
Diet Mixing and Supplementation Present an Opportunity to Increase the Use of Encroaching Woody Plants by Goats
 , , ,
, , ,
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Study Area
2.2. Study Plants and Animals
2.3. Experimental Designs
2.4. Experiment One
2.5. Experiment Two
2.6. Chemical Composition
2.7. Data Analyses
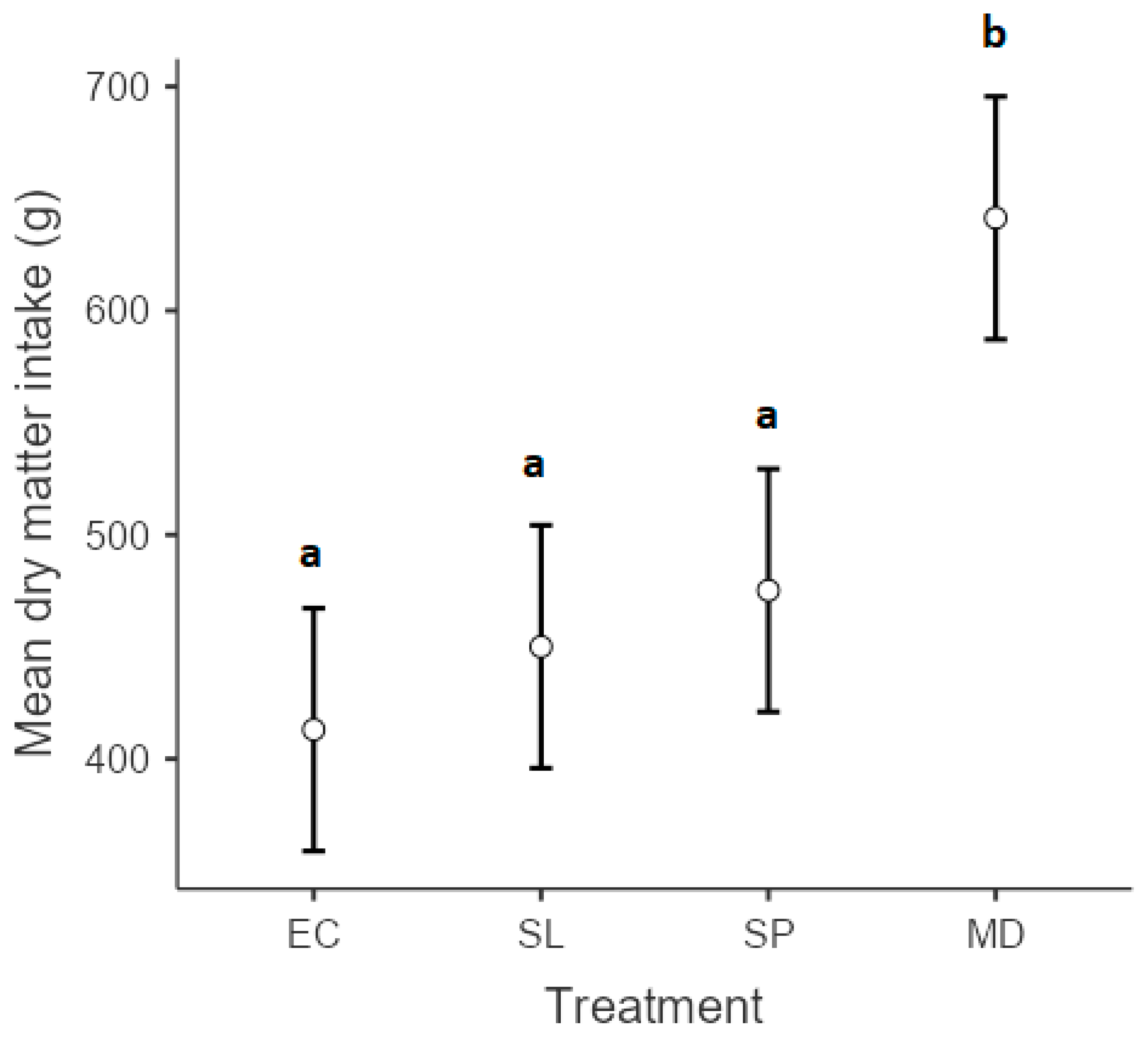
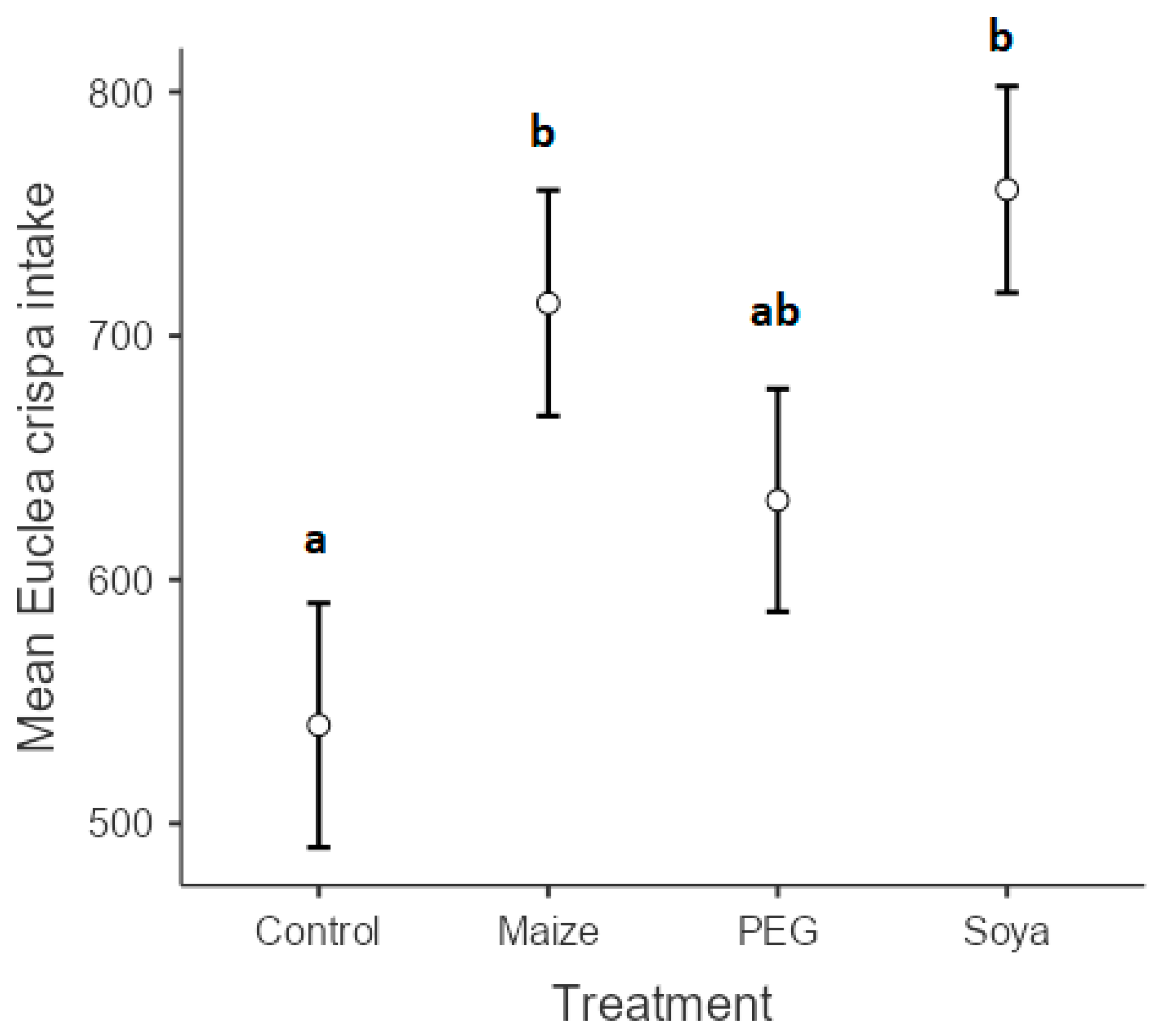
3. Results
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Hughes, L. Climate change and Australia: Trends, projections, and impacts. Aust. Ecol. 2003, 28, 423–443. [Google Scholar] [CrossRef]
- Archer, S.R.; Andersen, E.M.; Predick, K.I.; Schwinning, S.; Steidl, R.J.; Woods, S.R. Woody plant encroachment: Causes and consequences. In Rangeland Systems: Processes, Management, and Challenges; Briske, D.D., Ed.; Springer: New York, NY, USA, 2017; pp. 25–84. [Google Scholar] [CrossRef]
- FAO. The State of Food and Agriculture: Livestock in the Balance; Food and Agriculture Organisation of the United Nations: Rome, Italy, 2009. [Google Scholar]
- Estell, R.E.; Havstad, K.M.; Cibils, A.F.; Fredrickson, E.L.; Anderson, D.M.; Schrader, T.S.; James, D.K. Increasing Shrub Use by Livestock in a World with Less Grass. Rangel. Ecol. Manag. 2012, 65, 553–562. [Google Scholar] [CrossRef]
- Herrero, M.; Thornton, P.K.; Notenbaert, A.M.; Wood, S.; Msangi, S.; Freeman, H.A.; Bossio, D.; Dixon, J.; Peters, M.; van de Steeg, J.; et al. Smart investments in sustainable food production: Revisiting mixed crop-livestock systems. Science 2010, 327, 822–825. [Google Scholar] [CrossRef] [PubMed]
- Estell, R.E.; Havstad, K.M.; Cibils, A.F.; Anderson, D.M.; Schrader, T.S. The changing role of shrubs in rangeland-based livestock production systems: Can shrubs increase our forage supply? Rangelands 2014, 36, 25–31. [Google Scholar] [CrossRef]
- Cohen, J.E. Human population: The next half century. Science 2003, 302, 1172–1175. [Google Scholar] [CrossRef] [PubMed]
- Rogosic, J.; Estell, R.E.; Ivankovic, S.; Kezic, J.; Razov, J. Potential mechanisms to increase shrub intake and performance of small ruminants in Mediterranean shruby ecosystems. Small Rumin. Res. 2008, 74, 1–15. [Google Scholar] [CrossRef]
- Liraa, R.; MacAdam, J.; Sales, F.; Villalba, J.J. Supplemental levels of protein and energy influence ingestion of Romerillo (Chiliotrichum diffusum) by sheep in southern Patagonia. Small Rumin. Res. 2020, 191, 106160. [Google Scholar] [CrossRef]
- Coley, P.D.; Barone, J.A. Herbivory and plant defenses in tropical forests. Annu. Rev. Ecol. Syst. 1996, 27, 305–335. [Google Scholar] [CrossRef]
- Freeland, W.J.; Janzen, D.H. Strategies in herbivory by mammals: The role of plant secondary compounds. Am. Nat. 1974, 108, 269–286. [Google Scholar] [CrossRef]
- Marsh, K.J.; Wallis, I.R.; Mclean, S.; Sorensen, J.S.; Foley, W.J. Conflicting demands on detoxification pathways influence how Brushtail possums choose their diets. Ecology 2006, 87, 2103–2112. [Google Scholar] [CrossRef]
- Villalba, J.J.; Provenza, F.D.; Bryant, J.P. Consequences of the interaction between nutrients and plant secondary metabolites on herbivore selectivity: Benefits or detriments for plants? Oikos 2002, 97, 282–292. [Google Scholar] [CrossRef]
- Banner, R.E.; Rogosic, J.; Burritt, E.A.; Provenza, F.D. Supplemental barley and charcoal increase intake of sagebrush by lambs. J. Range Manag. 2000, 53, 415–420. [Google Scholar] [CrossRef]
- Dziba, L.E.; Provenza, F.D.; Villalba, J.J.; Atwood, S.B. Supplemental energy and protein increase use of sagebrush by sheep. Small Rumin. Res. 2007, 69, 203–207. [Google Scholar] [CrossRef]
- Illius, A.W.; Jessop, N.S. Metabolic constraints on voluntary intake in ruminants. J. Anim. Sci. 1996, 74, 3052–3062. [Google Scholar] [CrossRef] [PubMed]
- Estell, R.E. Coping with shrub secondary metabolites by ruminants. Small Rumin. Res. 2010, 94, 1–9. [Google Scholar] [CrossRef]
- Villalba, J.J.; Provenza, F.D.; Han, G.-D. Experience influences diet mixing by herbivores: Implications for plant biochemical diversity. Oikos 2004, 107, 100–109. [Google Scholar] [CrossRef]
- Papachristou, T.G.; Dziba, L.E.; Villalba, J.J.; Provenza, F.D. Patterns of diet mixing by sheep offered foods varying in nutrients and plant secondary compounds. Appl. Anim. Behav. Sci. 2007, 108, 68–80. [Google Scholar] [CrossRef]
- Rogosic, J.; Estell, R.E.; Skobic, D.; Martinovic, A.; Maric, S. Role of species diversity and secondary compound complementarity on diet selection of Mediterranean shrubs by goats. J. Chem. Ecol. 2006, 32, 1279–1287. [Google Scholar] [CrossRef]
- Rogosic, J.; Estell, R.E.; Skobic, D.; Stanic, S. Influence of secondary compound complementarity and species diversity on the consumption of Mediterranean shrubs by sheep. Appl. Anim. Behav. Sci. 2007, 107, 58–65. [Google Scholar] [CrossRef]
- Šarić, T.; Rogošić, J.; Provenza, F.; Zupan, I.; Tkalčić, S.; Franin, K.; Šikić, Z.; Ivanković, S.; Herceg, N. Mediterranean shrub diversity and its effect on food intake in goats. Ital. J. Anim. Sci. 2014, 13, 582–587. [Google Scholar] [CrossRef]
- Foley, W.J.; Iason, G.R.; McArthur, C. Role of plant secondary metabolites in the nutritional ecology of mammalian herbivores: How far have we come in 25 years? In Nutritional Ecology of Herbivores. Proceedings of the Fifth International Symposium on Nutritional Ecology of Herbivores; Jung, H.G., Fahey, G.C., Jr., Eds.; American Society of Animal Science: Savoy, IL, USA, 1999; pp. 130–209. [Google Scholar]
- Windels, S.K.; Hewitt, D.G.; Forbes, T.D.A. Effects of aeration on phenolic amine content of guajillo. J. Range Manag. 2003, 56, 529–533. [Google Scholar] [CrossRef]
- Bryant, J.P.; Chapin, F.S.; Klein, D.R. Carbon/nutrient balance of boreal plants in relation to vertebrate herbivory. Oikos 1983, 40, 357–368. [Google Scholar] [CrossRef]
- Bryant, J.P.; Reichardt, P.B.; Clausen, T. Chemically mediated interactions between woody plants and browsing mammals. J. Range Manag. 1992, 45, 18–24. [Google Scholar] [CrossRef]
- Scogings, P.F.; Hjältén, J.; Skarpe, C.; Hattas, D.; Zobolo, A.; Dziba, L.; Rooke, T. Nutrient and secondary metabolite concentrations in a savanna are independently affected by large herbivores and shoot growth rate. Plant Ecol. 2014, 215, 73–82. [Google Scholar] [CrossRef]
- Mueller-Harvey, I. Review: Unravelling the conundrum of tannins in animal nutrition and health. J. Sci. Food Agric. 2006, 86, 2010–2037. [Google Scholar] [CrossRef]
- Foley, W.L.; Lason, G.; Makkar, H. Transdisciplinary studies of plant secondary metabolites: Lessons from ecology for animal science and vice versa. In Proceedings of the 7th International Symposium on the Nutrition of Herbivores, Beijing, China, 16–22 September 2007; Meng, Q.X., Liu, J.X., Zhu, W.Y., Eds.; China Agricultural University Press: Beijing, China, 2007. [Google Scholar]
- Marsh, K.J.; Wallis, I.R.; Foley, W.J. The effect of inactivating tannins on the intake of Eucalyptus foliage by a specialist Eucalyptus folivore (Pseudocheirus peregrinus) and a generalist herbivore (Trichosurus vulpecula). Aust. J. Zool. 2003, 51, 31–42. [Google Scholar] [CrossRef]
- Marsh, K.J.; Wallis, I.R.; Foley, W.J. Detoxification rates constrain feeding in common brushtail possums (Trichosurus vulpecula). Ecology 2005, 86, 2946–2954. [Google Scholar] [CrossRef]
- Schmitt, M.H.; Ward, D.; Shrader, A.M. Salivary tannin-binding proteins: A foraging advantage for goats? Livest. Sci. 2020, 234, e103974. [Google Scholar] [CrossRef]
- Ventura-Cordero, J.; Sandoval-Castro, C.A.; Torres-Acosta, J.F.J.; Capetillo-Leal, C.M. Do goats have a salivary constitutive response to tannins? J. Appl. Anim. Res. 2017, 45, 29–34. [Google Scholar] [CrossRef]
- Provenza, F.D.; Villalba, J.J.; Dziba, L.E.; Atwood, S.B.; Banner, R.E. Linking herbivore experience, varied diets, and plant biochemical diversity. Small Rumin. Res. 2003, 49, 257–274. [Google Scholar] [CrossRef]
- Provenza, F.; Papachristou, T. Behavior-based management of ecosystems. Opt. Méditerranéennes Ser. A 2009, 85, 13–28. [Google Scholar]
- Mucina, I.; Rutherford, M.C. The Vegetation of South Africa, Lesotho and Swaziland. Strelitzia 19; South Africa National Biodiversity Institute: Pretoria, South Africa, 2006. [Google Scholar]
- Mkhize, N.R.; Heitkönig, I.M.A.; Scogings, P.F.; Dziba, L.E.; Prins, H.H.T. Seasonal regulation of condensed tannin consumption by free-ranging goats in a semi-arid savanna. PLoS ONE 2018, 13, e0189626. [Google Scholar] [CrossRef] [PubMed]
- Coates-Palgrave, M. Keith Coates-Palgrave trees of southern Africa, 3rd ed.; Struik Publishers: Cape Town, South Africa, 2005. [Google Scholar]
- Kerley, G.I.H.; Knight, M.H.; de Kock, M. Desertification of subtropical thicket in the Eastern Cape, South Africa: Are there alternatives? Environ. Monit. Assess. 1995, 37, 211–230. [Google Scholar] [CrossRef] [PubMed]
- Devendra, C. The digestive efficiency of goats. World Rev. Anim. Prod. 1978, 14, 9–12. [Google Scholar]
- Mdladla, K.; Dzomba, E.F.; Muchadeyi, F.C. Seroprevalence of Ehrlichia ruminantium antibodies and its associated risk factors in indigenous goats of South Africa. Prev. Vet. Med. 2016, 125, 99–105. [Google Scholar] [CrossRef]
- Heuzé, V.; Tran, G.; Kaushik, S. Soybean Meal. Feedipedia.org. Animal Feed Resources Information System. Last Updated 4 March 2020, 16:02. Available online: http://www.feedipedia.org/node/556 (accessed on 1 August 2023).
- Heuzé, V.; Tran, G.; Lebas, F. Maize Grain. Feedipedia.org. A Programme by INRA, CIRAD, AFZ and FAO. Last Updated on 7 September 2017, 14:59. Available online: http://www.feedipedia.org/node/556 (accessed on 1 August 2023).
- Association of Official Analytical Chemists. Official Methods of Analyses, 17th ed.; AOAC International: Arlington, VA, USA, 2000. [Google Scholar]
- Van Soest, P.J.; Robertson, J.B.; Lewis, B.A. Methods of dietary fiber, neutral detergent fiber and non-starch polysaccharides in relation to animal nutrition. J. Dairy Sci. 1991, 74, 3583–3597. [Google Scholar] [CrossRef]
- Goering, H.K.; Van Soest, P.J. Forage Fibre Analyses; Agricultural Handbook No 379; ARS-US Department of Agriculture: Washington, DC, USA, 1970.
- Bailey, D.W.; Provenza, F.D. Mechanisms determining large-herbivore distribution. In Resource Ecology: Spatial and Temporal Dynamics of Foraging; Prins, H.H.T., Van Langevelde, F., Eds.; Springer: Dordrecht, The Netherlands, 2008; pp. 7–28. [Google Scholar] [CrossRef]
- Mkhize, N.R.; Heitkönig, I.M.A.; Scogings, P.F.; Dziba, L.E.; Prins, H.H.T.; de Boer, W.F. Condensed tannins reduce browsing and increase grazing time of free-ranging goats in semi-arid savannas. Appl. Anim. Behav. Sci. 2015, 169, 33–37. [Google Scholar] [CrossRef]
- Chalupa, W. Rumen bypass and protection of proteins and amino Acids. J. Dairy Sci. 1975, 58, 1198–1218. [Google Scholar] [CrossRef]
- Lisonbee, L.D.; Villalba, J.J.; Provenza, F.D. Effects of tannin on selection by sheep of forages containing alkaloids, tannins and saponins. J. Sci. Food Agric. 2009, 89, 2668–2677. [Google Scholar] [CrossRef]
- Mkhize, N.R.; Heitkönig, I.M.A.; Scogings, P.F.; Dziba, L.E.; Prins, H.H.T.; de Boer, W.F. Supplemental nutrients increase the consumption of chemically defended shrubs by free-ranging herbivores. Agric. Ecosyst. Environ. 2016, 235, 119–126. [Google Scholar] [CrossRef]
- Makkar, H.P.S. Effects of tannins in ruminant animals, adaptation to tannins, and strategies to overcome detrimental effects of feeding tannin-rich feeds. Small Rumin. Res. 2003, 49, 241–256. [Google Scholar] [CrossRef]
- Shimada, I. Salivary proteins as a defence against dietary tannins. J. Chem. Ecol. 2006, 32, 1149–1163. [Google Scholar] [CrossRef] [PubMed]
- Liraa, R.; MacAdam, J.; Sales, F.; Villalba, J.J. Supplementation strategies to enhance intake of romerillo (Chiliotrichum diffusum) by sheep in southern Patagonia. Small Rumin. Res. 2020, 192, 106205. [Google Scholar] [CrossRef]

{kind=link}
{kind=link}
| Parameters | S. lancea | S. pyroides | E. crispa |
|---|---|---|---|
| CP (%) | 9.5 ± 0.6 | 12.1 ± 0.8 | 7.5 ± 0.2 |
| NDF (%) | 42.3 ± 0.6 | 44.4 ± 1.1 | 34.8 ± 0.7 |
| ADF (%) | 22.1 ± 0.5 | 28.2 ± 0.4 | 25.3 ± 0.5 |
| ADL (%) | 7.5 ± 1.0 | 13.3 ± 1.5 | 13.5 ± 0.6 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Monegi, P.; Mkhize, N.R.; Masondo, P.T.; Mbatha, K.R.; Luseba, D.; Tjelele, J.T. Diet Mixing and Supplementation Present an Opportunity to Increase the Use of Encroaching Woody Plants by Goats. Animals 2023, 13, 3509. https://doi.org/10.3390/ani13223509
Monegi P, Mkhize NR, Masondo PT, Mbatha KR, Luseba D, Tjelele JT. Diet Mixing and Supplementation Present an Opportunity to Increase the Use of Encroaching Woody Plants by Goats. Animals. 2023; 13(22):3509. https://doi.org/10.3390/ani13223509
Chicago/Turabian StyleMonegi, Piet, Ntuthuko Raphael Mkhize, Purity Thobekile Masondo, Khanyisile Rebecca Mbatha, Dibungi Luseba, and Julius Tlou Tjelele. 2023. "Diet Mixing and Supplementation Present an Opportunity to Increase the Use of Encroaching Woody Plants by Goats" Animals 13, no. 22: 3509. https://doi.org/10.3390/ani13223509
APA StyleMonegi, P., Mkhize, N. R., Masondo, P. T., Mbatha, K. R., Luseba, D., & Tjelele, J. T. (2023). Diet Mixing and Supplementation Present an Opportunity to Increase the Use of Encroaching Woody Plants by Goats. Animals, 13(22), 3509. https://doi.org/10.3390/ani13223509