Mechanism Analysis of UCP2 During the Oxidative Stress Injury of Intestinal Porcine Epithelial Cell Line-J2

 , , ,
, , ,
Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Construction of Lentiviral Vector Expressing UCP2 Gene
2.2. Production of Lentiviral Particles Carrying the UCP2 Gene
2.3. Establishment of UCP2-Overexpressing Porcine Intestinal Epithelial Cell Line (IPEC-J2-UCP2)
2.4. Cell Viability Assay of IPEC-J2-UCP2 Cells Under Hydrogen Peroxide–Induced Oxidative Stress
2.5. Measuring Intracellular Reactive Oxygen Species Levels Under Oxidative Stress Using Fluorescent Probes
2.6. Evaluation of Antioxidant Enzyme Activities in IPEC-J2-UCP2 Cells Under Oxidative Stress
2.7. Apoptosis-Related Gene Expression Levels Under Oxidative Stress
2.8. Proliferation of IPEC-J2 Cell Under Genipin Intervention
2.9. Data Analysis
3. Results
3.1. Generation and Validation of UCP2-Overexpressing IPEC-J2 Cell Line
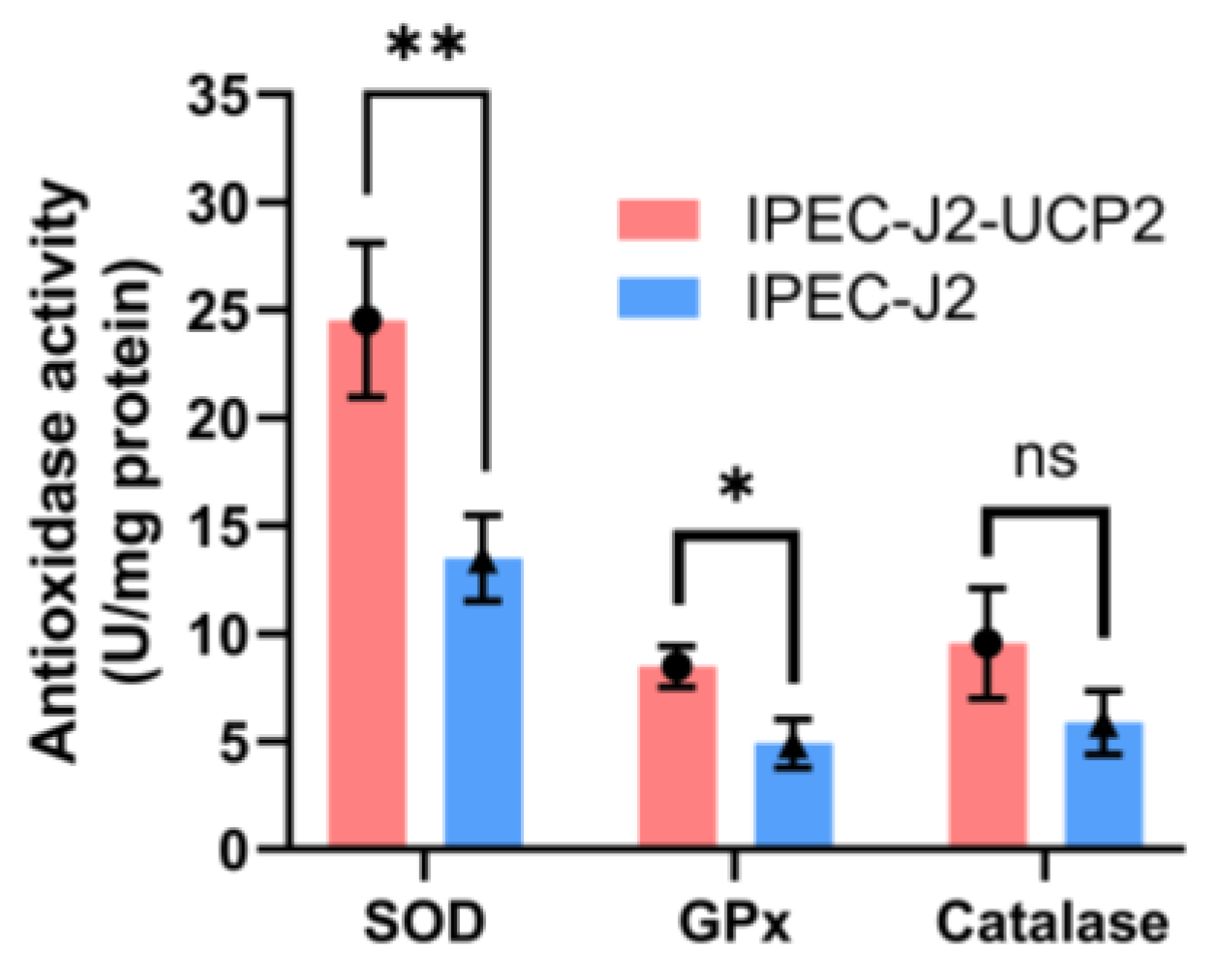
3.2. UCP2 Overexpression Enhances Oxidative Stress Resistance and Modulates Apoptotic Pathways in IPEC-J2 Cells
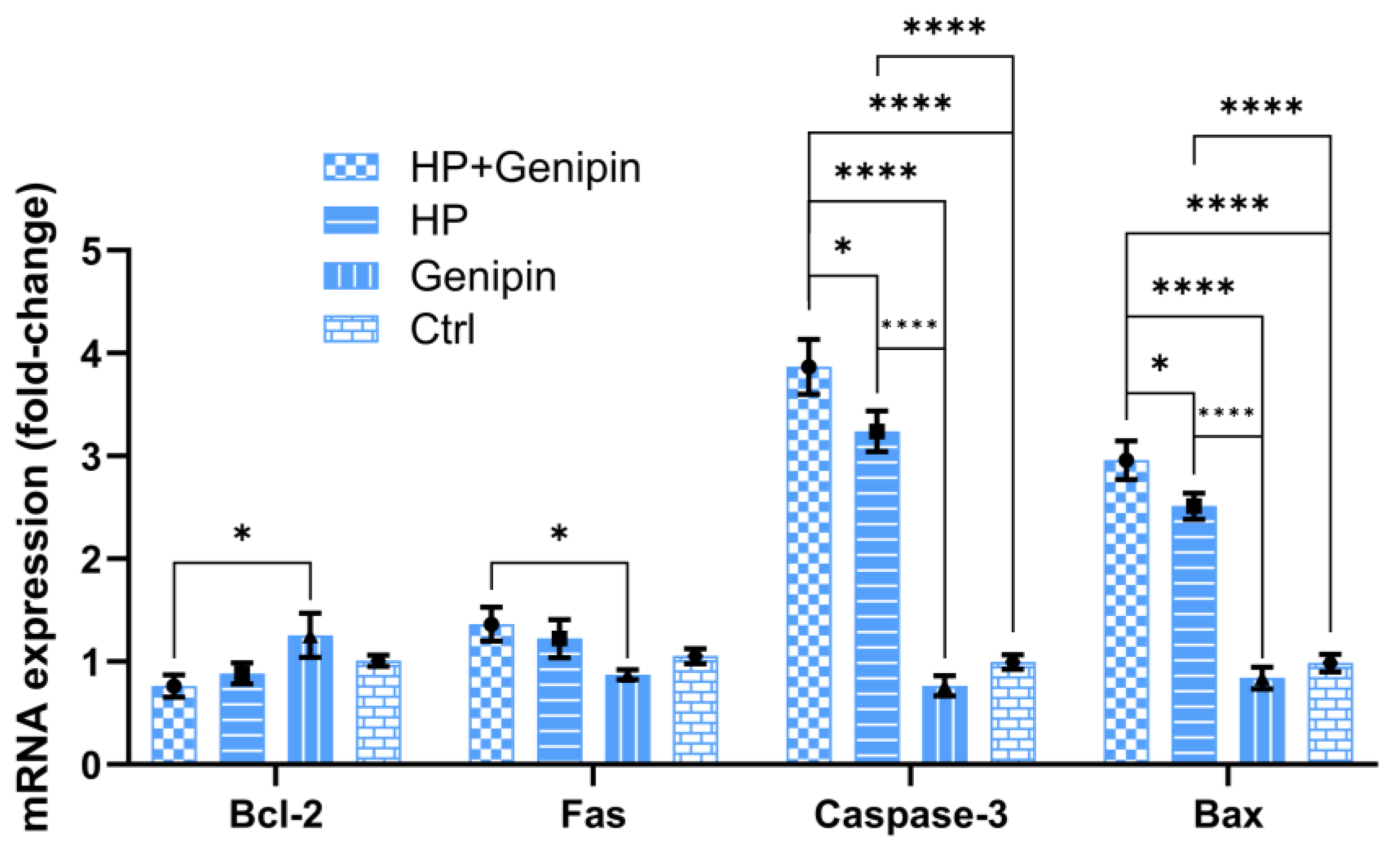
3.3. Genipin Modulates Cell Proliferation, UCP2 Expression, and Apoptotic Pathways in IPEC-J2 Cells Under Oxidative Stress
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Loveikyte, R.; Bourgonje, A.R.; van Goor, H.; Dijkstra, G.; van der Meulen-de Jong, A.E. The effect of iron therapy on oxidative stress and intestinal microbiota in inflammatory bowel diseases: A review on the conundrum. Redox Biol. 2023, 68, 102950. [Google Scholar] [CrossRef] [PubMed]
- Deng, H.; Jia, Q.; Ming, X.; Sun, Y.; Lu, Y.; Liu, L.; Zhou, J. Hippo pathway in intestinal diseases: Focusing on ferroptosis. Front. Cell Dev. Biol. 2023, 11, 1291686. [Google Scholar] [CrossRef] [PubMed]
- An, X.; Zhong, C.; Han, B.; Chen, E.; Zhu, Q.; Yang, Y.; Li, R.; Yang, R.; Zha, D.; Han, Y. Lysophosphatidic acid exerts protective effects on HEI-OC1 cells against cytotoxicity of cisplatin by decreasing apoptosis, excessive autophagy, and accumulation of ROS. Cell Death Discov. 2023, 9, 415. [Google Scholar] [CrossRef]
- Columbus, D.A. 314 Maxwell lecture: Role of amino acids and protein in health-challenged nursery pigs. J. Anim. Sci. 2023, 101, 137. [Google Scholar] [CrossRef]
- Yu, L.-e.; Mann, P.; Schlitzkus, L.; Chakradhar, J.; Li, Y. 56 Early weaning stress relocates epithelial enteroendocrine cells and alters intestinal nutrient sensing and transport functions in pigs. J. Anim. Sci. 2023, 101, 39–40. [Google Scholar] [CrossRef]
- Zarzosa, V.; Lomonte, B.; Zamudio, F.; Ponce-Lopez, R.; Olvera-Rodriguez, F.; Borja, M.; Alagon, A.; Neri-Castro, E. Venom of the neotropical rattlesnake, Crotalus culminatus: Intraspecific variation, neutralization by antivenoms, and immunogenicity in rabbits. Biochimie 2024, 216, 160–174. [Google Scholar] [CrossRef]
- Acevedo-Diaz, A.C.; Caggiano, E.G.; Yu, M.; Taniguchi, C. Abstract 4830: Uncoupling protein 2 (UCP2) loss of function mediates PDAC tumor suppression by mitochondrial fusion. Cancer Res. 2023, 83, 4830. [Google Scholar] [CrossRef]
- Schiffer, T.A.; Lof, L.; Gallini, R.; Kamali-Moghaddam, M.; Carlstrom, M.; Palm, F. Mitochondrial Respiration-Dependent ANT2-UCP2 Interaction. Front. Physiol. 2022, 13, 866590. [Google Scholar] [CrossRef] [PubMed]
- Stenberg, S.; Li, J.; Gjuvsland, A.B.; Persson, K.; Demitz-Helin, E.; Gonzalez Pena, C.; Yue, J.X.; Gilchrist, C.; Arengard, T.; Ghiaci, P.; et al. Genetically controlled mtDNA deletions prevent ROS damage by arresting oxidative phosphorylation. Elife 2022, 11, e76095. [Google Scholar] [CrossRef]
- Jezek, P.; Holendova, B.; Garlid, K.D.; Jaburek, M. Mitochondrial Uncoupling Proteins: Subtle Regulators of Cellular Redox Signaling. Antioxid. Redox Signal. 2018, 29, 667–714. [Google Scholar] [CrossRef]
- Kukat, A.; Dogan, S.A.; Edgar, D.; Mourier, A.; Jacoby, C.; Maiti, P.; Mauer, J.; Becker, C.; Senft, K.; Wibom, R.; et al. Loss of UCP2 attenuates mitochondrial dysfunction without altering ROS production and uncoupling activity. PLoS Genet. 2014, 10, e1004385. [Google Scholar] [CrossRef] [PubMed]
- Geng, Z.; Chen, W.; Lu, Q.; Fu, B.; Fu, X. UCP2 overexpression activates SIRT3 to regulate oxidative stress and mitochondrial dynamics induced by myocardial injury. Arch. Biochem. Biophys. 2024, 753, 109918. [Google Scholar] [CrossRef]
- Choi, S.; Mehrotra, S. UCP2 Deficiency Renders Th1/2 Pathogenic Phenotype to CD4+ T cells. J. Immunol. 2023, 210, 64.11. [Google Scholar] [CrossRef]
- Jin, C.; Zongo, A.W.; Du, H.; Lu, Y.; Yu, N.; Nie, X.; Ma, A.; Ye, Q.; Xiao, H.; Meng, X. Gardenia (Gardenia jasminoides Ellis) fruit: A critical review of its functional nutrients, processing methods, health-promoting effects, comprehensive application and future tendencies. Crit. Rev. Food Sci. Nutr. 2025, 65, 165–192. [Google Scholar] [CrossRef] [PubMed]
- Natallia, L.; Dama, A.; Gorica, E.; Darya, K.; Pena-Corona, S.I.; Cortes, H.; Santini, A.; Busselberg, D.; Leyva-Gomez, G.; Sharifi-Rad, J. Genipin's potential as an anti-cancer agent: From phytochemical origins to clinical prospects. Med. Oncol. 2024, 41, 186. [Google Scholar] [CrossRef]
- Djordjevic, S.; Lebon, D.; Itzykson, R.; Ouled-Haddou, H.; Jedraszak, G.; Harbonnier, J.; Collet, L.; Paubelle, E.; Duployez, N.; Marolleau, J.P. Prognostic value and functional consequences of calcium signalling in acute myeloid leukaemia. Blood 2023, 142, 5678. [Google Scholar] [CrossRef]
- Gupta, A.K.; Roy, S.; Das, P.K. Antileishmanial effect of the natural immunomodulator genipin through suppression of host negative regulatory protein UCP2. J. Antimicrob. Chemother. 2021, 76, 135–145. [Google Scholar] [CrossRef]
- Beikbaghban, T.; Proietti, L.; Ebner, J.; Sango, R.; Rattei, T.; Weichhart, T.; Grebien, F.; Sternberg, F.; Pohl, E.E. Differential regulation of mitochondrial uncoupling protein 2 in cancer cells. Biochim. Biophys. Acta Bioenerg. 2024, 1865, 149486. [Google Scholar] [CrossRef]
- Jin, S.; Wijerathne, C.U.B.; Au-Yeung, K.K.W.; Lei, H.; Yang, C.; O, K. Effects of high- and low-fiber diets on intestinal oxidative stress in growing-finishing pigs. J. Anim. Sci. 2022, 100, skac306. [Google Scholar] [CrossRef]
- Bacou, E.; Walk, C.; Rider, S.; Litta, G.; Perez-Calvo, E. Dietary oxidative distress: A Review of nutritional challenges as models for poultry, swine and fish. Antioxidants 2021, 10, 525. [Google Scholar] [CrossRef]
- Zhou, Y.; Sun, R.; Zhang, Z.W.; He, X.Y.; Li, L.; Zhang, C.J.; Liu, Y.; Yu, H.T. Proliferation Inhibited by Genipin in Human Leukemia K562 Cells: Involvement of Uncoupling Protein 2 in Mitochondrial Damage. World J. Oncol. 2024, 16, 83. [Google Scholar] [CrossRef] [PubMed]
- Kim, G.; Han, D.-W.; Lee, J.H. The Cytoprotective Effects of Baicalein on H2O2-Induced ROS by Maintaining Mitochondrial Homeostasis and Cellular Tight Junction in HaCaT Keratinocytes. Antioxidants 2023, 12, 902. [Google Scholar] [CrossRef] [PubMed]
- Cai, X.; Chen, X.; Wang, X.; Xu, C.; Guo, Q.; Zhu, L.; Zhu, S.; Xu, J. Pre-protective effect of lipoic acid on injury induced by H2O2 in IPEC-J2 cells. Mol. Cell. Biochem. 2013, 378, 73–81. [Google Scholar] [CrossRef] [PubMed]
- Luby, A.; Alves-Guerra, M.C. UCP2 as a cancer target through energy metabolism and oxidative stress control. Int. J. Mol. Sci. 2022, 23, 15077. [Google Scholar] [CrossRef]
- Forte, M.; Bianchi, F.; Cotugno, M.; Marchitti, S.; Stanzione, R.; Maglione, V.; Sciarretta, S.; Valenti, V.; Carnevale, R.; Versaci, F. An interplay between UCP2 and ROS protects cells from high-salt-induced injury through autophagy stimulation. Cell Death Dis. 2021, 12, 919. [Google Scholar] [CrossRef]
- Stanzione, R.; Forte, M.; Cotugno, M.; Bianchi, F.; Marchitti, S.; Busceti, C.L.; Fornai, F.; Rubattu, S. Uncoupling protein 2 as a pathogenic determinant and therapeutic target in cardiovascular and metabolic diseases. Curr. Neuropharmacol. 2022, 20, 662–674. [Google Scholar] [CrossRef]
- Agyekum, A.K.; Torrey, S.; Simard, F.; Abedin, M. PSI-27 Dietary supplementation of betaine and an antioxidant blend improved feed efficiency in commercially housed fattening pigs under heat stress conditions. J. Anim. Sci. 2024, 102, 689–690. [Google Scholar] [CrossRef]
- Durand, D.; Collin, A.; Merlot, E.; Baeza, E.; Guilloteau, L.A.; Le Floc'h, N.; Thomas, A.; Fontagne-Dicharry, S.; Gondret, F. Review: Implication of redox imbalance in animal health and performance at critical periods, insights from different farm species. Animal 2022, 16, 100543. [Google Scholar] [CrossRef]
- Zhou, D.; Zhuan, Q.; Luo, Y.; Liu, H.; Meng, L.; Du, X.; Wu, G.; Hou, Y.; Li, J.; Fu, X. Mito-Q promotes porcine oocytes maturation by maintaining mitochondrial thermogenesis via UCP2 downregulation. Theriogenology 2022, 187, 205–214. [Google Scholar] [CrossRef]
- Wang, Y.; Lv, J.; Liu, G.; Yao, Q.; Wang, Z.; Liu, N.; He, Y.; Il, D.; Tusupovich, J.I.; Jiang, Z. ZnO NPs Impair the Viability and Function of Porcine Granulosa Cells Through Autophagy Regulated by ROS Production. Antioxidants 2024, 13, 1295. [Google Scholar] [CrossRef]
- Guo, Z.; Lv, L.; Liu, D.; Ma, H.; Wang, L.; Fu, B.; Wang, F. Network meta-analysis: Effect of cold stress on the gene expression of swine adipocytes ATGL, CIDEA, UCP2, and UCP3. Curr. Issues Mol. Biol. 2024, 46, 3866–3876. [Google Scholar] [CrossRef] [PubMed]
- Jena, A.B.; Samal, R.R.; Bhol, N.K.; Duttaroy, A.K. Cellular Red-Ox system in health and disease: The latest update. Biomed. Pharmacother. 2023, 162, 114606. [Google Scholar] [CrossRef] [PubMed]
- Wang, L.; Li, X.; Chen, L.; Mei, S.; Shen, Q.; Liu, L.; Liu, X.; Liao, S.; Zhao, B.; Chen, Y. Mitochondrial uncoupling protein-2 ameliorates ischemic stroke by inhibiting ferroptosis-induced brain injury and neuroinflammation. Mol. Neurobiol. 2025, 62, 501–517. [Google Scholar] [CrossRef] [PubMed]
- Liu, W.; He, W.; Li, H. Exhaustive Training Increases Uncoupling Protein 2 Expression and Decreases Bcl-2/Bax Ratio in Rat Skeletal Muscle. Oxidative Med. Cell. Longev. 2013, 2013, 780719. [Google Scholar] [CrossRef] [PubMed]
- Caggiano, E.G.; Taniguchi, C.M. UCP2 and pancreatic cancer: Conscious uncoupling for therapeutic effect. Cancer Metastasis Rev. 2024, 43, 777–794. [Google Scholar] [CrossRef]
- Li, J.; Lim, J.Y.S.; Eu, J.Q.; Chan, A.K.M.H.; Goh, B.C.; Wang, L.; Wong, A.L.-A. Reactive oxygen species modulation in the current landscape of anticancer therapies. Antioxid. Redox Signal. 2024, 41, 322–341. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Product (bp) | Sequence (5′–3′) | GenBank Accession No. | Gene Name |
|---|---|---|---|
| 181 | F: GGACCTGACCGACTACCTCAT | DQ452569 | β-actin |
| R: GGGCAGCTCGTAGCTCTTCT | |||
| 180 | F: ACCTGAATGACCACCTAGAGC | NM_214285 | Bcl-2 |
| R: TCCGACTGAAGAGCGAAC | |||
| 103 | F: TGATGCCCAAGTGACTGACC | NM_213839 | Fas |
| R: GCAGAATTGACCCTCACGAT | |||
| 190 | F: GTGGGACTGAAGATGACA | NM_214131 | Caspase-3 |
| R: ACCCGAGTAAGAATGTG | |||
| 154 | F: CCGAAATGTTTGCTGACG | XM_003127290 | Bax |
| R: AGCCGATCTCGAAGGAAGT |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Su, W.; Xu, C.; Jiang, H.; Song, W.; Xiong, P.; Chen, J.; Ai, G.; Song, Q.; Zou, Z.; Wei, Q.; et al. Mechanism Analysis of UCP2 During the Oxidative Stress Injury of Intestinal Porcine Epithelial Cell Line-J2. Animals 2025, 15, 1654. https://doi.org/10.3390/ani15111654
Su W, Xu C, Jiang H, Song W, Xiong P, Chen J, Ai G, Song Q, Zou Z, Wei Q, et al. Mechanism Analysis of UCP2 During the Oxidative Stress Injury of Intestinal Porcine Epithelial Cell Line-J2. Animals. 2025; 15(11):1654. https://doi.org/10.3390/ani15111654
Chicago/Turabian StyleSu, Weide, Chuanhui Xu, Hongping Jiang, Wenjing Song, Pingwen Xiong, Jiang Chen, Gaoxiang Ai, Qiongli Song, Zhiheng Zou, Qipeng Wei, and et al. 2025. "Mechanism Analysis of UCP2 During the Oxidative Stress Injury of Intestinal Porcine Epithelial Cell Line-J2" Animals 15, no. 11: 1654. https://doi.org/10.3390/ani15111654
APA StyleSu, W., Xu, C., Jiang, H., Song, W., Xiong, P., Chen, J., Ai, G., Song, Q., Zou, Z., Wei, Q., & Chen, X. (2025). Mechanism Analysis of UCP2 During the Oxidative Stress Injury of Intestinal Porcine Epithelial Cell Line-J2. Animals, 15(11), 1654. https://doi.org/10.3390/ani15111654