody Levels
Blood samples were collected from five randomly selected chickens in each group at three time points: Prior to the first immunization (Day 5, D5), 7 days after the primary immunization (Day 12, D12), and 7 days after the secondary immunization (Day 19, D19). The samples were kept at room temperature for 2 h, then centrifuged at 2500 g for 10 min. The resulting serum was stored at −80 °C for subsequent analysis.
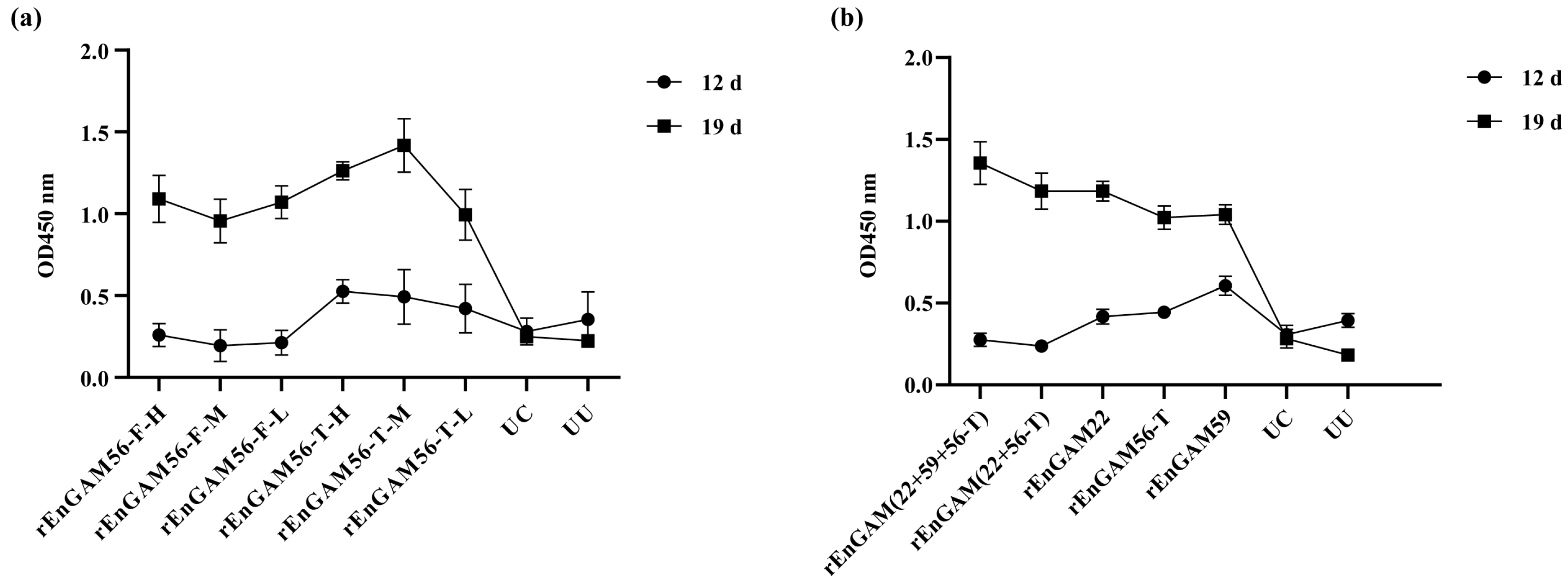
Indirect enzyme-linked immunosorbent assays (ELISAs) were employed to detect serum antibody levels of the chickens. Briefly, 96-well plates were coated overnight at 4 °C with 1 μg/well of purified rEnGAM22, rEnGAM59, rEnGAM56-F, or rEnGAM56-T. For the combined immunization group, rEtGAM22 and rEtGAM56 were mixed at a 1:1 ratio and coated, and rEtGAM22, rEtGAM56, and rEtGAM59 were mixed at a 1:1:1 ratio before coating. Plates were washed three times with 0.05% Tween-20 in PBS (PBST) for 15 min and then blocked with 1.0% bovine serum albumin (BSA; Sigma-Aldrich) in PBS for 1.5 h at 37 °C. After blocking, diluted serum samples (1:200) were added to the wells and incubated at 37 °C for 1.5 h. Plates were then washed three times with PBST, and bound antibodies were detected using horseradish peroxidase (HRP)-conjugated goat anti-chicken IgY (H + L) antibody (1:20,000; GenScript). The optical density at 450 nm (OD450) was measured using an ELISA reader (Sunrise-Basic, Tecan Trading AG, Männedorf, Switzerland). All samples were assayed in triplicate.
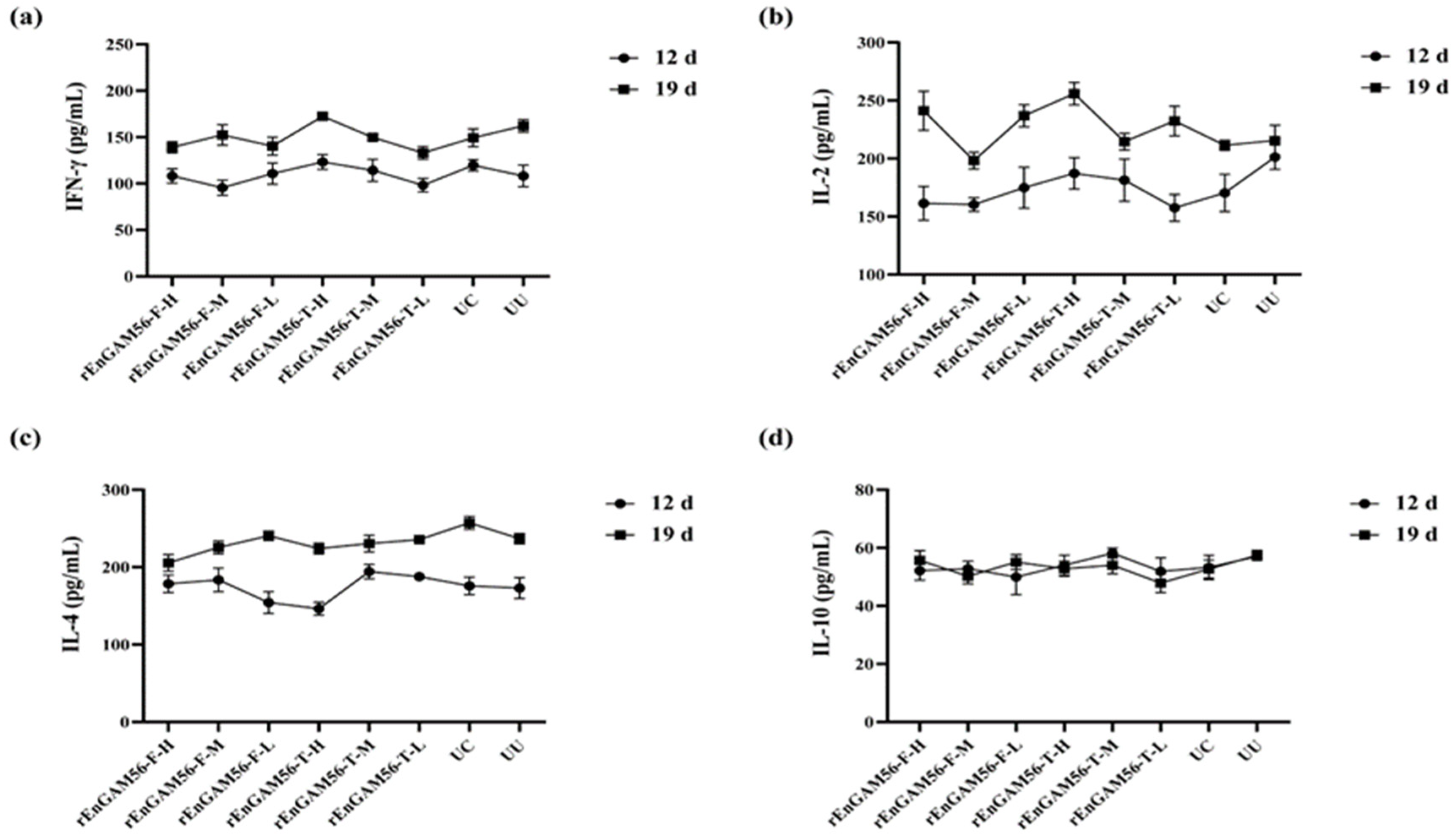
2.9. Determination of Serum Cytokine Levels
The cytokines interleukin-2 (IL-2), interleukin-4 (IL-4), interleukin-10 (IL-10), and interferon-γ (IFN-γ) were quantified in serum samples collected from five chickens per group on D5, D12, and D19. Quantification was performed using commercial cytokine detection kits (MEIMIAN Bio, Yancheng, China) following the manufacturer’s protocols. All serum samples were analyzed in triplicate.
2.10. Statistical Analysis
All data were expressed as means ± SEM, and statistical analysis was performed using the GraphPad Prism (GraphPad Prism 8.0, USA). ANOVA was used to test if there are statistically significant differences between treated and control groups. Significant differences were referred to as *: p < 0.05, **: p < 0.01, ***: p < 0.001, and ****: p < 0.0001.
4. Discussion
The gametocyte proteins serve as a precursor to the oocyst wall protein [
20,
21]. Several gametocyte antigens have been studied for immunization targets against various parasites, including
E. maxima [
22,
23],
E. tenella [
8,
13], and
Plasmodium falciparum [
24]. However, few reports have focused on gametocyte antigens from
E. necatrix. In our previous work, we successfully cloned and expressed the
E. necatrix gametocyte antigens EnGAM22 and EnGAM59. Subsequent studies demonstrated that the rEnGAM22 and rEnGAM59 provide protective immunity against
E. necatrix infection, with optimal immunizing doses determined to be 50 μg and 100 μg, respectively.
Belli et al. [
11,
12,
25] observed that EmGAM56 and EmGAM82 were proteolytically processed into small tyrosine-rich peptides with molecular weights of 8, 10, 12, and 31 kDa during oocyst wall formation. Comparable tyrosine-rich proteins originating from gametocyte proteins have also been identified in
E. tenella and
Eimeria acervuline [
26]. In
E. maxima, two gametocyte proteins, EmGAM82 (characterized by a high content of tyrosine and serine) and EmGAM56 (enriched in both tyrosine-serine and proline-methionine motifs), are localized within the WFBII and contribute to the formation of the inner oocyst wall. Similarly, EnGAM59, a structural homolog of EmGAM56 identified in
E. necatrix, contains tyrosine-serine- and proline-methionine-rich domains, enabling its association with WFBII and playing a critical role in the formation of the inner oocyst wall. In contrast, EnGAM22, which is rich in proline and histidine residues, localizes to WFBI and is associated with the outer oocyst wall in
E. necatrix. In the present study, the full-length EnGAM56, which includes both tyrosine-rich and proline-methionine-rich regions, exhibits an amino acid composition similar to that of EtGAM56 and EmGAM56 and was found to localize to both WFBII and the inner oocyst wall. These findings suggest distinct molecular pathways underlying the formation of the outer and inner oocyst walls.
Wallach et al. [
7] found that maternal antibodies produced by breeder hens immunized with
E. maxima gametocyte antigens confer partial protective immunity to offspring against both homologous and heterologous
Eimeria infections. Building upon these findings, our previous studies have further confirmed that gametocyte protein 56 (GAM56) represents a promising candidate antigen for the development of vaccines against coccidial infections [
27,
28,
29]. In this study, our results revealed that both rEnGAM56-F and rEnGAM56-T could provide effective immune protection against
E. necatrix infection, with the protective effects being stronger with rEnGAM56-T. In particular, the group rEnGAM56-T-L achieved the best performance in RWG (84.09%), oocyst production (66.20%), and ACI (158.59), followed by the group rEnGAM56-F-L. The oocyst reduction rate in the rEnGAM56-T-M group reached 58.2%, and the ACI index was 152.79.
The full-length rEnGAM56 protein exhibited lower immunoprotective efficacy compared to its truncated counterpart, aligning with findings reported in previous studies [
30,
31]. This reduced efficacy may be attributed to the masking of critical antigenic epitopes by other domains within the full-length protein, thereby diminishing the host’s immune recognition and response. Moreover, the structural complexity of the full-length protein increases the likelihood of misfolding, adversely affecting both antigen presentation and protein stability. In contrast, the truncated form facilitates better exposure of key antigenic epitopes, reduces the potential for misfolding, and enhances overall structural stability, collectively contributing to a more robust immune response. Additionally, truncation may eliminate immunosuppressive epitopes present in the full-length protein, further improving its immunogenicity. In terms of antibody levels, group rEnGAM56-T-L showed lower antibody levels following the second immunization compared to group rEnGAM56-T-M. This phenomenon may be attributed to the typical temporal lag in antibody production, wherein antibody titers do not reach their peak until a defined period following infection or immunization. During this interval, despite relatively low circulating antibody levels, other branches of the immune system, such as cellular immunity, may have been activated and effectively suppressed the
E. necatrix infection. This early immunity may account for the higher ACI observed in the rEnGAM56-T-L group.
Coccidiosis is a complex parasitic disease, and relying on a single antigen presents challenges in effectively targeting its diverse antigenic epitopes and multiple developmental stages. Therefore, incorporating a combination of antigens or multi-epitope vaccine strategies may be necessary to enhance protective efficacy and broaden the immune response spectrum. In an earlier study, immunization with 50 μg of rEnGAM22 resulted in a 15.82% reduction in oocyst shedding. Li [
32] reported a 39.55% reduction in oocyst output and an 85.31% relative weight gain in birds immunized with 200 μg of rEnGAM59. Based on these findings, the immunization dose for rEnGAM22 in the present study was set at 50 μg. Although 200 μg was previously identified as the optimal dose for rEtGAM59, this was reduced to 100 μg in the combined immunization strategy to minimize the total protein load per bird, thereby ensuring safety and avoiding potential immunity overstimulation. Remarkably, in the present study, the rEnGAM (22 + 59 + 56-T) and rEnGAM (22 + 56-T) groups achieved RWG values of 96.16% and 91.56%, respectively, both significantly higher than those observed in the UC group. The rEnGAM (22 + 59 + 56-T) group exhibited the most favorable overall performance, with the lowest lesion score (0.30 ± 0.08), the highest antibody level (OD
450 = 1.35 ± 0.05), a 29.84% reduction in oocyst shedding, and the highest anticoccidial index (ACI) of 173.16, collectively indicating a robust and highly effective anticoccidial immune response.
E. necatrix is an intracellular parasite that mainly induces the host cellular immune response. Cytokines are a class of immune factors secreted by immune cells and play an important role in the production and regulation of cellular immune response. Interferon γ (IFN-γ) and Interleukin-2 (IL-2) are classified as Th1 cytokines that could enhance the cellular immune response and mediate delayed hypersensitivity; Interleukin-4 (IL-4) and Interleukin-10 (IL-10) are typical Th2 cytokines, which mainly play a role in enhancing the humoral and allergic immune response [
33]. In this study, no significant differences were observed in the levels of IFN-γ, IL-2, IL-4, or IL-10 across all groups. However, IFN-γ and IL-2 levels exhibited an upward trend following secondary immunization. IL-4 levels increased with the age of the chickens in all groups, while IL-10 levels showed no variation at any time point. Subunit vaccines primarily elicit a humoral immune response through specific antigen fragments, while their ability to stimulate cellular immunity often relies on the careful selection of antigens and the incorporation of appropriate adjuvants [
6]. Although elevated antibody levels serve as a favorable indicator of vaccine-induced immunity, an insufficient cellular immune response remains a critical limiting factor in achieving full protection. Therefore, balancing humoral and cellular immunity is essential for eliciting comprehensive and long-lasting protection. Optimizing vaccine strategies to address this imbalance is imperative. Future research should focus on incorporating immune adjuvants capable of selectively enhancing Th1-type responses to bolster cellular immunity. Moreover, the development of multi-antigen combination immunization approaches may synergistically stimulate both humoral and cellular arms of the immune system, thereby improving overall immunoprotective efficacy.
5. Conclusions
In this study, we successfully cloned, expressed, and characterized the full-length Engam56 gene from E. necatrix, revealing a 1407 bp open reading frame encoding a 468-amino-acid protein with distinct structural and antigenic features. rEnGAM56-F was efficiently expressed in E. coli, purified via Ni-NTA affinity chromatography, and demonstrated strong immunoreactivity in Western blot assays with anti-6 × HIS monoclonal antibody, anti-rEnGAM56-F polyclonal antibody, and convalescent serum from chickens infected with E. necatrix. Immunofluorescence analysis further confirmed its localization to WFB2 in gametocytes and the oocyst wall, implying a functional role in parasite wall biosynthesis. These results suggest that EnGAM56 is a structurally conserved, immunogenic gametocyte antigen with potential relevance in host–parasite interactions and vaccine development.
Vaccination trials demonstrated that rEnGAM56, particularly the truncated version (rEnGAM56-T), conferred partial protection against E. necatrix infection, as evidenced by improved body weight gain, reduced lesion scores, and enhanced anticoccidial index (ACI). Moreover, the multivalent antigen combination including rEnGAM22, rEnGAM59, and rEnGAM56-T elicited the most robust protective immunity, achieving an ACI of 173.16 and significantly elevated IgY levels. These findings underscore the potential of EnGAM56, especially when incorporated into multi-antigen formulations, as a promising component of next-generation subunit vaccines against avian coccidiosis caused by E. necatrix.






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}