
The cAMP Receptor Protein (CRP) of Vibrio mimicus Regulates Its Bacterial Growth, Type II Secretion System, Flagellum Formation, Adhesion Genes, and Virulence
 ,
, {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
Simple Summary
Abstract
1. Introduction
2. Material and Methods
2.1. Bacterial Strains, Plasmids, and Growth Conditions
2.2. Construction of Deletion and Complementation Strains
2.3. Growth Assay
2.4. Hemolytic Activity Assay
2.5. Congo Red Binding Assays
2.6. Microtiter Dish Biofilm Formation Assay
2.7. Swimming Motility Assay and Transmission Electron Microscopy (TEM) Observation
2.8. Cell Adhesion Assay
2.9. Enzymatic Activity Assay
2.10. Cell Cytotoxicity Assay
2.11. Pathogenicity Assay in the Catfish Model
2.12. Quantitative Reverse Transcription Polymerase Chain Reaction (qRT-PCR) Analysis
2.13. Statistical Analysis
3. Results
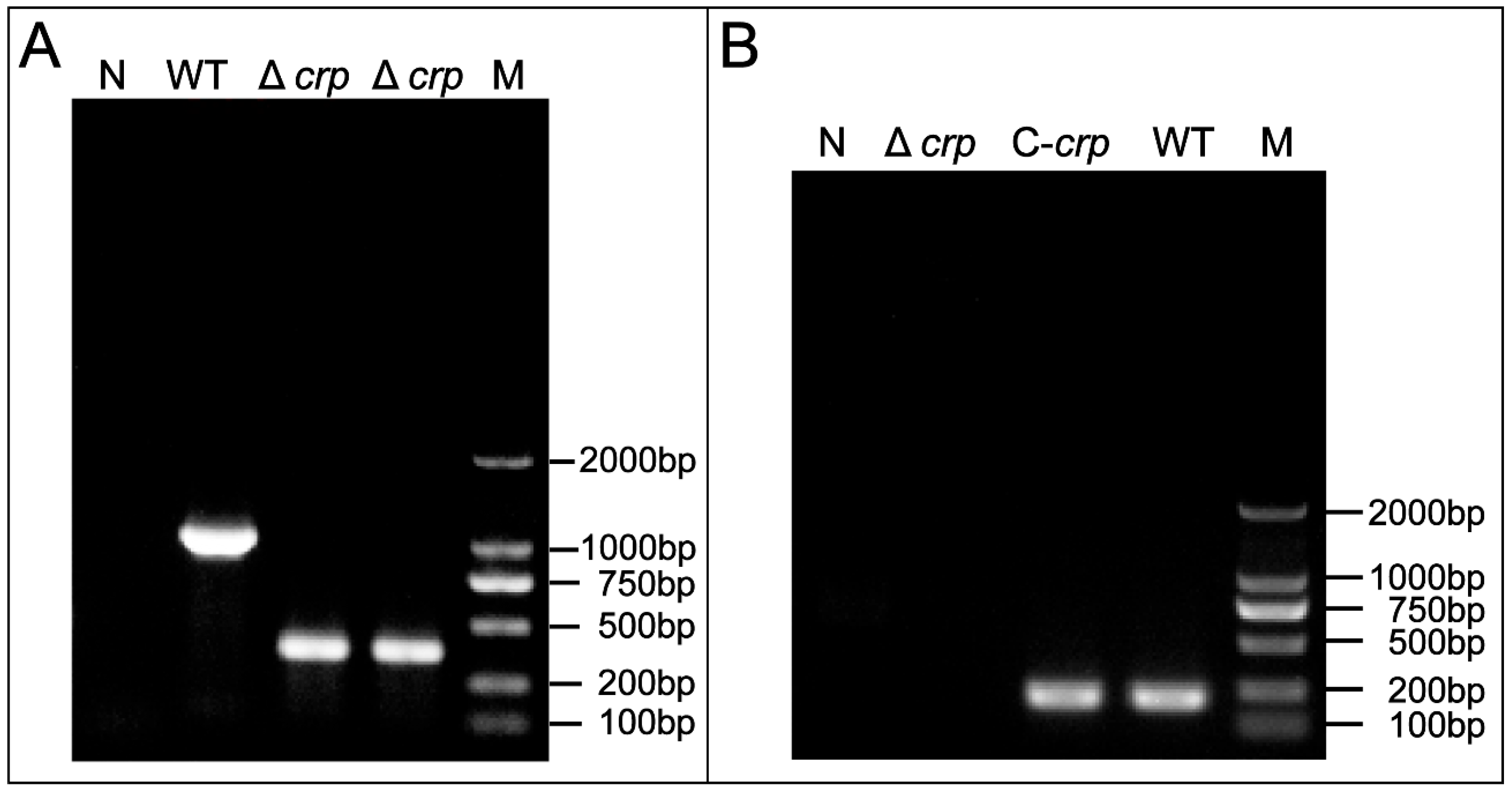
3.1. Construction and Detection of the V. mimicus Deletion Strain Δcrp and Complementary Strain C-crp
3.2. Effect of crp Deletion on Growth and Morphology
3.3. Effect of crp Deletion on Type II Secretion System
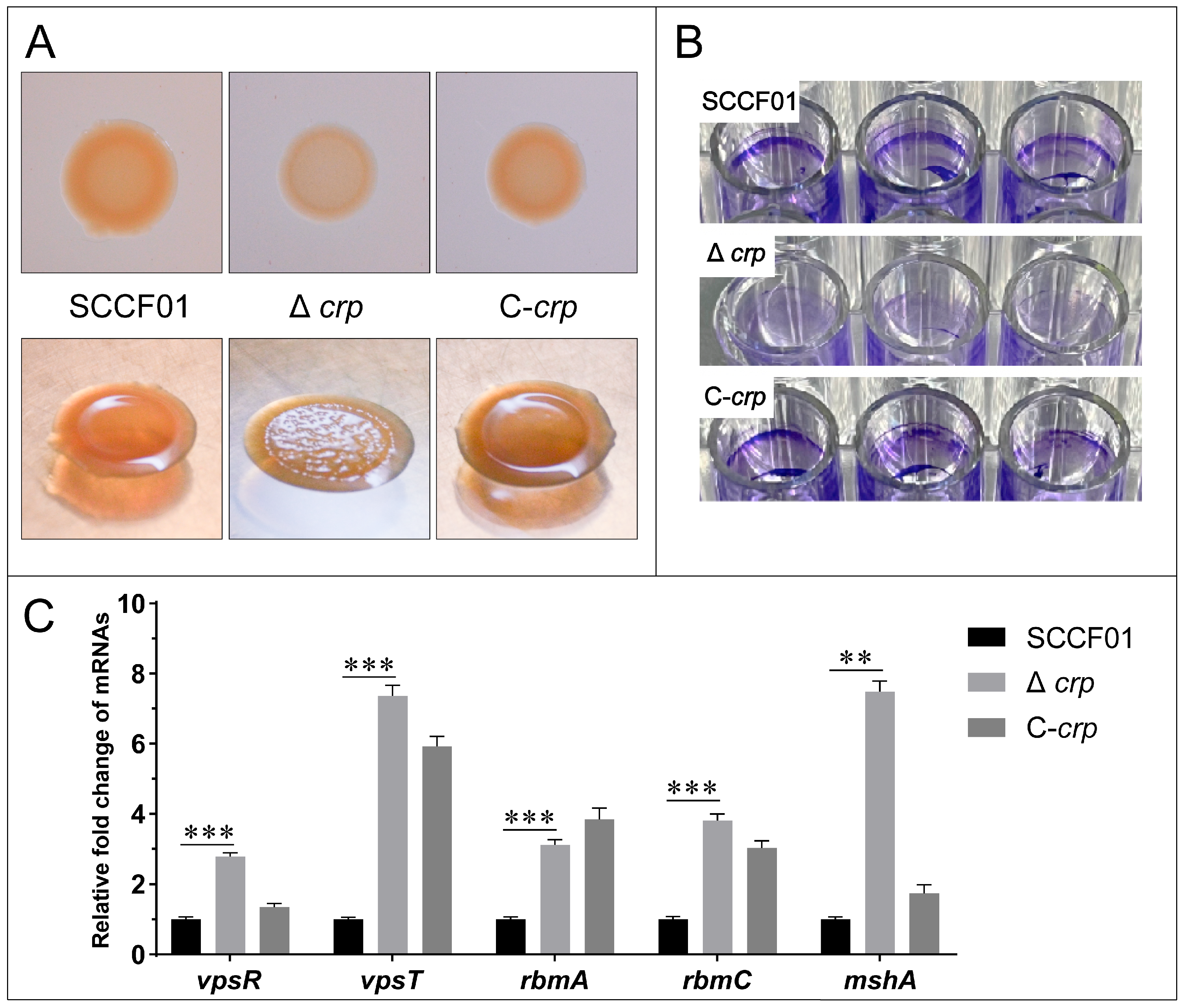
3.4. Effect of the crp Gene on Colonial Morphology and Biofilm Formation Ability
3.5. Effect of the crp Gene on Swimming Motility and Adhesion Ability
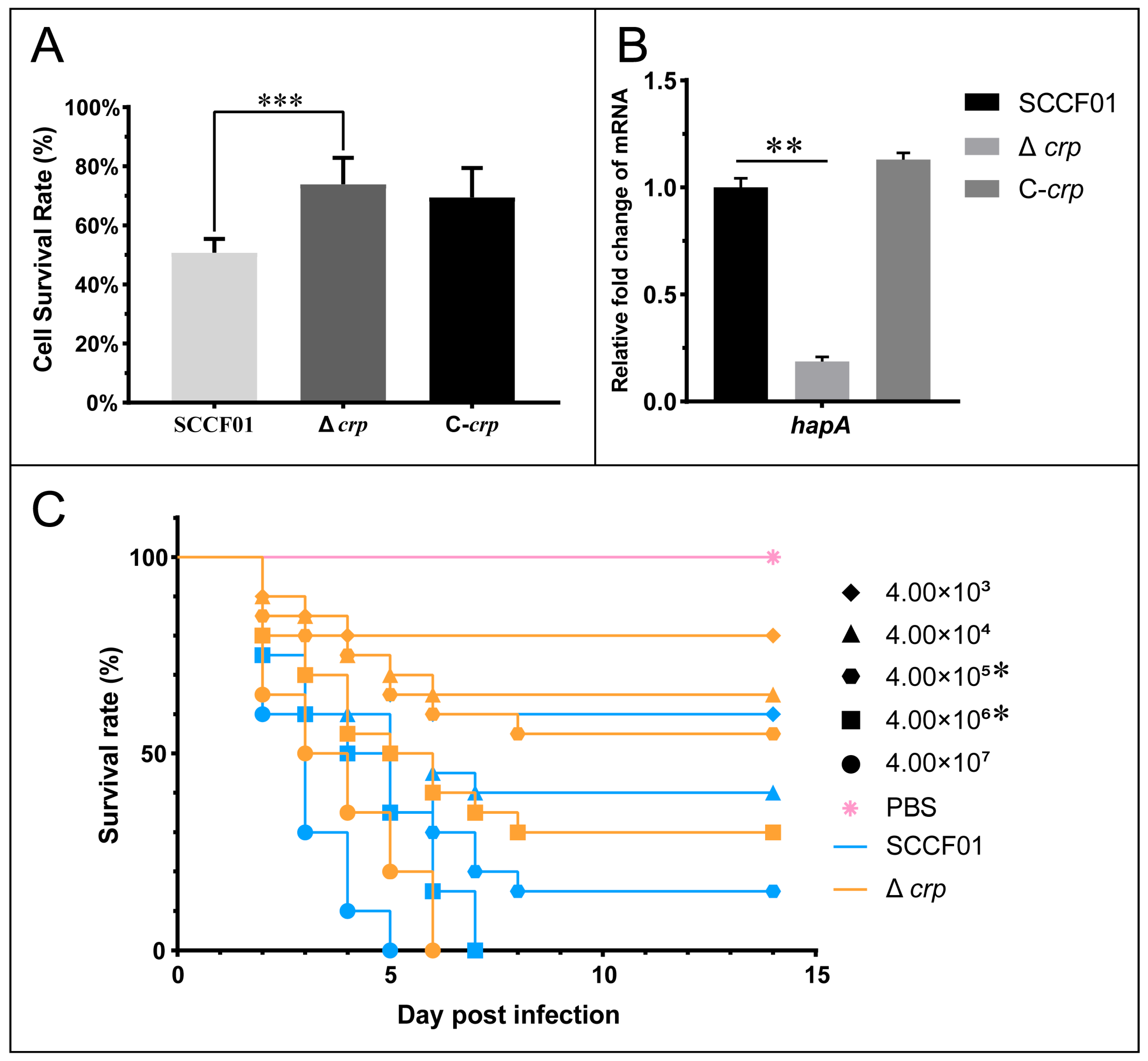
3.6. Pathogenicity of the crp Deletion Strain in Cells and Animals
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Davis, B.R.; Fanning, G.R.; Madden, J.M.; Steigerwalt, A.G.; Bradford, H.B., Jr.; Smith, H.L., Jr.; Brenner, D.J. Characterization of biochemically atypical cholerae strains and designation of a new pathogenic species, Vibrio mimicus. J. Clin. Microbiol. 1981, 14, 631–639. [Google Scholar] [CrossRef]
- Cen, J.; Liu, X.; Li, J.; Zhang, M.; Wang, W. Identification and immunogenicity of immunodominant mimotopes of outer membrane protein U (OmpU) of Vibrio mimicus from phage display peptide library. Fish Shellfish Immunol. 2013, 34, 291–295. [Google Scholar] [CrossRef]
- Prabina, D.; Swaminathan, T.R.; Mohandas, S.P.; Anjana, J.C.; Manjusha, K.; Preena, P.G. Investigation of antibiotic-resistant vibrios associated with shrimp (Penaeus vannamei) farms. Arch. Microbiol. 2022, 205, 41. [Google Scholar] [CrossRef] [PubMed]
- Tercero-Alburo, J.J.; González-Márquez, H.; Bonilla-González, E.; Quiñones-Ramírez, E.I.; Vázquez-Salinas, C. Identification of capsule, biofilm, lateral flagellum, and type IV pili in Vibrio mimicus strains. Microb. Pathog. 2014, 76, 77–83. [Google Scholar] [CrossRef]
- Hernández-Robles, M.F.; Natividad-Bonifacio, I.; Álvarez-Contreras, A.K.; Tercero-Alburo, J.J.; Quiñones-Ramírez, E.I.; Vázquez-Salinas, C. Characterization of Potential Virulence Factors of Vibrio mimicus Isolated from Fishery Products and Water. Int. J. Microbiol. 2021, 2021, 8397930. [Google Scholar] [CrossRef]
- Zhang, X.; Li, Y.-W.; Mo, Z.-Q.; Luo, X.-C.; Sun, H.-Y.; Liu, P.; Li, A.-X.; Zhou, S.-M.; Dan, X.-M. Outbreak of a novel disease associated with Vibrio mimicus infection in fresh water cultured yellow catfish, Pelteobagrus fulvidraco. Aquaculture 2014, 432, 119–124. [Google Scholar] [CrossRef]
- Yu, Z.; Wang, E.; Geng, Y.; Wang, K.; Chen, D.; Huang, X.; Ouyang, P.; Zuo, Z.; Huang, C.; Fang, J.; et al. Complete genome analysis of Vibrio mimicus strain SCCF01, a highly virulent isolate from the freshwater catfish. Virulence 2020, 11, 23–31. [Google Scholar] [CrossRef]
- Lin, L.; Feng, D.; Pan, X.; Yao, J.; Yin, W.; Cao, Z.; Liu, Y.; Xia, Y.; Shen, J. Identification, virulence-related factors, and antimicrobial susceptibility of Vibrio mimicus from yellow catfish, Pelteobagrus fulvidraco. Acta Hydrobiol. Sin. 2020, 44, 799–810. [Google Scholar] [CrossRef]
- Geng, Y.; Liu, D.; Han, S.; Zhou, Y.; Wang, K.Y.; Huang, X.L.; Chen, D.F.; Peng, X.; Lai, W.M. Outbreaks of vibriosis associated with Vibrio mimicus in freshwater catfish in China. Aquaculture 2014, 433, 82–84. [Google Scholar] [CrossRef]
- Colton, D.M.; Stabb, E.V. Rethinking the roles of CRP, cAMP, and sugar-mediated global regulation in the Vibrionaceae. Curr. Genet. 2016, 62, 39–45. [Google Scholar] [CrossRef]
- Shimada, T.; Fujita, N.; Yamamoto, K.; Ishihama, A. Novel roles of cAMP receptor protein (CRP) in regulation of transport and metabolism of carbon sources. PLoS ONE 2011, 6, e20081. [Google Scholar] [CrossRef] [PubMed]
- Green, J.; Stapleton, M.R.; Smith, L.J.; Artymiuk, P.J.; Kahramanoglou, C.; Hunt, D.M.; Buxton, R.S. Cyclic-AMP and bacterial cyclic-AMP receptor proteins revisited: Adaptation for different ecological niches. Curr. Opin. Microbiol. 2014, 18, 1–7. [Google Scholar] [CrossRef] [PubMed]
- Petersen, S.; Young, G.M. Essential role for cyclic AMP and its receptor protein in Yersinia enterocolitica virulence. Infect. Immun. 2002, 70, 3665–3672. [Google Scholar] [CrossRef] [PubMed]
- Zhan, L.; Han, Y.; Yang, L.; Geng, J.; Li, Y.; Gao, H.; Guo, Z.; Fan, W.; Li, G.; Zhang, L.; et al. The cyclic AMP receptor protein, CRP, is required for both virulence and expression of the minimal CRP regulon in Yersinia pestis biovar microtus. Infect. Immun. 2008, 76, 5028–5037. [Google Scholar] [CrossRef] [PubMed]
- Zhao, X.; Liu, Q.; Xiao, K.; Hu, Y.; Liu, X.; Li, Y.; Kong, Q. Identification of the crp gene in avian Pasteurella multocida and evaluation of the effects of crp deletion on its phenotype, virulence and immunogenicity. BMC Microbiol. 2016, 16, 125. [Google Scholar] [CrossRef] [PubMed]
- Santander, J.; Mitra, A.; Curtiss, R., 3rd. Phenotype, virulence and immunogenicity of Edwardsiella ictaluri cyclic adenosine 3′,5′-monophosphate receptor protein (Crp) mutants in catfish host. Fish Shellfish Immunol. 2011, 31, 1142–1153. [Google Scholar] [CrossRef]
- Prüß, B.M. Involvement of Two-Component Signaling on Bacterial Motility and Biofilm Development. J. Bacteriol. 2017, 199, e00259-17. [Google Scholar] [CrossRef]
- Fong, J.C.; Yildiz, F.H. Interplay between cyclic AMP-cyclic AMP receptor protein and cyclic di-GMP signaling in Vibrio cholerae biofilm formation. J. Bacteriol. 2008, 190, 6646–6659. [Google Scholar] [CrossRef]
- Manneh-Roussel, J.; Haycocks, J.R.J.; Magán, A.; Perez-Soto, N.; Voelz, K.; Camilli, A.; Krachler, A.M.; Grainger, D.C. cAMP Receptor Protein Controls Vibrio cholerae Gene Expression in Response to Host Colonization. mBio 2018, 9, e00966-18. [Google Scholar] [CrossRef]
- Kim, S.P.; Kim, C.M.; Shin, S.H. Cyclic AMP and cyclic AMP-receptor protein modulate the autoinducer-2-mediated quorum sensing system in Vibrio vulnificus. Curr. Microbiol. 2012, 65, 701–710. [Google Scholar] [CrossRef]
- Blokesch, M. Chitin colonization, chitin degradation and chitin-induced natural competence of Vibrio cholerae are subject to catabolite repression. Environ. Microbiol. 2012, 14, 1898–1912. [Google Scholar] [CrossRef]
- Lee, J.H.; Choi, S.H. Coactivation of Vibrio vulnificus putAP operon by cAMP receptor protein and PutR through cooperative binding to overlapping sites. Mol. Microbiol. 2006, 60, 513–524. [Google Scholar] [CrossRef]
- Purcell, A.B.; Simpson, B.W.; Trent, M.S. Impact of the cAMP-cAMP Receptor Protein Regulatory Complex on Lipopolysaccharide Modifications and Polymyxin B Resistance in Escherichia coli. J. Bacteriol. 2023, 205, e0006723. [Google Scholar] [CrossRef]
- Zahid, M.S.; Waise, T.M.; Kamruzzaman, M.; Ghosh, A.N.; Nair, G.B.; Mekalanos, J.J.; Faruque, S.M. The cyclic AMP (cAMP)-cAMP receptor protein signaling system mediates resistance of Vibrio cholerae O1 strains to multiple environmental bacteriophages. Appl. Environ. Microbiol. 2010, 76, 4233–4240. [Google Scholar] [CrossRef]
- Baharoglu, Z.; Krin, E.; Mazel, D. Connecting environment and genome plasticity in the characterization of transformation-induced SOS regulation and carbon catabolite control of the Vibrio cholerae integron integrase. J. Bacteriol. 2012, 194, 1659–1667. [Google Scholar] [CrossRef]
- Yu, Z.; Geng, Y.; Wang, K.; Chen, D.; Huang, X.; Ou, Y.; Peng, G. Complete genome sequence of Vibrio mimicus strain SCCF01 with potential application in fish vaccine development. Virulence 2017, 8, 1028–1030. [Google Scholar] [CrossRef]
- Datsenko, K.A.; Wanner, B.L. One-step inactivation of chromosomal genes in Escherichia coli K-12 using PCR products. Proc. Natl. Acad. Sci. USA 2000, 97, 6640–6645. [Google Scholar] [CrossRef]
- Cherepanov, P.P.; Wackernagel, W. Gene disruption in Escherichia coli: TcR and KmR cassettes with the option of Flp-catalyzed excision of the antibiotic-resistance determinant. Gene 1995, 158, 9–14. [Google Scholar] [CrossRef] [PubMed]
- Guzman, L.M.; Belin, D.; Carson, M.J.; Beckwith, J. Tight regulation, modulation, and high-level expression by vectors containing the arabinose PBAD promoter. J. Bacteriol. 1995, 177, 4121–4130. [Google Scholar] [CrossRef] [PubMed]
- Yu, Z.; Wang, E.; Geng, Y.; Wang, K.; Chen, D.; Huang, X.; Ouyang, P.; Zuo, Z.; He, C.; Tang, L.; et al. Multiplex genome editing by natural transformation in Vibrio mimicus with potential application in attenuated vaccine development. Fish Shellfish Immunol. 2019, 92, 377–383. [Google Scholar] [CrossRef] [PubMed]
- Zhao, R.; Qin, Z.; Feng, Y.; Geng, Y.; Huang, X.; Ouyang, P.; Chen, D.; Guo, H.; Deng, H.; Fang, J.; et al. Sialic acid catabolism contributes to Vibrio mimicus virulence. Aquaculture 2023, 574, 739660. [Google Scholar] [CrossRef]
- Sales, A.E.; de Souza, F.A.; Teixeira, J.A.; Porto, T.S.; Porto, A.L. Integrated process production and extraction of the fibrinolytic protease from Bacillus sp. UFPEDA 485. Appl. Biochem. Biotechnol. 2013, 170, 1676–1688. [Google Scholar] [CrossRef]
- Han, L.; Yuan, J.; Ao, X.; Lin, S.; Han, X.; Ye, H. Biochemical Characterization and Phylogenetic Analysis of the Virulence Factor Lysine Decarboxylase from Vibrio vulnificus. Front. Microbiol. 2018, 9, 3082. [Google Scholar] [CrossRef]
- Park, J.Y.; Kang, B.R.; Ryu, C.M.; Anderson, A.J.; Kim, Y.C. Polyamine is a critical determinant of Pseudomonas chlororaphis O6 for GacS-dependent bacterial cell growth and biocontrol capacity. Mol. Plant Pathol. 2018, 19, 1257–1266. [Google Scholar] [CrossRef]
- Guerra, P.R.; Herrero-Fresno, A.; Pors, S.E.; Ahmed, S.; Wang, D.; Thøfner, I.; Antenucci, F.; Olsen, J.E. The membrane transporter PotE is required for virulence in avian pathogenic Escherichia coli (APEC). Vet. Microbiol. 2018, 216, 38–44. [Google Scholar] [CrossRef] [PubMed]
- Sawant, K.; Shashidhar, R. The cAMP receptor protein (CRP) enhances the competitive nature of Salmonella Typhimurium. Arch. Microbiol. 2023, 205, 197. [Google Scholar] [CrossRef] [PubMed]
- Liu, L.; Li, F.; Xu, L.; Wang, J.; Li, M.; Yuan, J.; Wang, H.; Yang, R.; Li, B. Cyclic AMP-CRP Modulates the Cell Morphology of Klebsiella pneumoniae in High-Glucose Environment. Front. Microbiol. 2019, 10, 2984. [Google Scholar] [CrossRef] [PubMed]
- Sloup, R.E.; Konal, A.E.; Severin, G.B.; Korir, M.L.; Bagdasarian, M.M.; Bagdasarian, M.; Waters, C.M. Cyclic Di-GMP and VpsR Induce the Expression of Type II Secretion in Vibrio cholerae. J. Bacteriol. 2017, 199, e00106-17. [Google Scholar] [CrossRef] [PubMed]
- Korotkov, K.V.; Krumm, B.; Bagdasarian, M.; Hol, W.G.J. Structural and Functional Studies of EpsC, a Crucial Component of the Type 2 Secretion System from Vibrio cholerae. J. Mol. Biol. 2006, 363, 311–321. [Google Scholar] [CrossRef] [PubMed]
- Robien, M.A.; Krumm, B.E.; Sandkvist, M.; Hol, W.G.J. Crystal Structure of the Extracellular Protein Secretion NTPase EpsE of Vibrio cholerae. J. Mol. Biol. 2003, 333, 657–674. [Google Scholar] [CrossRef] [PubMed]
- Chami, M.; Guilvout, I.; Gregorini, M.; Rémigy, H.W.; Müller, S.A.; Valerio, M.; Engel, A.; Pugsley, A.P.; Bayan, N. Structural Insights into the Secretin PulD and Its Trypsin-resistant Core. J. Biol. Chem. 2005, 280, 37732–37741. [Google Scholar] [CrossRef] [PubMed]
- Lee, H.M.; Chen, J.R.; Lee, H.L.; Leu, W.M.; Chen, L.Y.; Hu, N.T. Functional dissection of the XpsN (GspC) protein of the Xanthomonas campestris pv. campestris type II secretion machinery. J. Bacteriol. 2004, 186, 2946–2955. [Google Scholar] [CrossRef] [PubMed]
- Fong, J.C.; Yildiz, F.H. The rbmBCDEF gene cluster modulates development of rugose colony morphology and biofilm formation in Vibrio cholerae. J. Bacteriol. 2007, 189, 2319–2330. [Google Scholar] [CrossRef] [PubMed]
- Teschler, J.K.; Zamorano-Sánchez, D.; Utada, A.S.; Warner, C.J.; Wong, G.C.; Linington, R.G.; Yildiz, F.H. Living in the matrix: Assembly and control of Vibrio cholerae biofilms. Nat. Rev. Microbiol. 2015, 13, 255–268. [Google Scholar] [CrossRef]
- Sinha-Ray, S.; Ali, A. Mutation in flrA and mshA Genes of Vibrio cholerae Inversely Involved in vps-Independent Biofilm Driving Bacterium Toward Nutrients in Lake Water. Front. Microbiol. 2017, 8, 1770. [Google Scholar] [CrossRef] [PubMed]
- Wang, L.; Ling, Y.; Jiang, H.; Qiu, Y.; Qiu, J.; Chen, H.; Yang, R.; Zhou, D. AphA is required for biofilm formation, motility, and virulence in pandemic Vibrio parahaemolyticus. Int. J. Food Microbiol. 2013, 160, 245–251. [Google Scholar] [CrossRef]
- Su, Y.C.; Liu, C. Vibrio parahaemolyticus: A concern of seafood safety. Food Microbiol. 2007, 24, 549–558. [Google Scholar] [CrossRef] [PubMed]
- Zhu, S.; Kojima, S.; Homma, M. Structure, gene regulation and environmental response of flagella in Vibrio. Front. Microbiol. 2013, 4, 410. [Google Scholar] [CrossRef]
- Moisi, M.; Jenul, C.; Butler, S.M.; New, A.; Tutz, S.; Reidl, J.; Klose, K.E.; Camilli, A.; Schild, S. A novel regulatory protein involved in motility of Vibrio cholerae. J. Bacteriol. 2009, 191, 7027–7038. [Google Scholar] [CrossRef]
- Klose, K.E.; Mekalanos, J.J. Distinct roles of an alternative sigma factor during both free-swimming and colonizing phases of the Vibrio cholerae pathogenic cycle. Mol. Microbiol. 1998, 28, 501–520. [Google Scholar] [CrossRef]
- Correa, N.E.; Lauriano, C.M.; McGee, R.; Klose, K.E. Phosphorylation of the flagellar regulatory protein FlrC is necessary for Vibrio cholerae motility and enhanced colonization. Mol. Microbiol. 2000, 35, 743–755. [Google Scholar] [CrossRef]
- Correa, N.E.; Barker, J.R.; Klose, K.E. The Vibrio cholerae FlgM homologue is an anti-sigma28 factor that is secreted through the sheathed polar flagellum. J. Bacteriol. 2004, 186, 4613–4619. [Google Scholar] [CrossRef]
- Klose, K.E.; Mekalanos, J.J. Differential regulation of multiple flagellins in Vibrio cholerae. J. Bacteriol. 1998, 180, 303–316. [Google Scholar] [CrossRef]
- Liang, W.; Pascual-Montano, A.; Silva, A.J.; Benitez, J.A. The cyclic AMP receptor protein modulates quorum sensing, motility and multiple genes that affect intestinal colonization in Vibrio cholerae. Microbiology 2007, 153, 2964–2975. [Google Scholar] [CrossRef] [PubMed]
- Luo, G.; Huang, L.; Su, Y.; Qin, Y.; Xu, X.; Zhao, L.; Yan, Q. flrA, flrB and flrC regulate adhesion by controlling the expression of critical virulence genes in Vibrio alginolyticus. Emerg. Microbes Infect. 2016, 5, e85. [Google Scholar] [CrossRef]
- Sperandio, V.; Girón, J.A.; Silveira, W.D.; Kaper, J.B. The OmpU outer membrane protein, a potential adherence factor of Vibrio cholerae. Infect. Immun. 1995, 63, 4433–4438. [Google Scholar] [CrossRef] [PubMed]
- Valiente, E.; Davies, C.; Mills, D.C.; Getino, M.; Ritchie, J.M.; Wren, B.W. Vibrio cholerae accessory colonisation factor AcfC: A chemotactic protein with a role in hyperinfectivity. Sci. Rep. 2018, 8, 8390. [Google Scholar] [CrossRef] [PubMed]
- Benitez, J.A.; Silva, A.J. Vibrio cholerae hemagglutinin(HA)/protease: An extracellular metalloprotease with multiple pathogenic activities. Toxicon 2016, 115, 55–62. [Google Scholar] [CrossRef]
- Zhou, P.; Han, X.; Ye, X.; Zheng, F.; Yan, T.; Xie, Q.; Zhang, Y.A.; Curtiss Iii, R.; Zhou, Y. Phenotype, Virulence and Immunogenicity of Edwardsiella piscicida Cyclic AMP Receptor Protein (Crp) Mutants in Catfish Host. Microorganisms 2020, 8, 517. [Google Scholar] [CrossRef] [PubMed]






Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Tian, Z.; Xiang, F.; Peng, K.; Qin, Z.; Feng, Y.; Huang, B.; Ouyang, P.; Huang, X.; Chen, D.; Lai, W.; et al. The cAMP Receptor Protein (CRP) of Vibrio mimicus Regulates Its Bacterial Growth, Type II Secretion System, Flagellum Formation, Adhesion Genes, and Virulence. Animals 2024, 14, 437. https://doi.org/10.3390/ani14030437
Tian Z, Xiang F, Peng K, Qin Z, Feng Y, Huang B, Ouyang P, Huang X, Chen D, Lai W, et al. The cAMP Receptor Protein (CRP) of Vibrio mimicus Regulates Its Bacterial Growth, Type II Secretion System, Flagellum Formation, Adhesion Genes, and Virulence. Animals. 2024; 14(3):437. https://doi.org/10.3390/ani14030437
Chicago/Turabian StyleTian, Ziqi, Fei Xiang, Kun Peng, Zhenyang Qin, Yang Feng, Bowen Huang, Ping Ouyang, Xiaoli Huang, Defang Chen, Weimin Lai, and et al. 2024. "The cAMP Receptor Protein (CRP) of Vibrio mimicus Regulates Its Bacterial Growth, Type II Secretion System, Flagellum Formation, Adhesion Genes, and Virulence" Animals 14, no. 3: 437. https://doi.org/10.3390/ani14030437
APA StyleTian, Z., Xiang, F., Peng, K., Qin, Z., Feng, Y., Huang, B., Ouyang, P., Huang, X., Chen, D., Lai, W., & Geng, Y. (2024). The cAMP Receptor Protein (CRP) of Vibrio mimicus Regulates Its Bacterial Growth, Type II Secretion System, Flagellum Formation, Adhesion Genes, and Virulence. Animals, 14(3), 437. https://doi.org/10.3390/ani14030437