
Age-Specific Demographic Response of a Long-Lived Scavenger Species to Reduction of Organic Matter in a Landfill


Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Study Area and Sampling
2.2. Ageing of Vultures and Sampling Interval
2.3. Apparent Survival Analysis
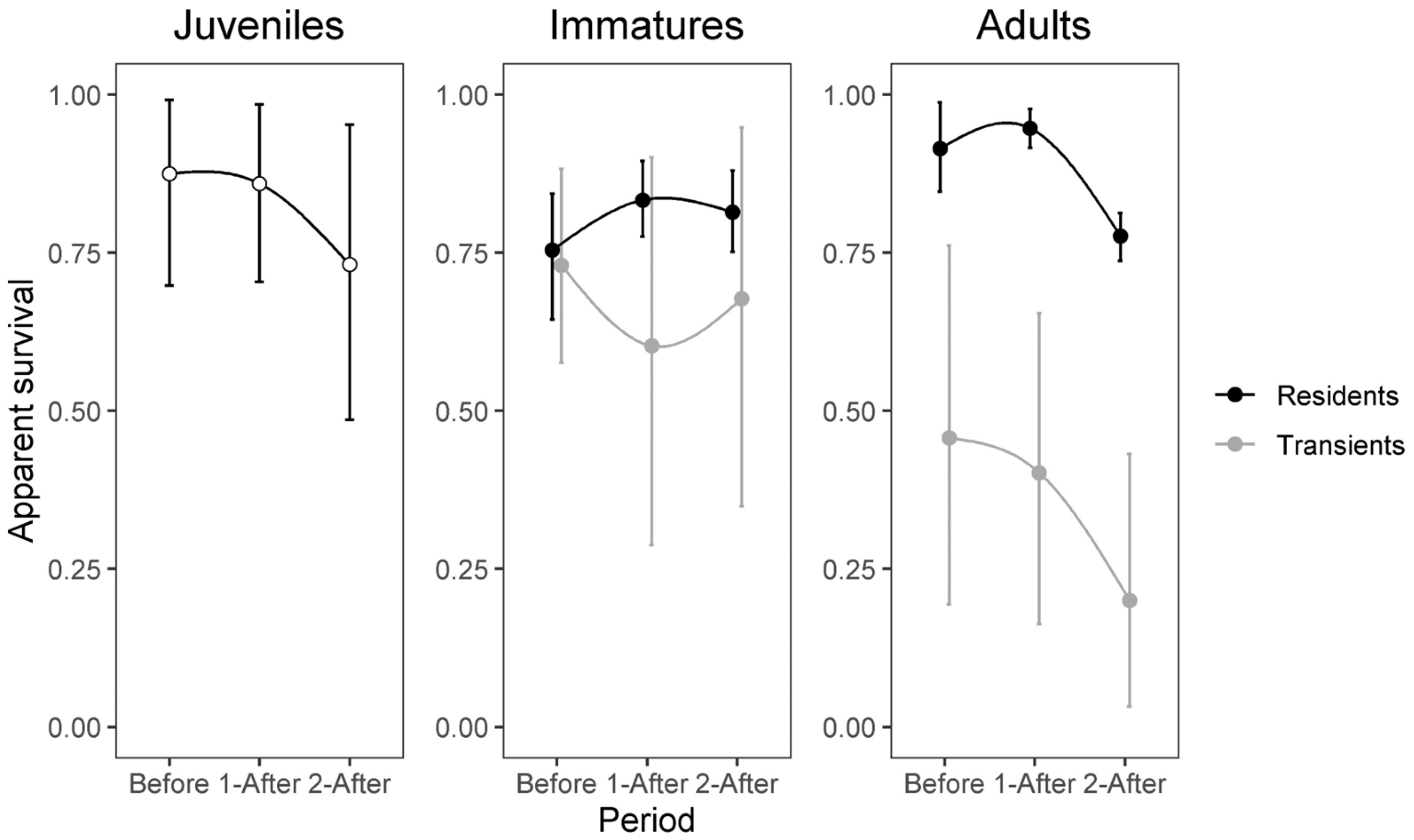
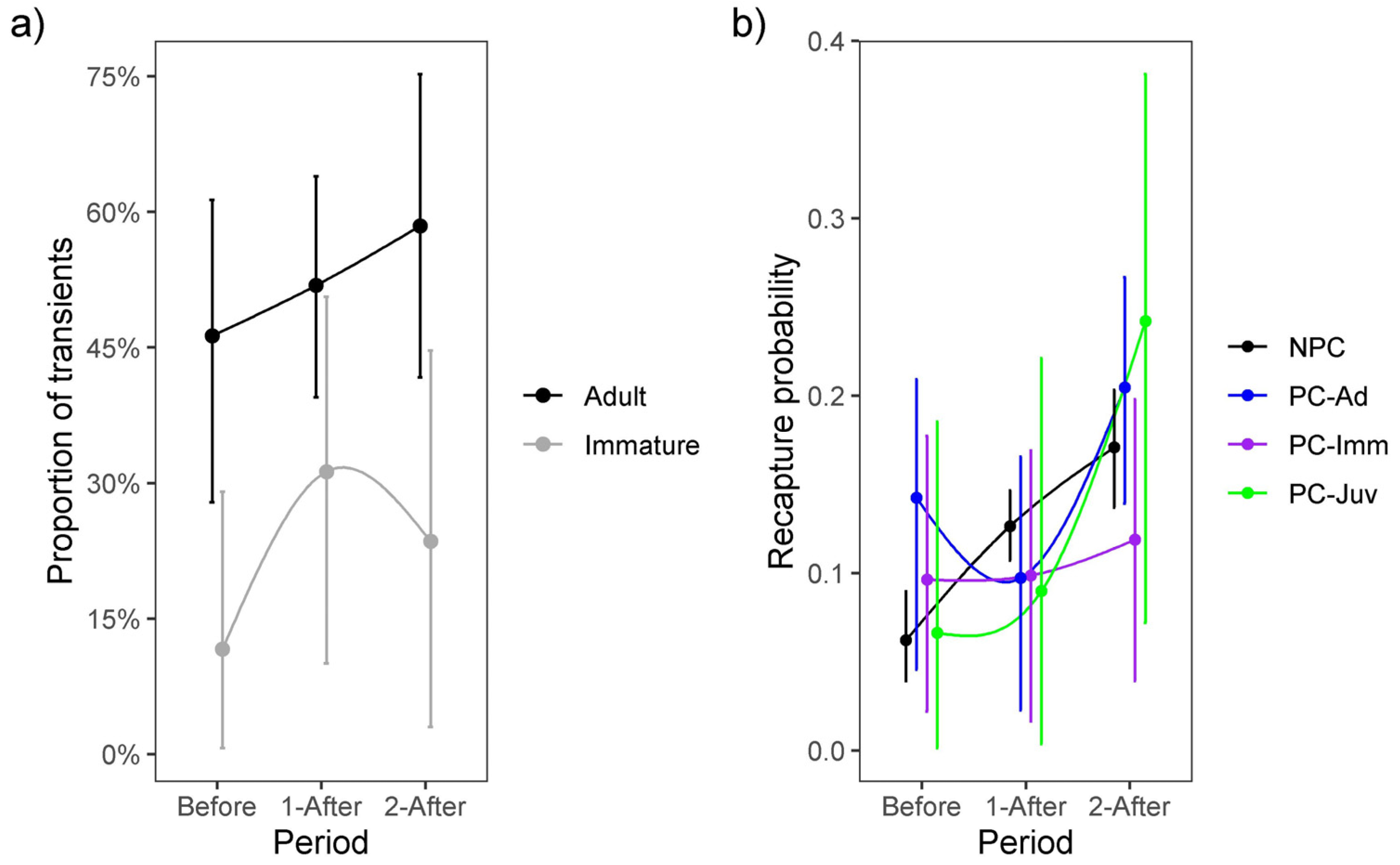
3. Results
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Appendix A

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
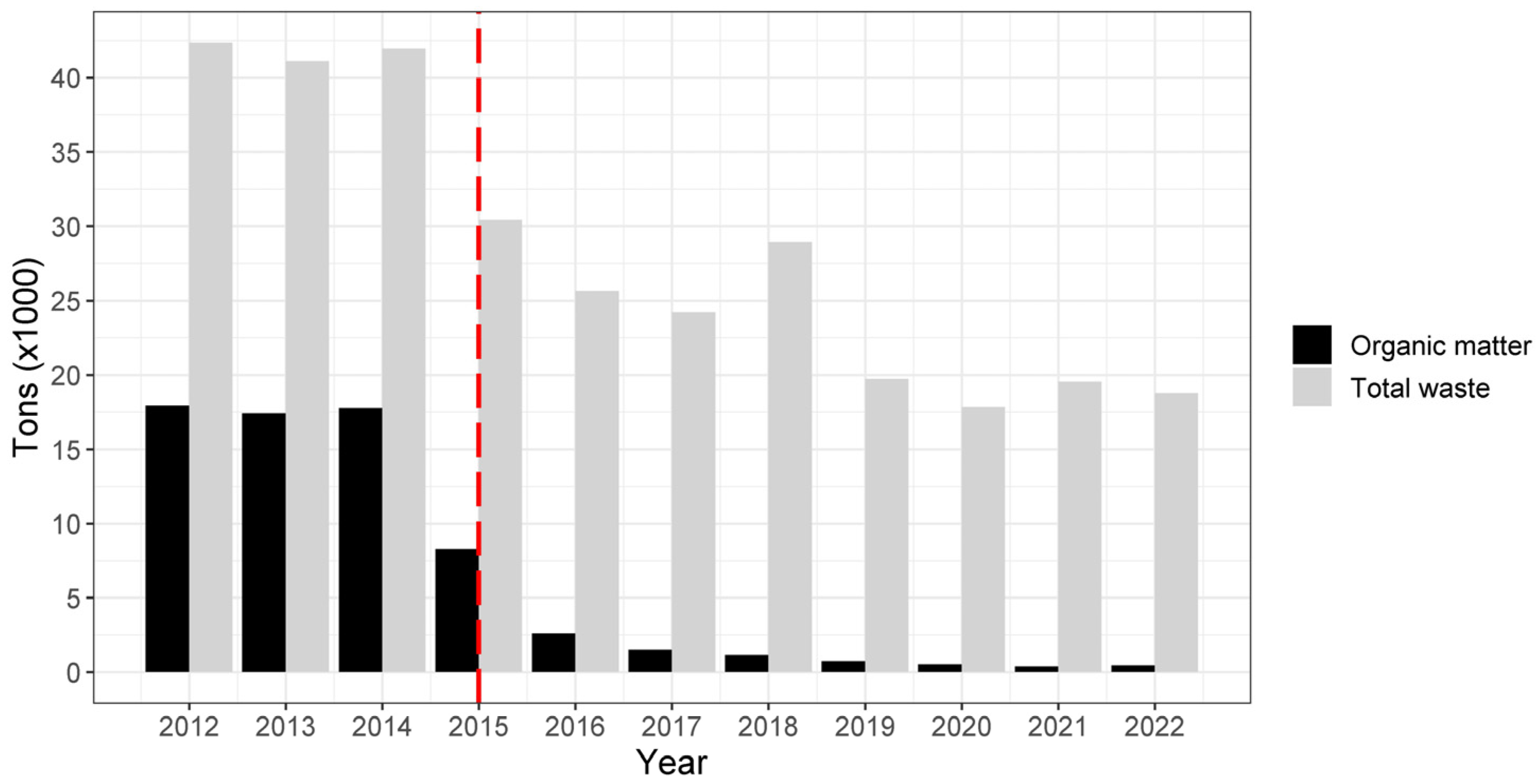
| Periods for CJS Modelling | Year | Organic Waste * | Mean Organic Waste * |
|---|---|---|---|
| Before WTC | 2012 | 17,942.03 | 14,388.68 |
| 2013 | 17,412.38 | ||
| 2014 | 17,775.45 | ||
| 2015 (January–June) | 4424.85 | ||
| 1st After WTC | 2015 (June–December) | 3,861.06 | 2,282.41 |
| 2016 | 2,603.39 | ||
| 2017 | 1,510.11 | ||
| 2018 | 1,155.06 | ||
| 2nd After WTC | 2019 | 723.32 | 514.00 |
| 2020 | 524.55 | ||
| 2021 | 355.12 | ||
| 2022 | 452.99 |
Appendix B
| Periods for CJS Modelling | Years | Marked | Recaptured |
|---|---|---|---|
| Before WTC | 2012 to 2015 (January–June) | 1268 | 239 |
| 1st After WTC | 2015 (June–December) to 2018 | 1174 | 513 |
| 2nd After WTC | 2019 to 2022 | 992 | 779 |
Appendix C
| Period | Parameter | Mean | SD | 95%BCI | |
|---|---|---|---|---|---|
| Lower | Upper | ||||
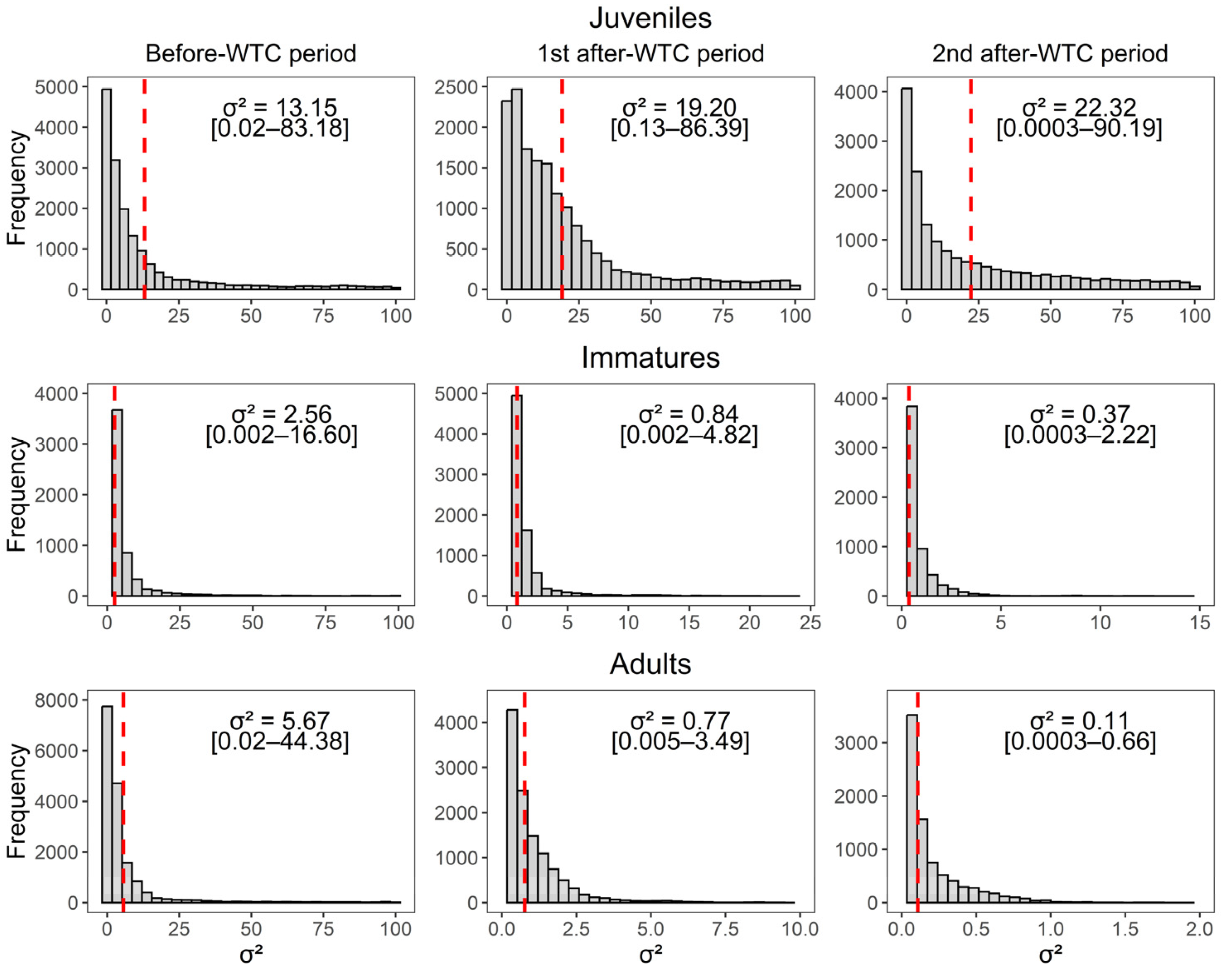
| Before-WTC (2012 to mid-2015) | ϕJuv | 0.8752 | 0.0808 | 0.6983 | 0.9918 |
| ϕRes-Imm | 0.8139 | 0.1100 | 0.5922 | 0.9796 | |
| ϕRes-Adult | 0.9145 | 0.0360 | 0.8468 | 0.9881 | |
| ϕTran-Imm | 0.7539 | 0.0503 | 0.6443 | 0.8434 | |
| ϕTran-Adult | 0.4569 | 0.1418 | 0.1944 | 0.7616 | |
| %TranImm | 0.1176 | 0.0856 | 0.0058 | 0.3150 | |
| %TranAdult | 0.5004 | 0.1586 | 0.1662 | 0.7900 | |
| PNPC | 0.0622 | 0.0126 | 0.0392 | 0.0896 | |
| PPC-Juv | 0.0663 | 0.0517 | 0.0013 | 0.1853 | |
| PPC-Imm | 0.0963 | 0.0423 | 0.0220 | 0.1772 | |
| PPC-Adult | 0.1424 | 0.0416 | 0.0455 | 0.2091 | |
| 1st after-WTC (mid-2015 to 2018) | ϕJuv | 0.8594 | 0.0778 | 0.7037 | 0.9848 |
| ϕRes-Imm | 0.8332 | 0.0301 | 0.7758 | 0.8950 | |
| ϕRes-Adult | 0.9468 | 0.0158 | 0.9157 | 0.9775 | |
| ϕTran-Imm | 0.6026 | 0.1571 | 0.2878 | 0.9010 | |
| ϕTran-Adult | 0.4019 | 0.1233 | 0.1628 | 0.6544 | |
| %TranImm | 0.3176 | 0.1688 | 0.0279 | 0.6721 | |
| %TranAdult | 0.5751 | 0.1313 | 0.3131 | 0.8283 | |
| PNPC | 0.1264 | 0.0100 | 0.1071 | 0.1466 | |
| PPC-Juv | 0.0898 | 0.0625 | 0.0036 | 0.2211 | |
| PPC-Imm | 0.0986 | 0.0394 | 0.0164 | 0.1690 | |
| PPC-Adult | 0.0971 | 0.0384 | 0.0226 | 0.1655 | |
| 2nd after-WTC (2018 to 2022) | ϕJuv | 0.7311 | 0.1205 | 0.4857 | 0.9526 |
| ϕRes-Imm | 0.8143 | 0.0326 | 0.7513 | 0.8799 | |
| ϕRes-Adult | 0.7764 | 0.0192 | 0.7370 | 0.8131 | |
| ϕTran-Imm | 0.6771 | 0.1573 | 0.3490 | 0.9478 | |
| ϕTran-Adult | 0.2003 | 0.1132 | 0.0320 | 0.4318 | |
| %TranImm | 0.2423 | 0.1588 | 0.0126 | 0.6038 | |
| %TranAdult | 0.7416 | 0.1463 | 0.4477 | 0.9587 | |
| PNPC | 0.1708 | 0.0172 | 0.1370 | 0.2032 | |
| PPC-Juv | 0.2420 | 0.0774 | 0.0719 | 0.3811 | |
| PPC-Imm | 0.1187 | 0.0411 | 0.0393 | 0.1980 | |
| PPC-Adult | 0.2046 | 0.0319 | 0.1391 | 0.2667 | |
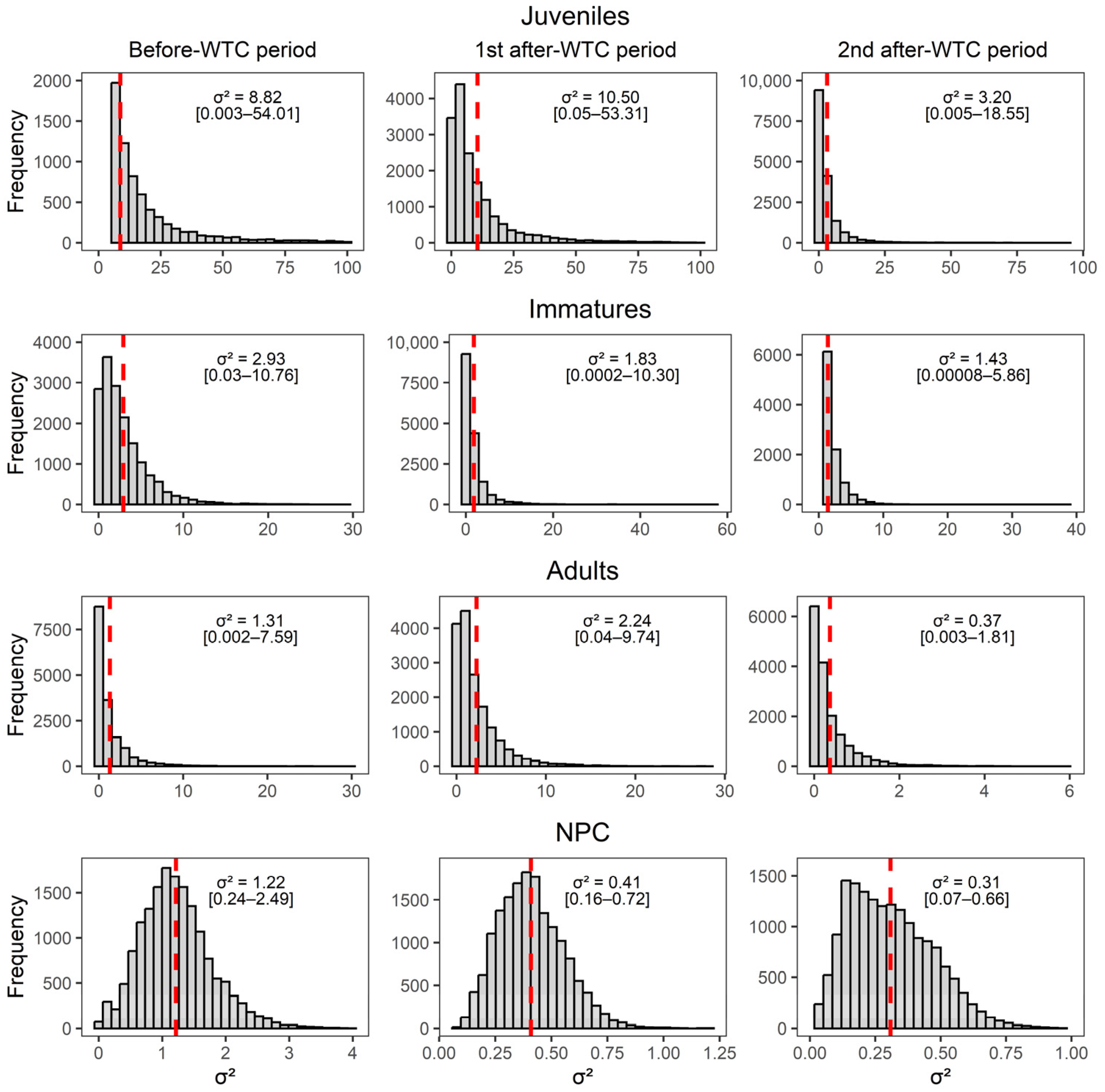
Appendix D
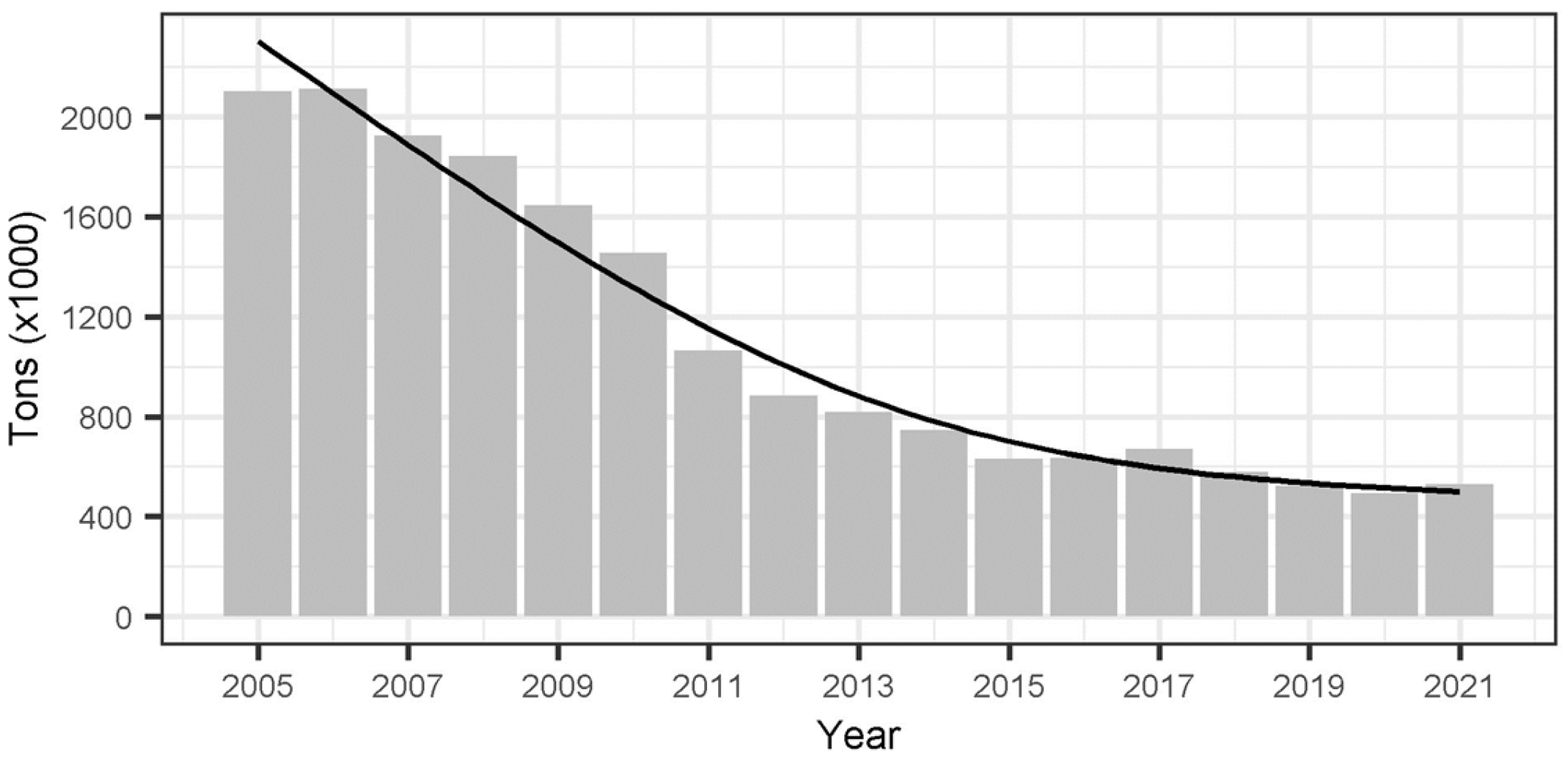
References
- Oro, D.; Cam, E.; Pradel, R.; Martínez-Abraín, A. Influence of food availability on demography and local population dynamics in a long-lived seabird. Proc. R. Soc. Lond. B 2004, 271, 387–396. [Google Scholar]
- McNamara, J.M.; Houston, A.I. Starvation and predation as factors limiting population size. Ecology 1987, 68, 1515–1519. [Google Scholar] [CrossRef]
- Krebs, C.J.; Boutin, S.; Boonstra, R.; Sinclair, A.R.E.; Smith, J.N.M.; Dale, M.R.T.; Martin, K.; Turkington, R. Impact of food and predation on the snowshoe hare cycle. Science 1995, 269, 1112–1115. [Google Scholar]
- Tavecchia, G.; Pradel, R.; Genovart, M.; Oro, D. Density-dependent parameters and demographic equilibrium in open populations. Oikos 2007, 116, 1481–1492. [Google Scholar]
- Briga, M.; Koetsier, E.; Boonekamp, J.J.; Jimeno, B.; Verhulst, S. Food availability affects adult survival trajectories depending on early developmental conditions. Proc. R. Soc. B 2017, 284, 20162287. [Google Scholar] [PubMed]
- Forslund, P.; Pärt, T. Age and reproduction in birds—hypotheses and tests. Trends Ecol. Evol. 1995, 10, 374–378. [Google Scholar]
- Jones, O.R.; Gaillard, J.-M.; Tuljapurkar, S.; Alho, J.S.; Armitage, K.B.; Becker, P.H.; Bize, P.; Brommer, J.; Charmantier, A.; Charpentier, M.; et al. Senescence rates are determined by ranking on the fast-slow life-history continuum. Ecol. Lett. 2008, 11, 664–673. [Google Scholar]
- Kirkwood, T.B.L.; Rose, M.R. Evolution of senescence: Late survival sacrificed for reproduction. Philos. Trans. R. Soc. B 1991, 332, 15–24. [Google Scholar]
- Beauchamp, G. Annual apparent survival across species is lower in juvenile than adult birds but has similar ecological correlates. Ibis 2023, 165, 448–457. [Google Scholar]
- Dewey, S.R.; Kennedy, P.L. Effects of supplemental food on parental-care strategies and juvenile survival of Northern Goshawks. The Auk 2001, 118, 352–365. [Google Scholar]
- Kennedy, P.L.; Ward, J.M. Effects of experimental food supplementation on movements of juvenile northern goshawks (Accipiter gentilis atricapillus). Oecologia 2003, 134, 284–291. [Google Scholar] [CrossRef] [PubMed]
- White, J.D.; Gardali, T.; Thompson, F.R., III; Faaborg, J. Resource selection by juvenile Swainson’s Thrushes during the postfledging period. Condor 2005, 107, 388–401. [Google Scholar] [CrossRef]
- Webb, W.C.; Boarman, W.I.; Rotenberry, J.T. Movements of juvenile common ravens in an arid landscape. J. Wildl. Manag. 2009, 73, 72–81. [Google Scholar] [CrossRef]
- Payo-Payo, A.; Oro, D.; Igual, J.M.; Jover, L.; Sanpera, C.; Tavecchia, G. Population control of an overabundant species achieved through consecutive anthropogenic perturbations. Ecol. Appl. 2015, 25, 2228–2239. [Google Scholar] [CrossRef] [PubMed]
- Oro, D.; Genovart, M.; Tavecchia, G.; Fowler, M.S.; Martínez-Abraín, A. Ecological and evolutionary implications of food subsidies from humans. Ecol. Lett. 2013, 16, 1501–1514. [Google Scholar] [CrossRef] [PubMed]
- Bicknell, A.W.; Oro, D.; Camphuysen, K.; Votier, S.C. Potential consequences of discard reform for seabird communities. J. Appl. Ecol. 2013, 50, 649–658. [Google Scholar] [CrossRef]
- Plaza, P.G.; Lambertucci, S.A. How are garbage dumps impacting vertebrate demography, heath, and conservation? Glob. Ecol. Conserv. 2017, 12, 9–20. [Google Scholar]
- Oro, D.; Margalida, A.; Carrete, M.; Heredia, R.; Donázar, J.A. Testing the goodness of supplementary feeding to enhance population viability in an endangered vulture. PLoS ONE 2008, 3, e4084. [Google Scholar] [CrossRef]
- Oro, D.; Hernández, N.; Jover, L.; Genovart, M. From recruitment to senescence: Food shapes the age-dependent pattern of breeding performance in a long-lived bird. Ecology 2014, 95, 446–457. [Google Scholar]
- Seward, A.M.; Beale, C.M.; Gilbert, L.; Jones, T.H.; Thomas, R.J. The impact of increased food availability on survival of a long-distance migratory bird. Ecology 2013, 94, 221–230. [Google Scholar] [CrossRef]
- Margalida, A.; Donázar, J.A.; Carrete, M.; Sánchez-Zapata, J.A. Sanitary versus environmental policies: Fitting together two pieces of the puzzle of European vulture conservation. J. Appl. Ecol. 2010, 47, 931–935. [Google Scholar] [CrossRef]
- Margalida, A.; Colomer, M.À. Modelling the effects of sanitary policies on European vulture conservation. Sci. Rep. 2012, 2, 753. [Google Scholar] [CrossRef] [PubMed]
- Margalida, A.; Colomer, M.À.; Oro, D. Man-induced activities modify demographic parameters in a long-lived species: Effects of poisoning and health policies. Ecol. Appl. 2014, 24, 436–444. [Google Scholar] [CrossRef] [PubMed]
- Almaraz, P.; Martínez, F.; Morales-Reyes, Z.; Sánchez-Zapata, J.A.; Blanco, G. Long-term demographic dynamics of a keystone scavenger disrupted by human-induced shifts in food availability. Ecol. Appl. 2022, 32, e2579. [Google Scholar] [CrossRef]
- Delgado, S.; Herrero, A.; Galarza, A.; Aldalur, A.; Zorrozua, N.; Arizaga, J. Demographic impact of landfill closure on a resident opportunistic gull. Popul. Ecol. 2021, 63, 238–246. [Google Scholar] [CrossRef]
- Arévalo-Ayala, D.J.; Real, J.; Durà, C.; Aymerich, J.; Hernández-Matías, A. Reduction of organic waste in a landfill lowers the visitation probability but does not the local abundance of a long-lived scavenger species. Bird Conserv. Int. 2023, 33, 1–13. [Google Scholar] [CrossRef]
- Garrido, J.R.; Sarasa, C.G.; Fernández-Cruz, M. Rubbish dumps as key habitats for migration and wintering in the Griffon Vulture (Gyps fulvus) at a migratory bottleneck: Implications for conservation. In Raptors in the New Millennium: Proceedings of the World Conference on Birds of Prey and Owls, Eilat, Israel, 2–8 April 2000; Yosef, R., Miller, M., Pepler, D., Eds.; International Birdwatching Centre Eilat: Eilat, Israel, 2002. [Google Scholar]
- Al Fazari, W.; McGrady, M. Counts of Egyptian vultures Neophron percnopterus and other avian scavengers at Muscat’s municipal landfill, Oman, November 2013–March 2015. Sandgrouse 2016, 38, 99–105. [Google Scholar]
- Rotics, S.; Turjeman, S.; Kaatz, M.; Resheff, Y.S.; Zurell, D.; Sapir, N.; Eggers, U.; Fiedler, W.; Flack, A.; Jeltsch, F.; et al. Wintering in Europe instead of Africa enhances juvenile survival in a long-distance migrant. Anim. Behav. 2017, 126, 79–88. [Google Scholar] [CrossRef]
- Fernández-Gómez, L.; Cortés-Avizanda, A.; Tiago, P.; Byrne, F.; Donázar, J.A. Food subsidies shape age structure in a top avian scavenger. Avian Conserv. Ecol. 2022, 17, 23. [Google Scholar] [CrossRef]
- Ho, C.; Marzluff, J.M.; Stahler, D.R.; Smith, D.W.; Mueller, T.; Wikelski, M.; Safi, K.; Loretto, M.-C. Scavengers use natural and anthropogenic resources connecting protected areas with surrounding lands. Front. Bird Sci. 2023, 2, 1119507. [Google Scholar] [CrossRef]
- Matejczyk, M.; Płaza, G.A.; Nałcz-Jawecki, G.; Ulfig, K.; Markowska-Szczupak, A. Estimation of the environmental risk posed by landfills using chemical, microbiological and ecotoxicological testing of leachates. Chemosphere 2011, 82, 1017–1023. [Google Scholar] [CrossRef]
- De la Casa-Resino, I.; Hernández-Moreno, D.; Castellano, A.; Pérez-López, M.; Soler, F. Breeding near a landfill may influence blood metals (Cd, Pb, Hg, Fe, Zn) and metalloids (Se, As) in white stork (Ciconia ciconia) nestlings. Ecotoxicology 2014, 23, 1377–1386. [Google Scholar] [CrossRef] [PubMed]
- Tauler-Ametller, H.; Pretus, J.L.; Hernández-Matías, A.; Ortiz-Santaliestra, M.E.; Mateo, R.; Real, J. Domestic waste disposal sites secure food availability but diminish plasma antioxidants in Egyptian vulture. Sci. Total Environ. 2019, 650, 1382–1391. [Google Scholar] [CrossRef]
- Ortiz-Santaliestra, M.E.; Tauler-Ametller, H.; Lacorte, S.; Hernández-Matías, A.; Real, J.; Mateo, R. Accumulation of pollutants in nestlings of an endangered avian scavenger related to territory urbanization and physiological biomarkers. Environ. Pollut. 2019, 252, 1801–1809. [Google Scholar] [CrossRef] [PubMed]
- López-García, A.; Sanz-Aguilar, A.; Aguirre, J.I. The trade-offs of foraging at landfills: Landfill use enhances hatching success but decrease the juvenile survival of their offspring on white storks (Ciconia ciconia). Sci. Total Environ. 2021, 778, 146217. [Google Scholar] [CrossRef] [PubMed]
- Gherbi-Salmi, R.; Bachir, A.S.; Ghazi, C.; Doumandji, S.E. How food supply in rubbish dumps affects the breeding success and offspring mortality of cattle Egret Bubulcus ibis? Avian Biol. Res. 2022, 15, 47–52. [Google Scholar] [CrossRef]
- Arrondo, E.; Sebastián-González, E.; Moleón, M.; Morales-Reyes, Z.; Gil-Sánchez, M.J.; Cortés-Avizanda, A.; Ceballos, O.; Donázar, J.A.; Sánchez-Zapata, J.A. Vulture culture: Dietary specialization of an obligate scavenger. Proc. R. Soc. B 2023, 290, 20221951. [Google Scholar] [CrossRef]
- Tauler-Ametller, H.; Hernández-Matías, A.; Pretus, J.L.; Real, J. Landfills determine the distribution of an expanding breeding population of the endangered Egyptian Vulture Neophron percnopterus. Ibis 2017, 159, 757–768. [Google Scholar] [CrossRef]
- Cerecedo-Iglesias, C.; Bartumeus, F.; Cortés-Avizanda, A.; Pretus, J.L.; Hernández-Matías, A.; Real, J. Resource predictability modulates spatial-use networks in an endangered scavenger species. Mov. Ecol. 2023, 11, 22. [Google Scholar] [CrossRef]
- Ogada, D.L.; Keesing, F.; Virani, M.Z. Dropping dead: Causes and consequences of vulture population declines worldwide. Ann. N. Y. Acad. Sci. 2012, 1249, 57–71. [Google Scholar] [CrossRef]
- Margalida, A.; Ogada, D. Old World vultures in a changing environment. In Birds of Prey: Biology and Conservation in the XXI Century; Sarasola, J.H., Grande, J.M., Negro, J.J., Eds.; Springer: New York, NY, USA, 2018; pp. 457–471. [Google Scholar]
- Safford, R.; Andevski, J.; Botha, A.; Bowden, C.; Crockford, N.; Garbett, R.; Margalida, A.; Ramírez, I.; Shobrak, M.; Tavares, J. Vulture conservation: The case for urgent action. Bird Conserv. Int. 2019, 29, 1–9. [Google Scholar] [CrossRef]
- Donázar, J.A.; Cortés-Avizanda, A.; Carrete, M. Dietary shifts in two vultures after the demise of supplementary feeding stations: Consequences of the EU sanitary legislation. Eur. J. Wildl. Res. 2010, 56, 613–621. [Google Scholar] [CrossRef]
- Martínez-Abraín, A.; Tavecchia, G.; Regan, H.M.; Jiménez, J.; Surroca, M.; Oro, D. Effects of wind farms and food scarcity on a large scavenging bird species following an epidemic of bovine spongiform encephalopathy. J. Appl. Ecol. 2012, 49, 109–117. [Google Scholar] [CrossRef]
- Chantepie, S.; Teplitsky, C.; Pavard, S.; Sarrazin, F.; Descaves, B.; Lecuyer, P.; Robert, A. Age-related variation and temporal patterns in the survival of a long-lived scavenger. Oikos 2016, 125, 167–178. [Google Scholar] [CrossRef]
- Pradel, R.; Hines, J.E.; Lebreton, J.-D.; Nichols, J.D. Capture-recapture survival models taking account of transients. Biometrics 1997, 53, 60–72. [Google Scholar] [CrossRef]
- Genovart, M.; Pradel, R. Transience effect in capture-recapture studies: The importance of its biological meaning. PLoS ONE 2019, 14, e0222241. [Google Scholar] [CrossRef]
- Oro, D.; Doak, D.F. Breeding transients in capture–recapture modeling and their consequences for local population dynamics. Sci. Rep. 2020, 10, 15815. [Google Scholar] [CrossRef]
- Dermody, B.J.; Tanner, C.J.; Jackson, A.L. The evolutionary pathway to obligate scavenging in Gyps vultures. PLoS ONE 2011, 6, e24635. [Google Scholar] [CrossRef]
- Zuberogoitia, I.; González-Oreja, J.A.; Martínez, J.R.; Zabala, J.; Gómez, I.; López-López, P. Foraging movements of Eurasian griffon vultures (Gyps fulvus): Implications for supplementary feeding management. Eur. J. Wildl. Res. 2013, 59, 421–429. [Google Scholar] [CrossRef]
- Bosè, M.; Duriez, O.; Sarrazin, F. Intra-specific competition in foraging Griffon Vultures Gyps fulvus: 1. Dynamics of group feeding. Bird Study 2012, 59, 182–192. [Google Scholar] [CrossRef]
- Cortés-Avizanda, A.; Jovani, R.; Carrete, M.; Donázar, J.A. Resource unpredictability promotes species diversity and coexistence in an avian scavenger guild: A field experiment. Ecology 2012, 93, 2570–2579. [Google Scholar] [CrossRef] [PubMed]
- Duriez, O.; Herman, S.; Sarrazin, F. Intra-specific competition in foraging Griffon Vultures Gyps fulvus: 2. The influence of supplementary feeding management. Bird Study 2012, 59, 193–206. [Google Scholar] [CrossRef]
- Duriez, O.; Eliotout, B.; Sarrazin, F. Age identification of Eurasian Griffon Vultures Gyps fulvus in the field. Ring. Migr. 2011, 26, 24–30. [Google Scholar] [CrossRef]
- Zuberogoitia, I.; De la Puente, J.; Elorriaga, J.; Alonso, R.; Palomares, L.E.; Martínez, J.E. The flight feather molt of Griffon vultures (Gyps fulvus) and associated biological consequences. J. Raptor Res. 2013, 47, 292–303. [Google Scholar] [CrossRef]
- Xirouchakis, S.M.; Poulakakis, N. Biometrics, sexual dimorphism and gender determination of Griffon Vultures Gyps fulvus from Crete. Ardea 2008, 96, 91–98. [Google Scholar] [CrossRef]
- Blanco, G.; Martinez, F.; Travekso, J.M. Pair bond and age distribution of breeding Griffon Vultures Gyps fulvus in relation to reproductive status and geographic area in Spain. Ibis 1997, 139, 180–183. [Google Scholar] [CrossRef]
- Hargrove, J.W.; Borland, C.H. Pooled population parameter estimates from mark-recapture data. Biometrics 1994, 50, 1129–1141. [Google Scholar] [CrossRef]
- O’Brien, S.; Robert, B.; Tiandry, H. Consequences of violating the recapture duration assumption of mark–recapture models: A test using simulated and empirical data from an endangered tortoise population. J. Appl. Ecol. 2005, 42, 1096–1104. [Google Scholar] [CrossRef]
- Dupont, P.; Milleret, C.; Gimenez, O.; Bischof, R. Population closure and the bias-precision trade-off in spatial capture–recapture. Methods Ecol. Evol. 2019, 10, 661–672. [Google Scholar] [CrossRef]
- Fernández, J.A. Comportamiento del buitre leonado (Gyps f. fulvus) en nido. Ardeola 1997, 22, 29–54. [Google Scholar]
- Cramp, S.; Simmons, K.E.L. The Birds of the Western Palearctic, 1st ed.; Oxford University Press: Oxford, UK, 1980. [Google Scholar]
- Xirouchakis, S.M. Breeding biology and reproductive performance of Griffon Vultures Gyps fulvus on the island of Crete (Greece). Bird Study 2010, 57, 213–225. [Google Scholar] [CrossRef]
- Choquet, R.; Lebreton, J.D.; Gimenez, O.; Reboulet, A.M.; Pradel, R. U-CARE: Utilities for performing goodness of fit tests and manipulating CApture-REcapture data. Ecography 2009, 32, 1071–1074. [Google Scholar] [CrossRef]
- Gimenez, O.; Rossi, V.; Choquet, R.; Dehais, C.; Doris, B.; Varella, H.; Vila, J.P.; Pradel, R. State-space modelling of data on marked individuals. Ecol. Model. 2007, 206, 431–438. [Google Scholar] [CrossRef]
- Royle, J.A. Modeling individual effects in the Cormack-Jolly-Seber model: A state-space formulation. Biometrics 2008, 64, 364–370. [Google Scholar] [CrossRef] [PubMed]
- Kéry, M.; Schaub, M. Bayesian Population Analysis Using WinBUGS: A Hierarchical Perspective; Academic Press: Waltham, MA, USA, 2012. [Google Scholar]
- Ver Hoef, J.M.; Boveng, P.L. Iterating on a single model is a viable alternative to multimodel inference. J. Wildl. Manag. 2015, 79, 719–729. [Google Scholar] [CrossRef]
- Cam, E.; Link, W.A.; Cooch, E.G.; Monnat, J.Y.; Danchin, E. Individual covariation in life-history traits: Seeing the trees despite the forest. Am. Nat. 2002, 159, 96–105. [Google Scholar] [CrossRef] [PubMed]
- Kéry, M. Introduction to WinBUGS for Ecologists. A Bayesian Approach to Regression, ANOVA, Mixed Models and Related Analyses; Academic Press: Burlington, VT, USA, 2010. [Google Scholar]
- Service, P.M. Heterogeneity in individual mortality risk and its importance for evolutionary studies of senescence. Am. Nat. 2000, 156, 1–13. [Google Scholar] [CrossRef]
- Gimenez, O.; Cam, E.; Gaillard, J.M. Individual heterogeneity and capture–recapture models: What, why and how? Oikos 2018, 127, 664–686. [Google Scholar] [CrossRef]
- Abadi, F.; Botha, A.; Altwegg, R. Revisiting the effect of capture heterogeneity on survival estimates in capture-mark-recapture studies: Does it matter? PLoS ONE 2013, 8, e62636. [Google Scholar] [CrossRef]
- Plummer, M. JAGS: A program for analysis of Bayesian graphical models using Gibbs sampling. In Proceedings of the 3rd International Workshop on Distributed Statistical Computing (DSC 2003), Vienna, Austria, 20–22 March 2003. [Google Scholar]
- Kellner, K.; Meredith, M. jagsUI: A Wrapper around Rjags to Streamline JAGS Analyses. 2021. R Package Version 1.5.2. Available online: https://github.com/kenkellner/jagsUI (accessed on 20 May 2023).
- R Core Team. R. A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2021; Available online: https://www.r-project.org/ (accessed on 15 April 2023).
- Gelman, A.; Hill, J. Data Analysis Using Regression and Multilevel/Hierarchical Models, 1st ed.; Cambridge University Press: Cambridge, UK, 2006. [Google Scholar]
- Pradel, R. Flexibility in survival analysis from recapture data: Handling trap-dependence. In Marked Individuals in the Study of Bird Population; Lebreton, J.D., North, P.M., Eds.; Birkhäuser Verlag: Basel, Switzerland, 1993; pp. 29–37. [Google Scholar]
- Lebreton, J.D.; Burnham, K.P.; Clobert, J.; Anderson, D.R. Modeling survival and testing biological hypothesis using marked animals: A unified approach with case studies. Ecol. Monogr. 1992, 62, 67–118. [Google Scholar] [CrossRef]
- Xirouchakis, S.M.; Grivas, C.; Andreou, G.; Georgopoulou, E. Home range size, space use and resource selection of griffon vultures in an insular environment. J. Zool. 2021, 314, 116–131. [Google Scholar] [CrossRef]
- García-Ripollés, C.; López-López, P.; Urios, V. Ranging behaviour of non-breeding Eurasian Griffon Vultures Gyps fulvus: A GPS-telemetry study. Acta Ornithol. 2011, 46, 127–134. [Google Scholar] [CrossRef]
- Monsarrat, S.; Benhamo, S.; Sarrazin, F.; Bessa-Gomes, C.; Bouten, W.; Duriez, O. How predictability of feeding patches affects home range and foraging habitat selection in avian social scavengers? PLoS ONE 2013, 8, e53077. [Google Scholar] [CrossRef] [PubMed]
- Harel, R.; Duriez, O.; Spiegel, O.; Fluhr, N.; Horvitz, N.; Getz, W.M.; Bouten, W.; Sarrazin, F.; Hatzofe, O.; Nathan, R. Decision-making by a soaring bird: Time, energy and risk considerations at different spatio-temporal scales. Phil. Trans. R. Soc. B 2016, 371, 20150397. [Google Scholar] [CrossRef] [PubMed]
- Morant, J.; Arrondo, E.; Sánchez-Zapata, J.A.; Donázar, J.A.; Cortés-Avizanda, A.; De La Riva, M.; Blanco, G.; Martínez, F.; Oltra, J.; Carrete, M.; et al. Large-scale movement patterns in a social vulture are influenced by seasonality, sex, and breeding region. Ecol. Evol. 2023, 13, e9817. [Google Scholar] [CrossRef]
- Moreno-Opo, R.; Trujillano, A.; Margalida, A. Larger size and older age confer competitive advantage: Dominance hierarchy within European vulture guild. Sci. Rep. 2020, 10, 2430. [Google Scholar] [CrossRef]
- Carneiro, M.; Colaço, B.; Brandão, R.; Azorín, B.; Nicolas, O.; Colaço, J.; Pires, M.J.; Agustí, S.; Casas-Díaz, E.; Lavin, S. Assessment of the exposure to heavy metals in Griffon vultures (Gyps fulvus) from the Iberian Peninsula. Ecotoxicol. Environ. Saf. 2015, 113, 295–301. [Google Scholar] [CrossRef]
- Margalida, A.; Sánchez-Zapata, J.A.; Blanco, G.; Hiraldo, F.; Donázar, J.A. Diclofenac approval as a threat to Spanish vultures. Conserv. Biol. 2014, 28, 631–632. [Google Scholar] [CrossRef]
- Green, R.E.; Donazar, J.A.; Sánchez-Zapata, J.A.; Margalida, A. Potential threat to Eurasian griffon vultures in Spain from veterinary use of the drug diclofenac. J. Appl. Ecol. 2016, 53, 993–1003. [Google Scholar] [CrossRef]
- Herrero-Villar, M.; Sánchez-Barbudo, I.S.; Camarero, P.R.; Taggart, M.A.; Mateo, R. Increasing incidence of barbiturate intoxication in avian scavengers and mammals in Spain. Environ. Pollut. 2021, 284, 117452. [Google Scholar] [CrossRef]
- Oliva-Vidal, P.; Martínez, J.M.; Sánchez-Barbudo, I.S.; Camarero, P.R.; Colomer, M.À.; Margalida, A.; Mateo, R. Second-generation anticoagulant rodenticides in the blood of obligate and facultative European avian scavengers. Environ. Pollut. 2022, 315, 120385. [Google Scholar] [CrossRef] [PubMed]
- Molina-López, R.A.; Casal, J.; Darwich, L. Causes of morbidity in wild raptor populations admitted at a wildlife rehabilitation centre in Spain from 1995-2007: A long-term retrospective study. PLoS ONE 2011, 6, e24603. [Google Scholar] [CrossRef] [PubMed]
- Carrete, M.; Sánchez-Zapata, J.A.; Benítez, J.R.; Lobón, M.; Montoya, F.; Donázar, J.A. Mortality at wind-farms is positively related to large-scale distribution and aggregation in griffon vultures. Biol. Conserv. 2012, 145, 102–108. [Google Scholar] [CrossRef]
- Espín, S.; Martínez-López, E.; Jiménez, P.; María-Mojica, P.; García-Fernández, A.J. Effects of heavy metals on biomarkers for oxidative stress in Griffon vulture (Gyps fulvus). Environ. Res. 2014, 129, 59–68. [Google Scholar] [CrossRef]
- Badia-Boher, J.A.; Sanz-Aguilar, A.; de la Riva, M.; Gangoso, L.; van Overveld, T.; García-Alfonso, M.; Luzardo, O.P.; Suarez-Pérez, A.; Donázar, J.A. Evaluating European LIFE conservation projects: Improvements in survival of an endangered vulture. J. Appl. Ecol. 2019, 56, 1210–1219. [Google Scholar] [CrossRef]
- Santangeli, A.; Pakanen, V.M.; Bridgeford, P.; Boorman, M.; Kolberg, H.; Sanz-Aguilar, A. The relative contribution of camera trap technology and citizen science for estimating survival of an endangered African vulture. Biol. Conser. 2020, 246, 108593. [Google Scholar] [CrossRef]
- Pradel, R.; Sanz-Aguilar, A. Modeling trap-awareness and related phenomena in capture-recapture studies. PLoS ONE 2012, 7, e32666. [Google Scholar] [CrossRef]
- Crespin, L.; Choquet, R.; Lima, M.; Merritt, J.; Pradel, R. Is heterogeneity of catchability in capture–recapture studies a mere sampling artifact or a biologically relevant feature of the population? Popul. Ecol. 2008, 50, 247–256. [Google Scholar] [CrossRef]
- Oliva-Vidal, P.; Hernández-Matías, A.; García, D.; Colomer, M.À.; Real, J.; Margalida, A. Griffon vultures, livestock and farmers: Unraveling a complex socio-economic ecological conflict from a conservation perspective. Biol. Conserv. 2022, 272, 109664. [Google Scholar] [CrossRef]
- Arkumarev, V.; Dobrev, D.; Stamenov, A.; Terziev, N.; Delchev, A.; Stoychev, S. Seasonal dynamics in the exploitation of natural carcasses and supplementary feeding stations by a top avian scavenger. J. Ornithol. 2021, 162, 723–735. [Google Scholar] [CrossRef]
- Moreno-Opo, R.; Trujillano, A.; Margalida, A. Optimization of supplementary feeding programs for European vultures depends on environmental and management factors. Ecosphere 2015, 6, 127. [Google Scholar] [CrossRef]
- Morales-Reyes, Z.; Pérez-García, J.M.; Moleón, M.; Botella, F.; Carrete, M.; Donázar, J.A.; Cortés-Avizanda, A.; Arrondo, E.; Moreno-Opo, R.; Jiménez, J.; et al. Evaluation of the network of protection areas for the feeding of scavengers (PAFs) in Spain: From biodiversity conservation to greenhouse gas emission savings. J. Appl. Ecol. 2017, 54, 1120–1129. [Google Scholar] [CrossRef]
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Arévalo-Ayala, D.J.; Real, J.; Mañosa, S.; Aymerich, J.; Durà, C.; Hernández-Matías, A. Age-Specific Demographic Response of a Long-Lived Scavenger Species to Reduction of Organic Matter in a Landfill. Animals 2023, 13, 3529. https://doi.org/10.3390/ani13223529
Arévalo-Ayala DJ, Real J, Mañosa S, Aymerich J, Durà C, Hernández-Matías A. Age-Specific Demographic Response of a Long-Lived Scavenger Species to Reduction of Organic Matter in a Landfill. Animals. 2023; 13(22):3529. https://doi.org/10.3390/ani13223529
Chicago/Turabian StyleArévalo-Ayala, Diego J., Joan Real, Santi Mañosa, Joan Aymerich, Carles Durà, and Antonio Hernández-Matías. 2023. "Age-Specific Demographic Response of a Long-Lived Scavenger Species to Reduction of Organic Matter in a Landfill" Animals 13, no. 22: 3529. https://doi.org/10.3390/ani13223529
APA StyleArévalo-Ayala, D. J., Real, J., Mañosa, S., Aymerich, J., Durà, C., & Hernández-Matías, A. (2023). Age-Specific Demographic Response of a Long-Lived Scavenger Species to Reduction of Organic Matter in a Landfill. Animals, 13(22), 3529. https://doi.org/10.3390/ani13223529