Simple Summary
In this study, the levels of infestation of two different species of pikeworm, Triaenophorus crassus and T. nodulosus, inhabiting a deep, oligotrophic mountain lake in Siberia were revealed. Based on results of five years of studies, a significant interannual variation in T. crassus infestation level was found, whereas the stability of T. nodulosus infestation in their host fishes was seen. Differences in feeding habits and physiology of fishesprobably had an effect on the mass of parasite abundance. Moreover, an asymmetry in parasite infestations between the number of T. crassus cysts in the left and right body surfaces of the ‘‘benthivorous” Coregonus lavaretus pidschian was also observed.
Abstract
In the present study, we estimated the levels of infestation of the main fish species that are hosts for two Triaenophorus species: T. crassus and T. nodulosus. The prevalence of T. crassus and T. nodulosus infestations in the intestine of their definitive host–pike Esox lucius was similar (71.0% and 77.4%, respectively). At the same time, the prevalence of T. crassus infestation in muscle tissue was significantly different between the second intermediate hosts, Coregonus lavaretus pidschian (31.4%) and Cor. l. pravdinellus (91.2%), due to considerable differences in their diets. For T. nodulosus, we found significant variations in the levels of prevalence among the second intermediate hosts—100% for Lota lota, 81.8% for Cottus sibiricus 31.9% for Thymallus arcticus, and 24.5% for Perca fluviatilis—that we also explained using different diets. Moreover, analysis of the symmetry of parasite infestations did not reveal any asymmetry between the number of cysts in the left and right body surfaces of the “planktivorous” form/species of whitefish, whereas in the ‘‘benthivorous”, an asymmetry of parasite infestations was found.
1. Introduction
The majority of helminths have complex lifecycles that can include one or several intermediate hosts and are transmitted through food webs [1]. Different fish species are characterized by very diverse feeding habits and strategies while occupying different positions in aquatic food webs. At the same time, fishes may consume a plethora of different species of invertebrates that are common first intermediate hosts for helminths [1]. Due to the species-specific ranges of hosts for various helminths and different feeding habits among fish species, the parasites found in or on fish are used as one of the trophic indicators that may clarify the trophic relationships among species in aquatic food webs. The role of parasites as indicators of trophic diversification of fish has been discussed in recent studies [1,2,3,4]. In general, a high level of fish infestation positively correlates with high consumption of infected food items that are the first intermediate host for transmitted helminths. Moreover, the specific parasite found in a fish host may indicate that this fish consumes specific food items, even if this food item was never registered by other approaches like direct observation of gut content [4].
Tapeworms of the genus Triaenophorus (Cestoda, Bothriocephalidea: Triaenophoridae) are widespread and highly specialized parasites of Holarctic fish. According to a recent study based on sequencing of partial cox1 mtDNA and nuclear 28S rRNA genes, the Eurasian pikeworm Triaenophorus spp. consists of five valid species (T. amurensis Kuperman, 1968; T. crassus Forel, 1868; T. meridionalis Kuperman, 1968; T. nodulosus Pallas, 1781, Rudolphi, 1793; and T. orientalis Kuperman, 1968) [5]. The life cycle of these cestodes is complex and proceeds with the passage through two intermediate hosts and one definitive host. Cestodes become sexually mature only in the intestines of pike Esox lucius Linnaeus, 1758 after feeding on fishes infected with these cestodes [6].
Infestations with plerocercoids and adults of different Triaenophorus species are particularly common in freshwater fishes. The geographic distribution of the Triaenophorus species are circumpolar and includes more than 70 fish species that have been described as possible second intermediate and definitive hosts [5,6,7]. For freshwater bodies of West Siberia (Russia), two species, T. crassus and T. nodulosus, are of epizootic importance. Triaenophorus cestodes do not present a health hazard to humans, but these parasites can cause financial damage to fisheries due to the unappetizing appearance of infested fish. T. crassus and T. nodulosus differ in morphological features and localization in the second intermediate hosts. The main morphological differences of T. crassus from T. nodulosus are the shape of the scolex hooks, which have a massive basal part and small straight prongs [5,6]. The number of second intermediate hosts for T. crassus is much lower than for T. nodulosus; some hosts are common to both species. The plerocercoid of T. crassus is localized in the musculature of fish from the subfamilies Salmoninae and Coregoninae, while T. nodulosus is localized in the liver of Percidae, Thimallinae, Cobitidae, Osmeridae, Cyprinidae, Cottiidae, Lotidae, and several other families and subfamilies of fishes [6].
Several fish species inhabiting Teletskoye Lake (Altai Republic, Western Siberia, Russia) are known as common hosts for these two Triaenophorus species. Thus, T. nodulosus is commonly found in the liver of Siberian sculpin Cottus sibiricus Kessler, 1889; perch Perca fluviatilis Linnaeus, 1758; arctic grayling Thymallus arcticus Pallas, 1776; and burbot Lota lota Linnaeus, 1758 [5,8]. While T. crassus typically parasitizes muscle tissue of a sympatric pair of coregonid whitefishes—the planktivorous “dwarf” form/species named Coregonus lavaretus pravdinellus Dulkeit, 1949 and the benthivorous “normal” form/species named Cor. l. pidschian Gmelin, 1789 [5,9]. The definitive host for both Triaenophorus species in the lake is pike, Esox lucius Linnaeus, 1758 where these helminths are found in the intestine.
Teletskoye Lake is one of the biggest montane, oligotrophic, deep-water bodies located in the south of Siberia that is a refugium since the last glaciations for several fish species including endemic whitefish—Cor. l. pravdinellus. Although the main host ranges are known for both Triaenophorus species in this lake, previous studies have provided very limited information regarding the ecological features of these parasites such as transmissions in the food web of the lake, distribution of parasites in different parts of the fish body, interannual variability of infestation levels, etc.
Thus, the main aim of the present study was to estimate the infestation levels of both Triaenophorus species in the second intermediate and the definitive hosts from Teletskoye Lake. We tested the following three hypotheses: (1) the main driver that determines the levels of infestation in second and definitive hosts is feeding habits; this factor does not directly reflect the expected route of parasite transmission, e.g., planktonic crustaceans (first intermediate host), planktivorous fish (second intermediate host), pike (definitive host); (2) the infestation levels of pike, as the definitive host, will be different for each Triaenophorus species due to the studied Triaenophorus species having different life strategies: T. crassus infects only two secondary host fish species, whereas T. nodulosus is able to infect at least six fish species as the second intermediate host; and (3) the body mass of the studied Triaenophorus plerocercoids will be different in various second intermediate hosts.
2. Materials and Methods
2.1. Sample Collection
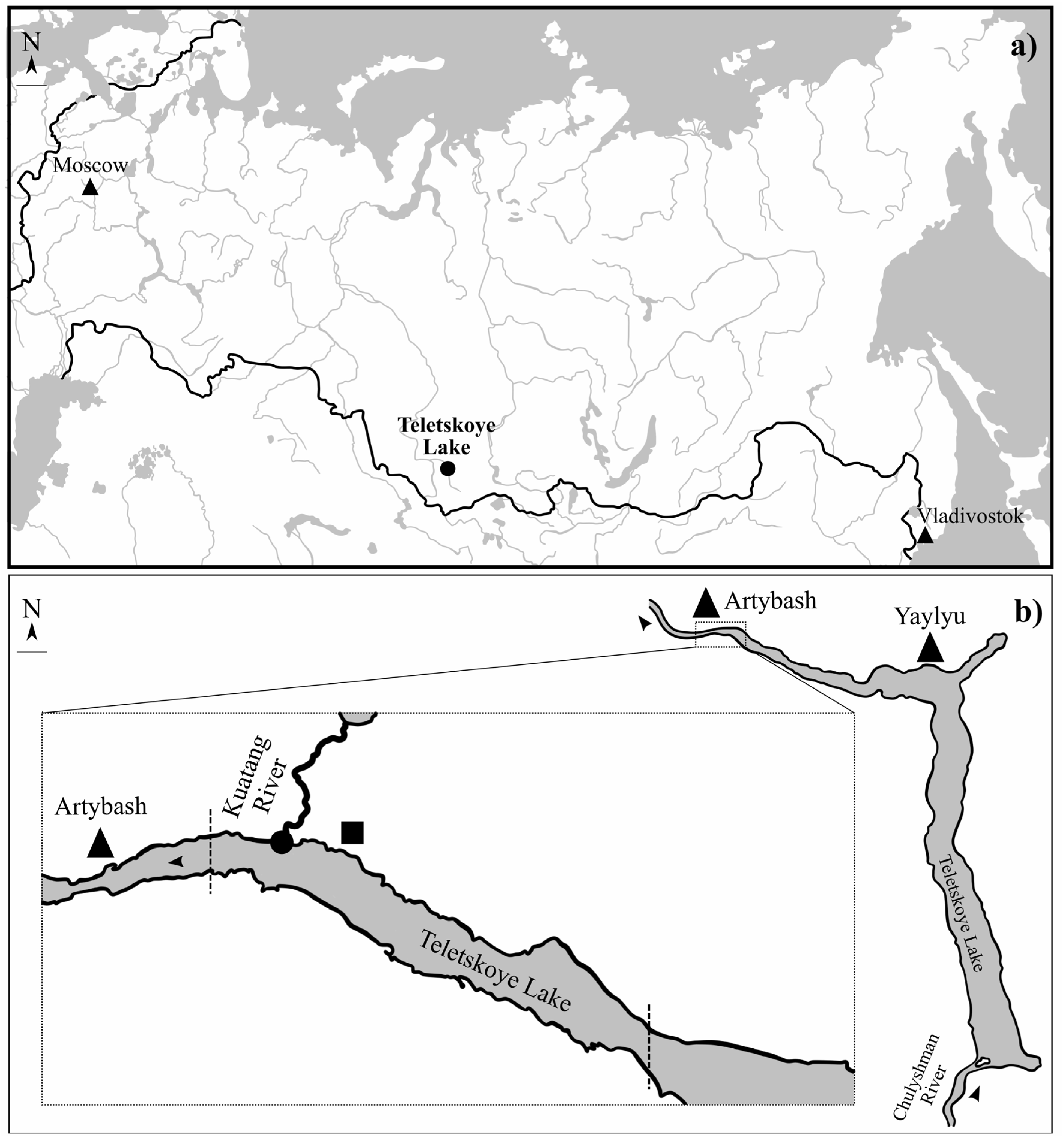
A total of 507 fishes, belonging to (Coregoninae, Cottidae, Percidae, Gadidae, Cyprinidae were investigated (Table 1). The fishes (with exception common minnow Phoxinus phoxinus Linnaeus, 1758) were collected at the end of August and the end of October between 2019 and 2022 in the north part of Teletskoye Lake (51°79′ N; 87°30′ E). Teletskoye Lake is a large (223 km2) and deep (maximum 325 m) oligotrophic lake (basin of Ob River) in the Altai Mountains (Altai Republic, Russia). Ph. phoxinus was collected from a small river, Kuatang, that flows into Teletskoye Lake. Fish from the lake were captured using gill nets (mesh sizes 18–60 mm) at depths from 2 to 40 m. Ph. Phoxinus was collected using a fish net at a depth 0.5 m (Figure 1). After capture, all fish were transported to the laboratory in the Teletskoye Lake field station of the Institute of Systematic and Ecology of Animals SB RAS.

Table 1.
Summary data of total body weight (BW), eviscerated body weight (EBW), total length (TL), sex ratio, and fish diet of seven fish species examined for presence of Triaenophorus crassus and T. nodulosus from Teletskoye Lake through different years. * F—female, M—male.
Figure 1.
General map of Russian Federation (a) and map of the Teletskoye Lake with sampling point and area (b): dot—sampling point of Phoxinus phoxinus; dotted lines—sampling area of other fishes; triangle—village; rectangle—field station; arrow—water flow direction.
The fishes were humanely sacrificed by a blow to the head before sampling. Afterwards, the fishes were identified, measured (total length, TL), and weighed (total body weight, or “BW”, and eviscerated body weight, or “EBW”). Male and female fishes were identified according to gonadal development. The total number of fishes collected for study included: 140 “normal” whitefish Cor. l. pidschian and 89 “dwarf” whitefish Cor. l. pravdinellus, 55 Siberian sculpin Cot. sibiricus, 102 perch Pe. fluviatilis, 47 arctic grayling Th. arcticus, 35 burbot L. lota, 8 common minnow Ph. phoxinus, and 31 pike E. lucius.
2.2. Parasitological Analysis
Plerocercoids were removed from cysts and stored in 70% ethanol. The scolex hooks were measured on the squashed scolices mounted in Berlese’s medium. The species identification of the cestodes was performed in accordance with identification keys using characters described earlier [6,10]. The prevalence and mean intensity of parasite infestation were calculated according to the standard definitions described by Bush et al. [11]. In addition, the wet weight of plerocercoids/adult worms and wet weight of fish livers and intestines (E. lucius) were measured using an electronic scale SCOUT™ STX (Göteborg, Sweden). The ratio of the wet weight of plerocercoids/adult worm from wet weight of infected organ (liver or intestine), total body weight of fish and eviscerated body weight of fish were calculated.
2.3. Statistical Analysis
All data are presented as a mean ± standard error (SE). To estimate the differences between parasite intensity and abundance across different sampling years (2019–2022), as well as in terms of fish total length and body weight among different sampling years, the Tukey HSD post-hoc test was applied using PAST v. 3.16 [12]. In the same software, to explore the effect of gender (sex) as a factor on the parasite abundance and intensity, this factor was tested using one-way ANOVA with statistical significance established at p < 0.05. To estimate the level of correlations between parasite intensity, abundance and fish total length, total body weight, and gender (sex), a Spearman rank correlation test (ρ) was used. The relationship between the number of T. crassus and T. nodulosus individuals and the total length or total body weight of definitive host—E. lucius—were estimated using the linear regression realized in Statistica 8.0. In the same software, the relationship between the weight of T. crassus and T. nodulosus and number of cysts in different intermediate hosts were estimated. Differences of slopes of two regression lines in terms of total length and body weight of E. lucius were calculated using the Student’s t-test implemented in Excel MS 2016. Density plots were created using ggplot2 R package [13]. To characterize the distribution, the aggregation indices were calculated using QPweb Version 1.0.15 [14]. To analyze the symmetry of T. crassus infestations in different forms/species of whitefishes Coregonus lavaretus a binomial exact test was performed using aspi R package in R software version 4.1.2 (R Core Team, 2021) [15].
3. Results
3.1. Prevalence and Intensity of Fish Infestation with Triaenophorus spp.
3.1.1. Intermediate Hosts Infected by T. crassus
T. crassus cysts was found in muscle of both forms/species of whitefishes Coregonus lavaretus. A single cyst from muscle contained one plerocercoid. The prevalence of T. crassus infestation in the muscles of the “dwarf” whitefish Cor. l. pravdinellus was higher than that in the muscles of the ‘‘normal” whitefish Cor. l. pidschian for all studied years and ranged from 89.7 to 100% and from 27.3 to 41.5%, respectively (Table 2).
Table 2.
Infestation indices of T. crassus and T. nodulosus in different intermediate and definitive hosts from Teletskoye Lake.
The mean intensity and abundance of parasite infestation in the muscles of the “dwarf” whitefish Cor. l. pravdinellus ranged from 4.5 ± 0.5 to 5.0 ± 0.8 worms and from 4.0 ± 0.5 to 5.0 ± 0.8 worms, respectively, whereas the same indices of parasite infestation in the muscles of the “normal” whitefish Cor. l. pidschian ranged from 1.8 ± 0.4 to 1.9 ± 0.5 worms and from 0.5 ± 0.2 to 0.8 ± 0.2 worms, respectively (Table 2).
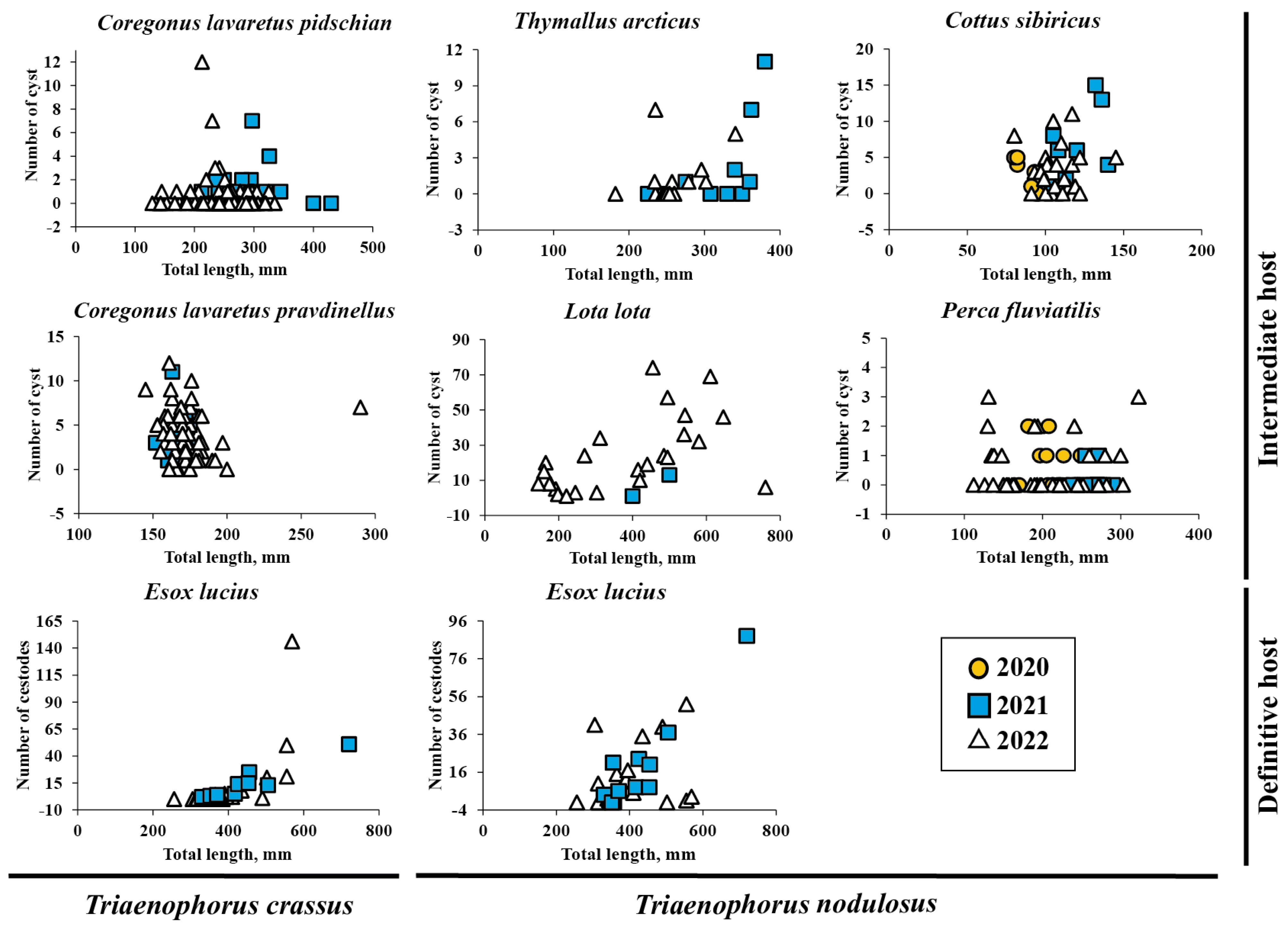
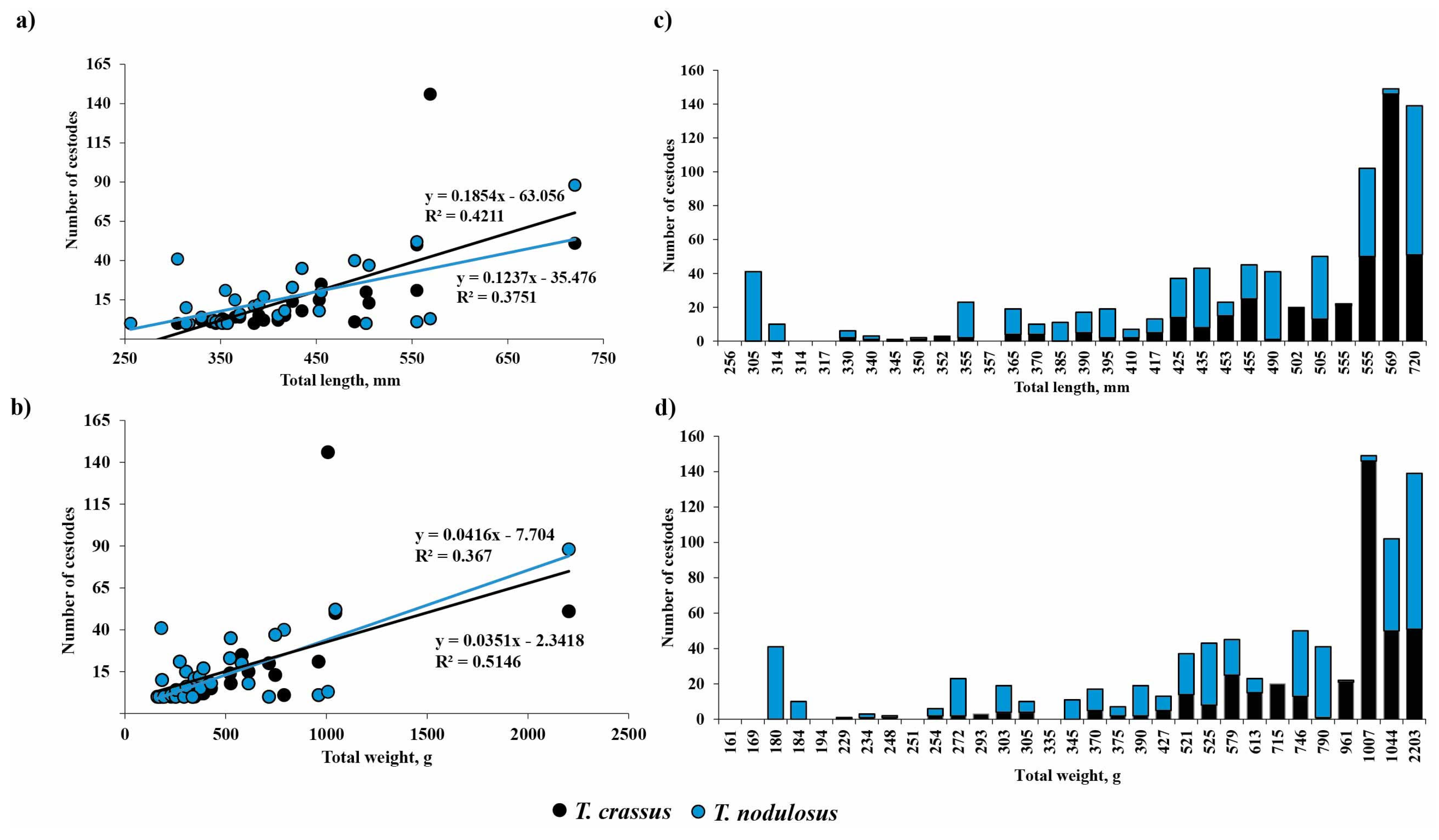
The abundance of T. crassus compared with the total length of studied fishes is shown in Figure 2. There was no difference in the mean intensity and abundance of T. crassus infecting the muscles of both forms/species of whitefishes Coregonus lavaretus between the studied years, 2021 and 2022 (“dwarf”: ANOVA (intensity), F = 0.28, df = 35.8, p = 0.60, ANOVA (abundance) F = 1.1, df = 34.6, p = 0.31; ‘‘normal”: ANOVA (intensity), F = 0.002, df = 41.9, p = 0.96, ANOVA (abundance) F = 0.96, df = 82.2, p = 0.33).
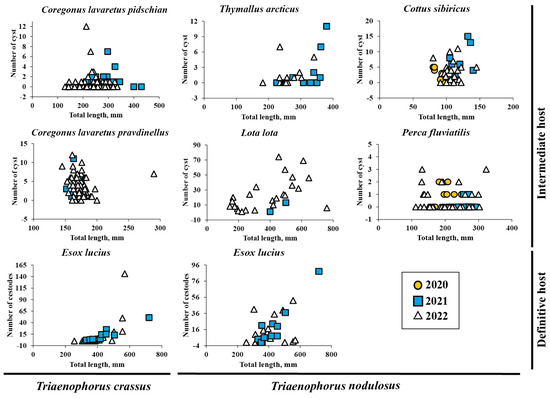
Figure 2.
The abundance of Triaenophorus crassus and T. nodulosus contrasted with total length of different intermediate and definitive hosts from Teletskoye Lake from 2020 to 2022.
A positive significant correlation was found between total body weight of the “dwarf” whitefish Cor. l. pravdinellus and the number of cysts in muscle tissue in 2021 (Spearman ρ = 0.55, p = 0.01, n = 20). In other cases, no statistically significant correlations were found between the number of cysts in muscle and both length and weight of “dwarf” and ‘‘normal” forms/species of whitefishes Coregonus lavaretus.
The factor “sex” had no effect on the number of cysts T. crassus in fish muscles (“dwarf”: Spearman ρ = −0.05, p = 0.86, n = 16, 2021; Spearman ρ = −0.11, p = 0.43, n = 52, 2022; ‘‘normal”: Spearman ρ = 0.07, p = 0.21, n = 30, 2021; Spearman ρ = −0.19, p = 0.08, n = 83, 2022).
3.1.2. Intermediate Hosts Infected by T. nodulosus
T. nodulosus was identified in the liver of Cot. sibiricus, Pe. fluviatilis, Th. arcticus, and L. lota. A single cyst from the liver contained one plerocercoid. In only one fish species (Ph. phoxinus), were there no plerocercoids found in their liver. The prevalence of T. nodulosus infestation in the liver of L. lota (100% for years 2021 and 2022) was higher than that in the liver of Cot. sibiricus, Th. arcticus, Pe. fluviatilis, where the prevalence level ranged from 66.7 to 100%, 30.4 to 33.3%, and 20.0 to 27.3%, respectively, for those same years (Table 2). The mean intensity of parasite infestation in the liver of L. lota, Cot. sibiricus, Th. arcticus, and Pe. fluviatilis ranged from 7.4 ± 2.3 to 23.5 ± 3.7, 3.6 ± 0.8 to 8.0 ± 2.3, 2.6 ± 0.8 to 4.1 ± 1.3, and 1.0 ± 0.0 to 1.8 ± 0.2, correspondingly (Table 2).
The mean abundance of parasite infestation in the liver of L. lota, Cot. sibiricus, Th. arcticus, and Pe. fluviatilis ranged from 7.4 ± 2.3 to 23.5 ± 3.7, 2.4 ± 0.8 to 8.0 ± 2.3, 0.8 ± 0.1 to 1.4 ± 0.6, and 0.2 ± 0.1 to 0.5 ± 0.1, respectively. The abundance of T. nodulosus compared with the total length of studied fishes is shown in Figure 2. The mean intensity and abundance of T. nodulosus infecting the liver of fishes were not significantly different between studied years (2019–2022). There was one exception for Cot. sibiricus, where the significant differences in mean abundance was obtained between the years 2020 (8.0 ± 2.3) and 2021 (3.3 ± 0.6) (ANOVA, Tukey HSD post-hoc test, F = 4.2, df = 3, p = 0.013) and between the years 2021 and 2022 (2.4 ± 0.8) (ANOVA, Tukey HSD post-hoc test, F = 4.2, df = 3, p = 0.015).
A positive significant correlation was found between L. lota total body weight/total length and the number of cysts in its liver in 2022 (total length: Spearman ρ = 0.61, p = 0.0006, n = 30), (total weight: Spearman ρ = 0.60, p = 0.002, n = 25). A positive significant correlation was also found between total length of the Th. arcticus and the number of cysts in its liver in 2021 (Spearman ρ = 0.54, p = 0.020, n = 19). Throughout all of the studied years, a positive significant correlation was found between the total length/weight of the Th. arcticus and the number of cysts (total length: Spearman ρ = 0.37, p = 0.020, n = 38; total weight: Spearman ρ = 0.39, p = 0.019, n = 36). In other cases, there were no statistically significant correlations found between the number of cysts in liver and both length and weight of fishes. The factor “sex” had no effect on the intensity and abundance of T. nodulosus in fish liver.
3.1.3. Distribution of T. crassus and T. nodulosus Plerocercoids
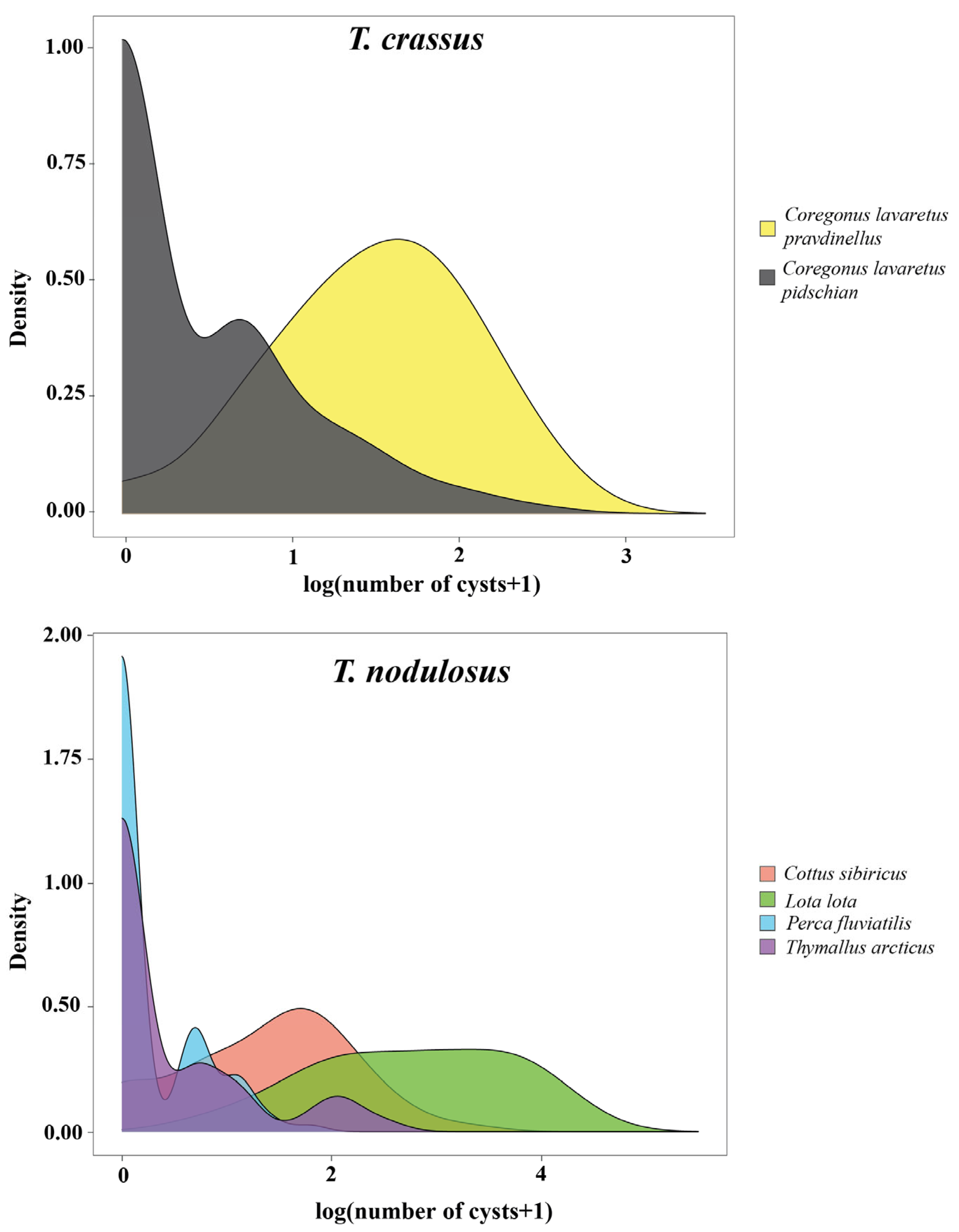
The distribution patterns of T. crassus plerocercoids in two forms/species of whitefishes Coregonus lavaretus were strictly different (Figure 3). Parasite aggregation was significantly higher for Cor. l. pidschian in comparison with Cor. l. pravdinellus (Poulin’s discrepancy index were amounted to 0.756 and 0.391, respectively). The variance/mean ratio differed to a lesser extent and amounted to 3.17 and 2.70, respectively. Cor. l. pidschian infestations were characterized by the presence of a large number of uninfected individuals and at low intensity, while plerocercoids from Cor. l. pravdinellus were more evenly distributed (Figure 3).
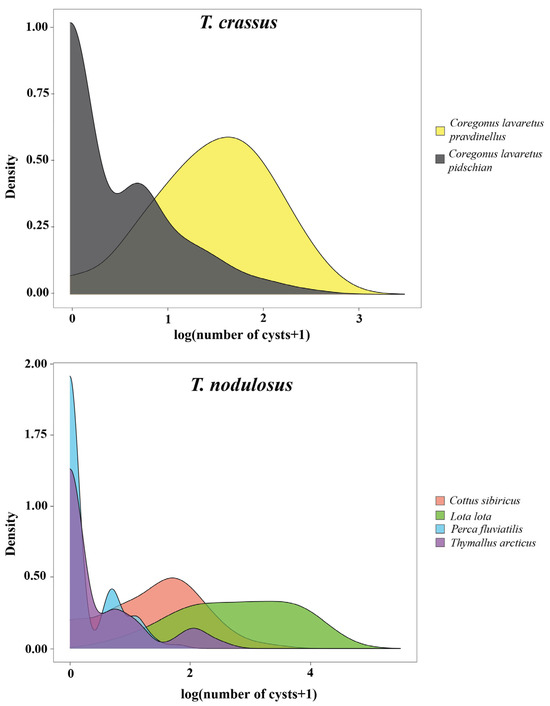
Figure 3.
Distribution plot of the number of T. crassus and T. nodulosus plerocercoids per host species.
The highest Poulin’s discrepancy index for T. nodulosus was shown for Th. arcticus (0.809), the lowest one for L. lota (0.454). The variance/mean ratio was minimal in Pe. fluviatilis and maximal in L. lota (1.71 and 17.36, respectively). The shape of distribution of T. nodulosus was similar for both Pe. fluviatilis and Th. arcticus. Cot. sibiricus and L. lota were characterized by a small number or absence of uninfected individuals and higher values of intensity. In general, the variance/mean ratio values of more than one indicated an aggregation of plerocercoids among hosts and preclude a random or uniform distribution.
3.1.4. Definitive Host
The prevalence of T. crassus infestation in the intestine of E. lucius was higher in 2022 than that in 2021 and amounted to 78.6 and 64.7%, respectively, whereas the prevalence of T. nodulosus in the intestine of E. lucius did not change during the studied years (76.5 and 78.6% for 2021 and 2022 years) (Table 2). The mean intensity and abundance of T. crassus infestation in the intestine of E. lucius ranged from 12.3 ± 5.6 to 23.6 ± 6.3 worms and 9.6 ± 3.8 to 15.3 ± 8.8 worms, respectively, whereas the mean intensity and abundance of T. nodulosus ranged from 18.8 ± 4.8 to 19.7 ± 6.8 worms and 14.4 ± 4.1 to 15.5 ± 6.3 worms, respectively (Table 2). The mean intensity and abundance of T. crassus and T. nodulosus infecting the intestine of E. lucius were not different between studied years (ANOVA (intensity), F = 0.68, df = 1.0, p = 0.42, T. crassus; ANOVA (abundance), F = 0.3, df = 1.0, p = 0.59, T. crassus; ANOVA (intensity), F = 0.01, df = 1.0, p = 0.91, T. nodulosus; ANOVA (abundance), F = 0.03, df = 1.0, p = 0.88, T. nodulosus).
Mean abundance of T. crassus in the intestine of E. lucius significantly increased with increasing total length and weight of fish (Figure 4, TL: R2 = 0.42, F = 0.0001, n = 30; TW: R2 = 0.37, F = 0.0003, n = 31). There are no differences for the slopes of the two regression lines in terms of length and weight of fish.
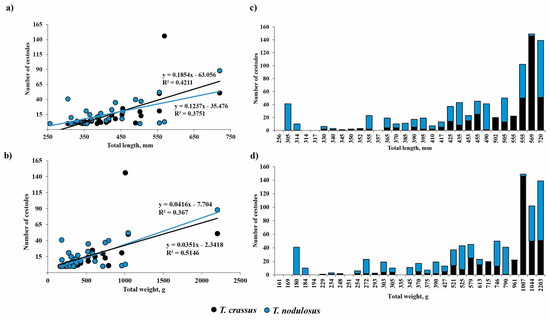
Figure 4.
Relationship between the number of Triaenophorus crassus and T. nodulosus and total length (a,c) or total body weight (b,d) of definitive host—Esox lucius—from Teletskoye Lake during studied years (2021, 2022). The black and blue lines on Figure (a,b) show the linear regression with R-squared for T. crassus and T. nodulosus, correspondingly.
No sex-related differences of mean abundance (R2 = 0.034, F = 0.397, n = 23) were found for T. crassus in the intestine of E. lucius. The mean abundance of T. nodulosus in E. lucius intestine had also significantly increased with fish total length and total bodyweight (Figure 4a,b, TL: R2 = 0.38, F = 0.0003, n = 30; TW: R2 = 0.51, F = 5.59 × 10−6, n = 31). A significant relationship was also found between mean abundance of T. nodulosus and gender (sex) of E. lucius (R2 = 0.28, F = 0.009, n = 23). In the group of fish with total length from 250 to 555 mm and total body weight from 150 to 800 g, the number of T. nodulosus in the intestine of E. lucius was higher in comparison with number of T. crassus, whereas in the larger fish group (TL and TW ranged from 555 to 1000 mm and from 1000 to 2300 g, respectively), the ratio of T. crassus was replaced by T. nodulosus and reached a maximum of 146 and 88 cestodes in one pike specimen (Figure 4c,d).
3.2. The Relationship between Wet Weight of Cestodes and Their Hosts
3.2.1. Intermediate Hosts Infected by T. crassus
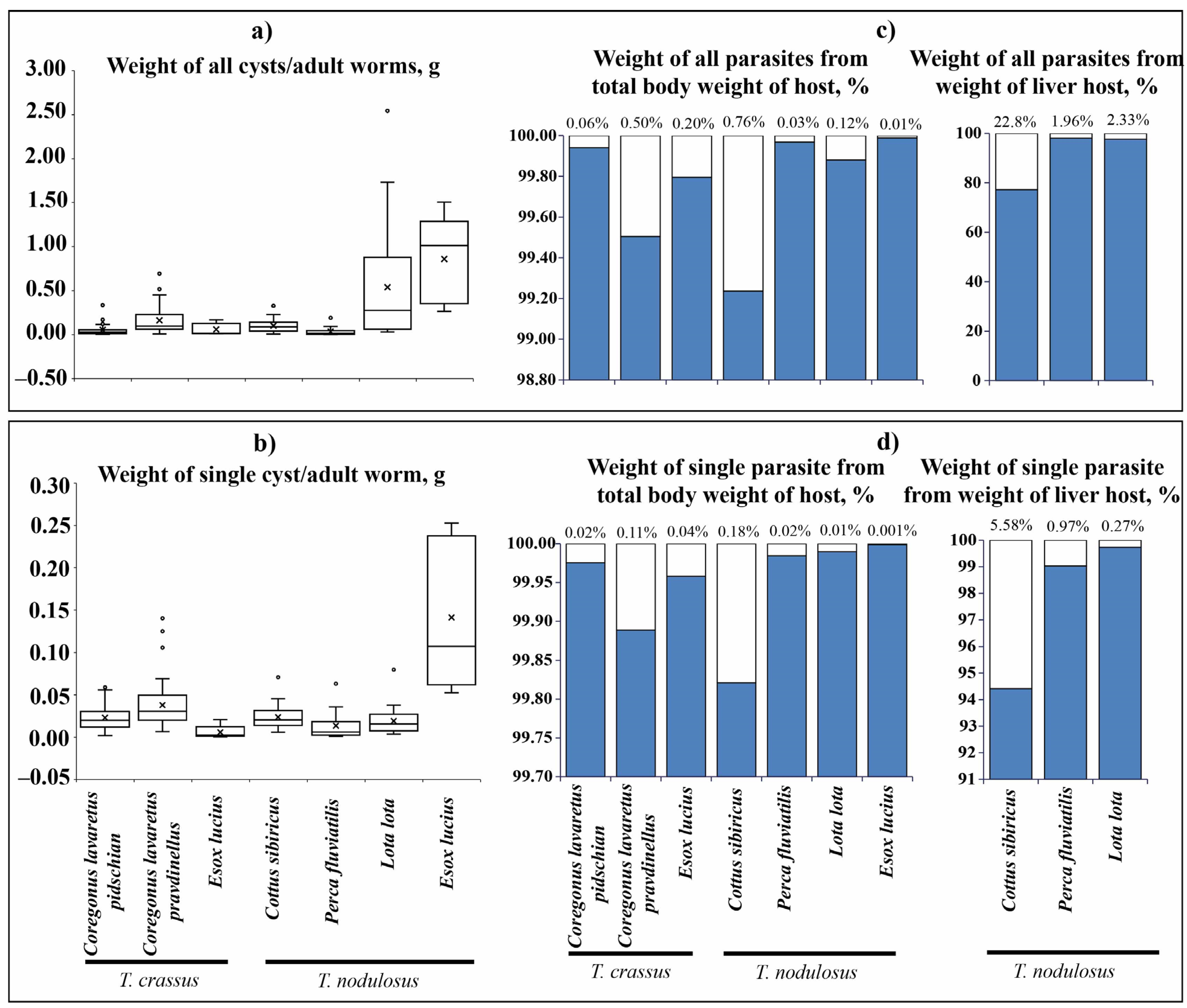
The total wet weight of T. crassus plerocercoids infecting the muscles of both forms/species of whitefishes Coregonus lavaretus ranged from 0.002 to 0.693 g (Figure 5a). The mean wet weight of all T. crassus plerocercoids from muscle of the “dwarf” form/species Cor. l. pravdinellus (0.161 ± 0.019 g) was significantly higher if compared to the ‘‘normal” whitefish Cor. l. pidschian (0.054 ± 0.011 g) (ANOVA, F = 17.1, df = 1.0, p = 0.00008). The mean wet weight of individual specimens of T. crassus plerocercoids from muscles of “dwarf” form/species Cor. l. pravdinellus (0.04 ± 0.002 g) was also significantly higher when compared to the ‘‘normal” (0.02 ± 0.003 g) whitefish Cor. l. pidschian (ANOVA, F = 10.1, df = 1.0, p = 0.002) (Figure 5b). The wet weight of T. crassus plerocercoids did not exceed one percent of the host total weight and weight eviscerated (Figure 5c,d; Figure S1). A positive significant correlation was found between the number of T. crassus cysts and their mean wet weight of cysts. In case of the average individual wet weight of cysts and number of cysts, there were no statistically significant correlations found (Table 3).
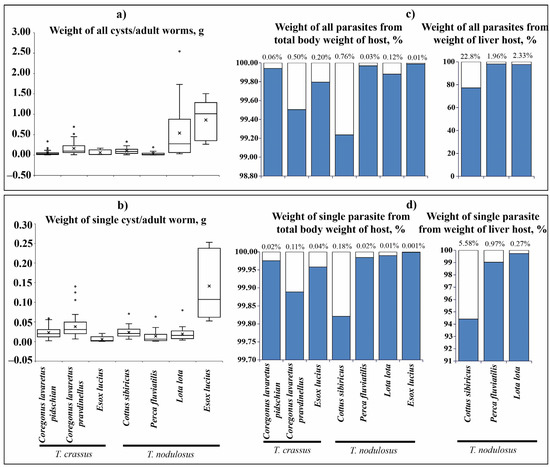
Figure 5.
Size of cestodes ( ) and their relationship with fish total body weight (
) and their relationship with fish total body weight ( ). (a) Weight of all cysts or adult worms, g; (b) weight of single cyst or adult worm, g; (c) weight of all cysts or adult worms from total body weight, and weight of liver of fish, %; (d) weight of single cyst or adult worm from total body weight, and weight of liver of fish, %. Boxplots represent mean (crosses) with outliers (empty circles). The percent on the top of graph (c,d) indicates the weight of parasites.
). (a) Weight of all cysts or adult worms, g; (b) weight of single cyst or adult worm, g; (c) weight of all cysts or adult worms from total body weight, and weight of liver of fish, %; (d) weight of single cyst or adult worm from total body weight, and weight of liver of fish, %. Boxplots represent mean (crosses) with outliers (empty circles). The percent on the top of graph (c,d) indicates the weight of parasites.
) and their relationship with fish total body weight (). (a) Weight of all cysts or adult worms, g; (b) weight of single cyst or adult worm, g; (c) weight of all cysts or adult worms from total body weight, and weight of liver of fish, %; (d) weight of single cyst or adult worm from total body weight, and weight of liver of fish, %. Boxplots represent mean (crosses) with outliers (empty circles). The percent on the top of graph (c,d) indicates the weight of parasites.
Table 3.
Relationship between the wet weight of T. crassus and T nodulosus and number of cysts in different intermediate hosts from Teletskoye Lake.
3.2.2. Intermediate Hosts Infected by T. nodulosus
The total wet weight of T. nodulosus infecting the liver of intermediate hosts (Cot. sibiricus, L. lota, and Pe. fluviatilis) ranged from 0.001 to 2.543 g (Figure 5a). For T. nodulosus from different hosts, the highest total wet weight of plerocercoids was registered from L. lota (0.538 ± 0.111 g) in comparison with Cot. sibiricus (0.100 ± 0.015 g), and Pe. fluviatilis (0.035 ± 0.014 g) (ANOVA, Tukey HSD post-hoc test, F = 10.7, df = 65.0, p < 0.05). The highest individual weight of T. nodulosus was registered from the liver of Cot. sibiricus (0.024 ± 0.003), whereas from L. lota (0.019 ± 0.002 g) and Pe. fluviatilis (0.014 ± 0.005) values were lower, though insignificant (ANOVA, Tukey HSD post-hoc test, F = 1.9, df = 65.0, p > 0.05) (Figure 5b).
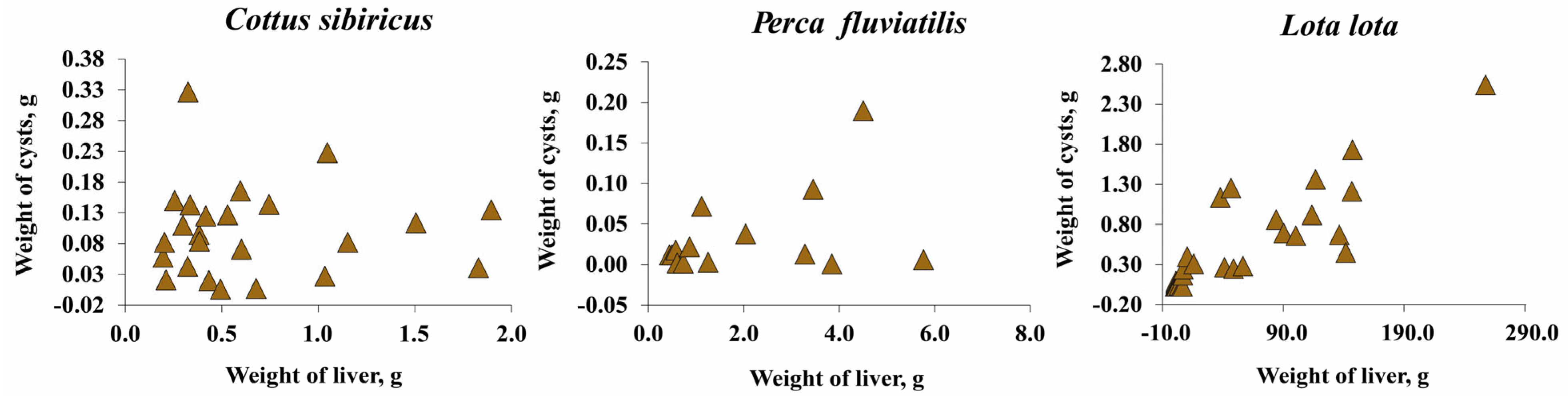
The total wet weight of T. nodulosus plerocercoids did not exceed one percent of the host total body weight and eviscerated body weight of fish (Figure 5c,d; Figure S1). The total wet weight of T. nodulosus plerocercoids amounted to 22.8 ± 4.6, 1.9 ± 0.1, and 2.3 ± 0.5 % of the liver weight of Cot. sibiricus, Pe. fluviatilis, and L. lota, respectively (Figure 5c). The weight of individual specimens of T. nodulosus plerocercoids amounted to 5.6 ± 1.0, 1.0 ± 0.3, and 0.3 ± 0.1% of the liver weight of Cot. sibiricus, Pe. fluviatilis, and L. lota, respectively (Figure 5d). The scatter plots of all cysts weight to weight of the liver of host are shown in Figure 6.
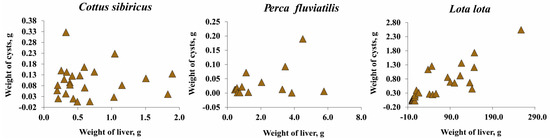
Figure 6.
The wet weight of all cysts of Triaenophorus nodulosus with weight of liver of different intermediate hosts from Teletskoye Lake.
A positive significant correlation was found between the number of T. nodulosus cysts and their total weight as well as to relation of the average individual weight of cysts and number of cysts with the exception of Cot. sibiricus (Table 3).
3.2.3. Definitive Host
The total and individual weight of adult specimens of T. nodulosus (0.86 ± 0.23 and 0.14 ± 0.04 g, correspondingly) was significantly higher than T. crassus (0.06 ± 0.03 and 0.006 ± 0.004 g, correspondingly) parasitizing the same intestine of E. lucius (Figure 5a,b, total weight, ANOVA: F = 12.4, df = 1.0, p = 0.008; individual weight, ANOVA: F = 11.0, df = 1.0, p = 0.011). The total weight of adult T. nodulosus and T. crassus amounted to 9.1 ± 0.5 and 2.7 ± 0.2% of the weight of E. lucius intestine, respectively. Whereas the weight of individual specimens of adult T. nodulosus and T. crassus amounted to 1.7 ± 0.05 and 0.8 ± 0.03% % of the weight of E. lucius intestine, respectively.
3.3. Clinical Signs of Infestation and Localization of T. nodulosus and T. crassus in Fish
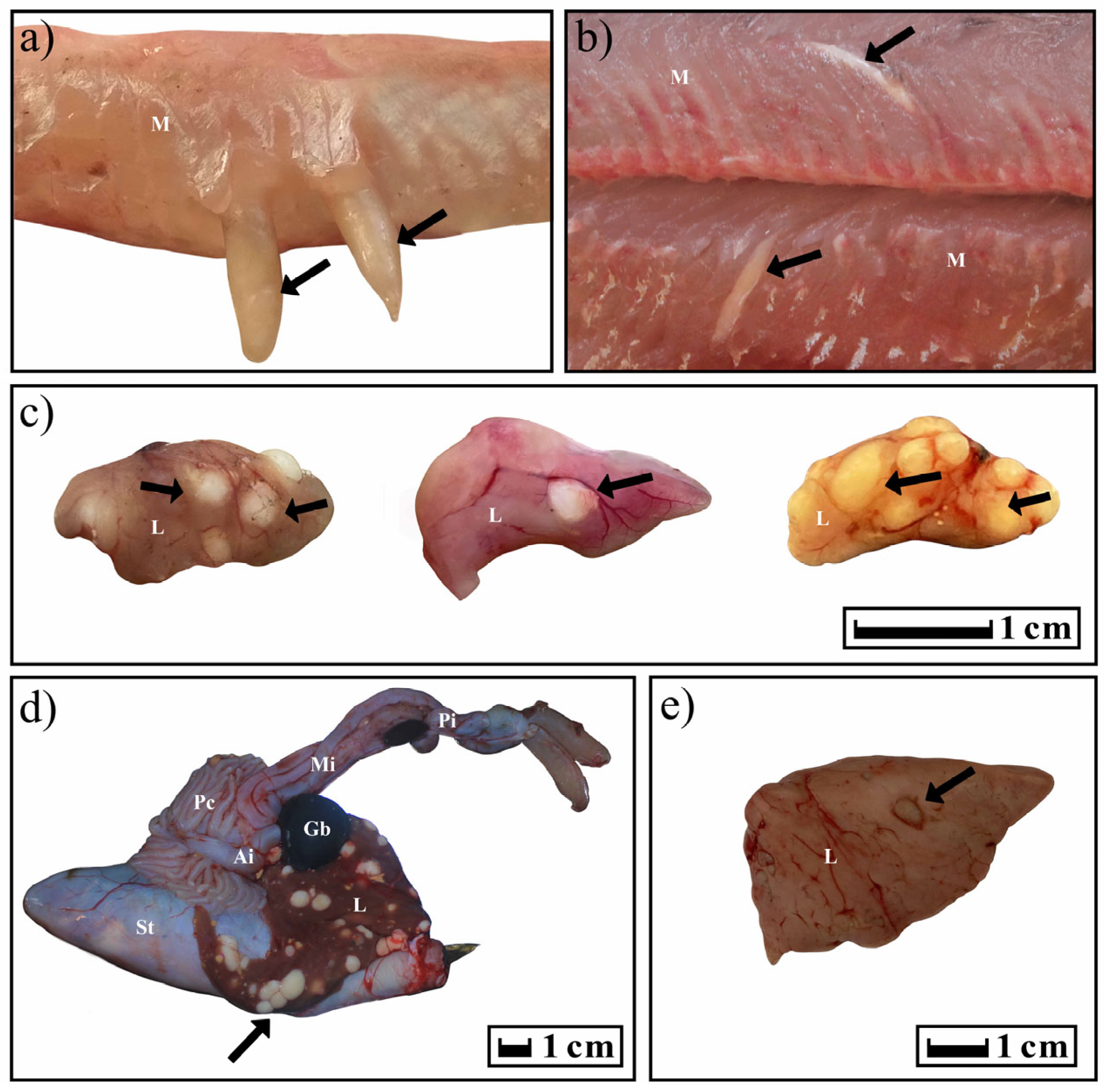
In the case of T. crassus plerocercoids, cone-shaped formations under the skin and in the muscles were clearly visible in infected fish. In fish infected with T. nodulosus plerocercoids, the liver in several cases was lighter than usual. Sexually mature cestodes caused mechanical damage to the E. lucius intestine, accompanied by the growth of connective tissue around the scolexes. In other cases, the fish looked conditionally healthy.
The typical distribution of T. nodulosus and T. crassus plerocercoids in fishes from Teletskoye Lake is shown in Figure 7. For years 2019–2020, atypical localization of T. nodulosus plerocercoids was noted on the wall of the intestine (L. lota) and body cavity (Th. arcticus and Cot. sibiricus). In an exceptional case, an immature T. nodulosus was observed in the intestine of one single L. lota.
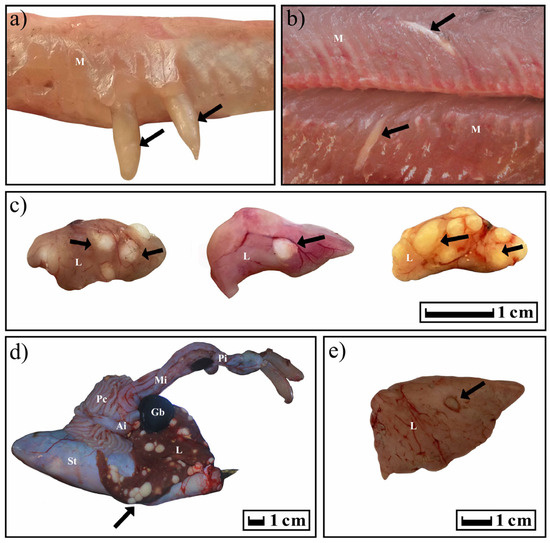
Figure 7.
Localization of T. crassus and T. nodulosus plerocercoids in different intermediate hosts. Black arrows show the localization of cysts. (a,b) Cor. l. pidschian; (c) Cot. sibiricus; (d,e) L. lota. Abbreviation: St—stomach; L—liver; Ai—anterior intestine; Mi—middle intestine; Pi—posterior intestine; Pc—pyloric caeca; Gb—gallbladder; M—muscles.
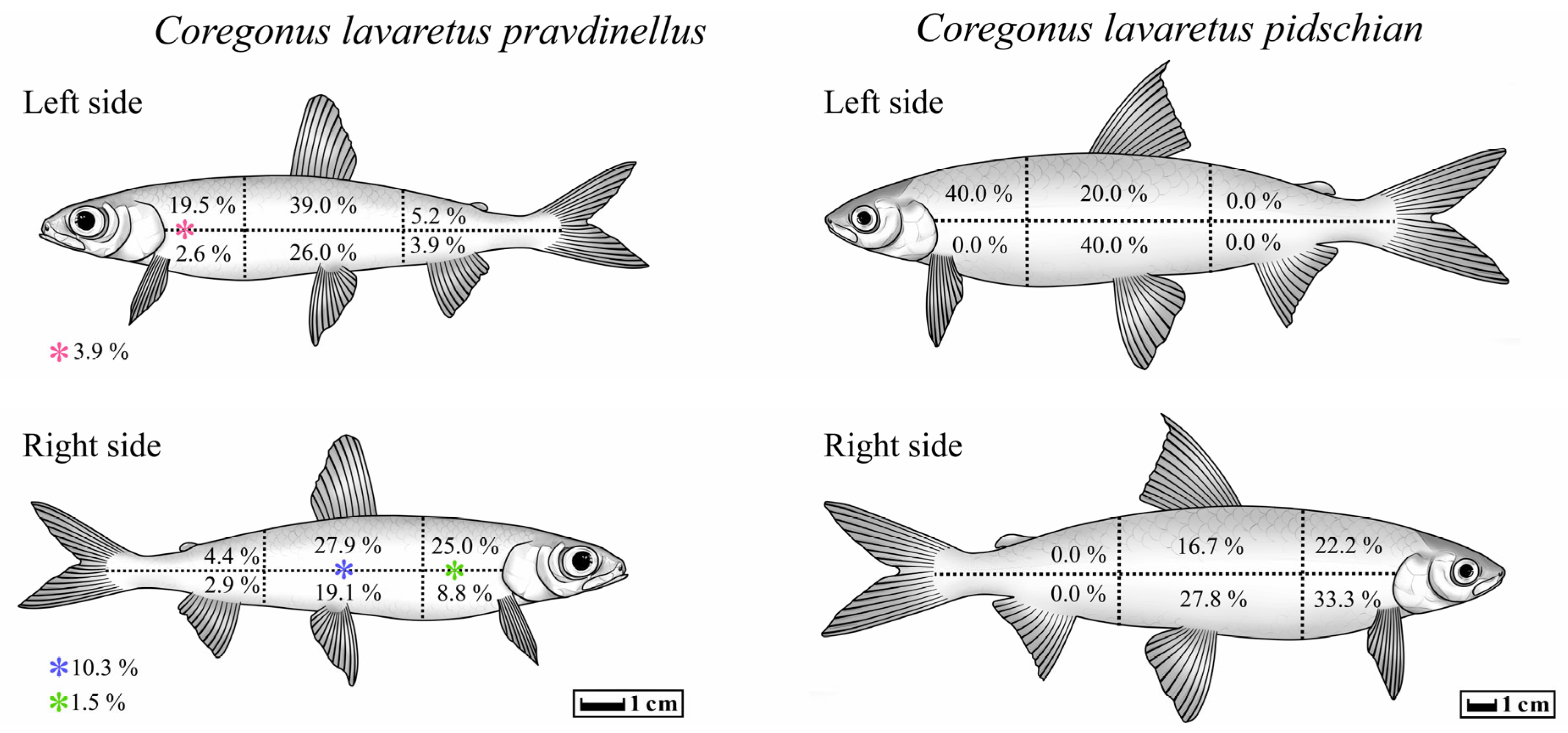
In five cases out of all studied forms/species of Coregonus lavaretus whitefishes, an atypical localization of T. crassus plerocercoids was observed only for the ‘‘normal” whitefish Cor. l. pidschian. Four capsules with plerocercoids were located in the liver, and another one on the surface of the stomach. In the ‘‘normal” whitefish Cor. l. pidschian the cysts were more numerous on the right side of the body (n = 18) than on the left (n = 5), whereas in the “dwarf” whitefish Cor. l. pravdinellus, the cysts were more numerous on the left side of the body (n = 77), than on the right (n = 68). Analysis of symmetry of parasite infestations (binomial exact test) did not reveal any asymmetry (p = 0.51) between the number of cysts in the left and right body surfaces of the “dwarf” whitefish Cor. l. pravdinellus, whereas in the ‘‘normal” whitefish Cor. l. pidschian, an asymmetry of parasite infestations was shown with p = 0.011.
On the left and right side of the body of the ‘‘normal” whitefish Cor. l. pidschian, the higher number of cysts were located in the hypaxial muscles (n = 13) as compared to the epaxial muscles (n = 10). In contrast to the ‘‘normal” whitefish Cor. l. pidschian, the higher number of cysts on both sides of the body of “dwarf” whitefish Cor. l. pravdinellus were located in the epaxial muscles (n = 88) in comparison with hypaxial muscles (n = 46) and cysts located on the lateral line (n = 11) (Table 4).
Table 4.
Localization and percent distribution of T. crassus plerocercoids in muscles of forms/species of whitefishes Coregonus lavaretus.
The distribution percentage of the number of cysts in different parts of the body muscles is shown in Figure 8. On the left and right side of the body of both forms/species of whitefishes Coregonus lavaretus, the highest percentage values for distribution of the number of cysts were found on the anterior (from head to dorsal fin) and middle (between dorsal fin and adipose fin) part of the fish body. Distribution of cysts in the posterior (tail muscles) of the body were very rare (“dwarf” whitefish Cor. l. pravdinellus) or absent (‘‘normal” whitefish Cor. l. pidschian).
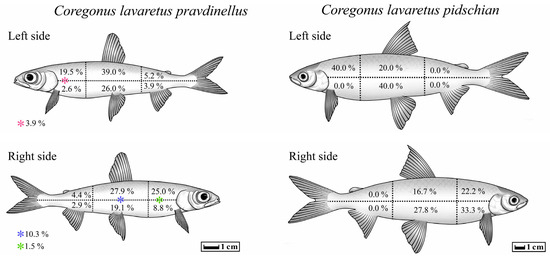
Figure 8.
Percentage distribution of T. crassus plerocercoids in different parts of the body of different forms/species of whitefishes.
4. Discussion
The first data associated with diversity of fish parasites in Teletskoye Lake were reported by Titova in 1954 [8]. There were twelve fish hosts for T. nodulosus (Th. arcticus, Brachymystax lenok, E. lucius, L. lota, Leuciscus leuciscus, Pe. fluviatilis, Cot. sibiricus, Cot. poecilopus, Hucho taimen, Cor. lavaretus sp., Barbatula toni, and Ph. phoxinus) identified and three host fish for T. crassus (E. lucius, L. lota, and Cor. lavaretus sp.) as intermediate and definitive hosts. A relatively high level of prevalence of T. nodulosus in the liver was observed for lenok B. lenok (85.8%) as the second intermediate host. In the present study, we analyzed the prevalence in liver of the same fish species with the exception of B. lenok and taimen H. taimen due to these species now being listed in the local Red Book and in need of special conservation attention [16]. In addition, we were not able to analyze the infestation level in Cot. poecilopus and B. toni due to the absence of these fish species in sites where we set the gill nets. Moreover, according to our results, the common minnow Ph. phoxinus was uninfected by T. nodulosus, while in the first report [8], a relative high level of prevalence of infestation (up to 66.0%) was found. The contradictions between our results and findings observed early are explained by differences in sites for collection of fish samples. We collected Ph. phoxinus from a small river where these fish do not consume zooplankton from Teletskoye Lake, whereas other research investigated Ph. phoxinus inhabiting a part of the lake near Yaylyu village (Figure 1b) where planktonic crustaceans were probably much more infescted by T. nodulosus.
Based on infestation levels observed, the highest prevalence of T. nodulosus infestation in Teletskoye Lake was found for burbot L. lota (100%) and is in good agreement with data (93.0%) reported in 1954 [8]. L. lota has been reported as a common host for T. nodulosus from a wide range of water bodies: Bothnian Bay, Finland (prevalence ranged from 36 to 92%) [17,18]; Baikal Lake, Russia (prevalence 26.7%) [19]; Tsipo-Tsipikan and Kuanda-Chara lakes, Russia (prevalence ranged from 60 to 100%) [20]; etc. We also found T. nodulosus in intestines of L. lota, but these worms apparently were not able to attach to the intestinal mucosa of L. lota and, consequently, were eliminated from the intestine. Other researchers found similar results when immature T. nodulosus were observed in L. lota intestine [6,17]. Moreover, relatively large L. lota do not feed on zooplankton crustaceans, which are the first intermediate hosts for both Triaenophorus species, but instead actively feed on strongly planktivorous “dwarf” whitefish–Cor. l. pravdinellus. We assume that L. lota can be defined as a paratenic host and become infected by Triaenophorus larvae when they consume fish whose stomachs are full of infected crustaceous zooplankton (Figure 9). A possible route for the reinvasion of large burbots L. lota by T. nodulosus plerocercoids is by their penetration from the intestine into the liver with subsequent re-encapsulation, an idea that was also expressed previously [21]. The reinvasion ability of plerocercoids of Triaenophorus spp., described as a characteristic feature of this genus, was reported early [22]. From an evolutionary point of view, T. nodulosus that infect the liver of L. lota never complete their lifecycle due to the adult L. lota being one of the largest fish in the lake that cannot be commonly consumed by other carnivorous fish like E. lucius, H. taimen, and B. lenok. Moreover, the illegal fisheries in the lake have eliminated large and valuable individuals of H. taimen and B. lenok, further reducing pressure on the burbot’s population. At the same time, the young individuals of L. lota could be a common prey for other carnivorous fish like E. lucius in the lake as was shown for water bodies in northern Finland [17].
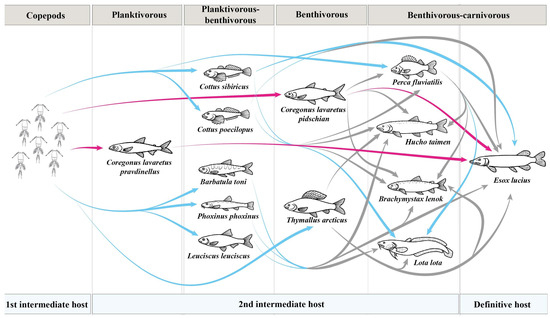
Figure 9.
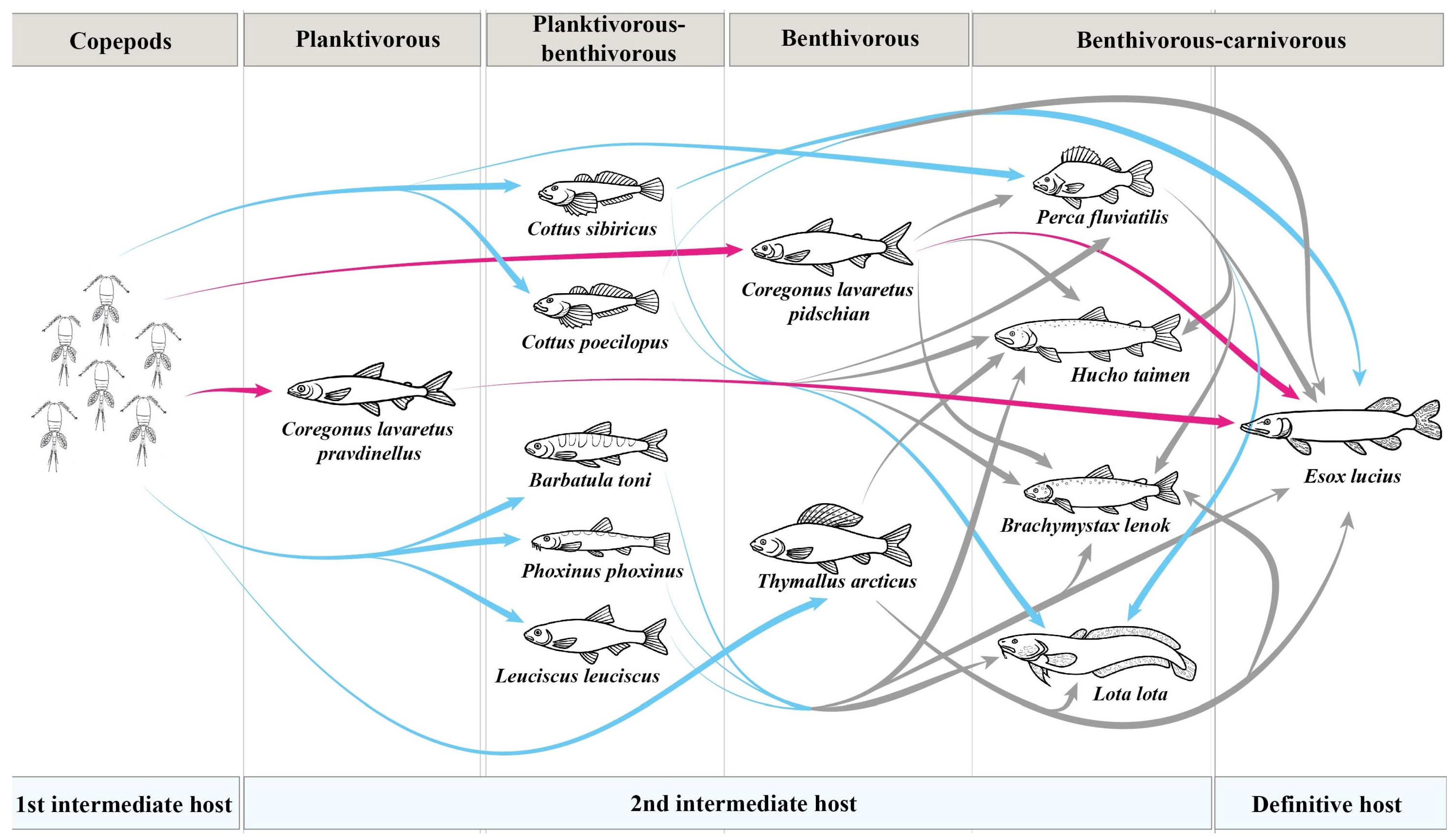
Transmission of Triaenophorus crassus and T. nodulosus through food webs in Teletskoye Lake. Pink arrow shows the transmission of T. crassus, while blue arrow shows the transmission of T. nodulosus according to our data and data obtained by Titova, 1954 [8]. Grey arrow indicate a hypothetical rout of infestation.
Among intermediate hosts, the highest prevalence of T. nodulosus infestation in Teletskoye Lake was found in the Siberian sculpin Cot. sibiricus (prevalence was up to 81.8%). The most common food items of Cot. sibiricus in Teletskoye Lake were filamentous algae, larvae of chironomids and Trichoptera, gammarids, molluscs, detritus, and zooplankton. Hence, Cot. sibiricus could be directly infected by T. nodulosus from the first intermediate host (copepods). At the same time, the ratio of this fish in the E. lucius diet was relatively low during the time of sampling (end of August–September, October). Perhaps, Cot. sibiricus is an important part of the E. lucius diet during other seasons of the year (spring, summer, or winter), but this issue was not the focus of the present study (Figure 9).
Compared to other second intermediated hosts in Teletskoye Lake, the lower values of prevalence, based on infestation levels of T. nodulosus, were shown for perch Pe. fluviatilis (24.5%) and arctic grayling Th. arcticus (31.9%). The prevalence of T. nodulosus in the liver of Pe. fluviatilis and Th. arcticus were lower almost by half when compared to results of Titova (1954) (65.0% and 76.0%, respectively). Unfortunately, Titova did not describe the size or age groups for Th. arcticus and Pe. fluviatilis that had been studied, nor the month (it was only noted that it was “summer”) when fish were collected; thus, we cannot explain the differences between hers and our results [8]. Pe. fluviatilis is one of the most frequently reported second intermediate hosts for T. nodulosus occurring in a wide range of water bodies. Previous studies found that the prevalence level of T. nodulosus infestation in Pe. fluviatilis from other lakes and rivers was much higher (46.7, 69.9, and 2.9–100%, respectively) than observed in our data [19,23,24]. To a lesser extent, Th. arcticus is also mentioned as a second intermediate host. The prevalence of T. nodulosus in Th. arcticus from Teletskoye Lake was higher (31.9%) than those found in a subspecies of arctic grayling Th. a. mertensii (12%) from the Penzhina river (Kamchatsky Krai, Russia) [25]. At the same time, the prevalence level of T. nodulosus in T. a. mertensi from the Gizhiga River basin (Russia) was 30%, which is in agreement with the level of prevalence found in our study [26]. According to feeding habits, Pe. fluviatilis from Teletskoye Lake is a facultative carnivorous fish and generally consumes larvae of chironomids, gammarids, and fish fry, whereas Th. arcticus is a benthivorous fish, whose diet includes filamentous algae, larvae of chironomids and Trichoptera, gammarids, mollusks, and stink bugs from the Pentatomidae family. Infestation of these fish occurs as a result of accidental ingestion of copepods in the case of Th. arcticus, or as a result of eating fish fry in the case of Pe. fluviatilis, in the stomachs of which zooplankton are present in large numbers (Figure 9).
In Teletskoye Lake, the main second intermediate host for T. crassus are sympatric forms/species of whitefishes (“normal” Cor. l. pidschian and “dwarf” Cor. l. pravdinellus). Unfortunately, Titova [8] did not differentiate these fishes as different forms/species and named all of them as “Teletskoye whitefish”. Titova [8] found that whitefish were more heavily infected by plerocercoids of T. crassus at a young age. As shown later, the adult specimens of these whitefishes have many morphological differences [27], but the young ones could be very similar in terms of morphology. Another study demonstrated a significantly higher level of T. crassus infestation for “dwarf” Cor. l. pravdinellus if compared to “normal” Cor. l. pidschian in Teletskoye Lake [28]. Further, Kashinskaya et al. [9] confirmed the findings previously obtained [28]. In the present study, we also found significant differences between infestation levels for forms/species of whitefishes. According to published data [9], the levels of infestation obtained for ‘‘normal” whitefish Cor. l. pidschian in the years 2019–2020 were significantly different in comparison with the levels of infestation obtained in 2022 (present study). Thus, the mean intensity and abundance of T. crassus infecting the muscles of the ‘‘normal” form/species Cor. l. pidschian in 2021–2022 was significantly lower in comparison with previous years [9]. While for the “dwarf” form/species Cor. l. pravdinellus, based on collected results over five years (2017; 2019–2022), the infestation levels of T. crassus were shown to be relative stable. These differences in levels of infestation between two forms/species of whitefishes were apparently related to feeding habits of these fishes. Indeed, the “dwarf” form/species Cor. l. pravdinellus only feeds on zooplankton crustaceans, whereas the diet of the “normal” form/species Cor. l. pidschian consists of mollusks, gammarids, detritus, larvae of different aquatic insects, and zooplankton crustaceans that play an insignificant role and perhaps significantly vary from season to season and year to year (Figure 9).
One of the main aims of the present study was also to estimate the interannual variability of infestation levels of the two species of Triaenophorus across the range of common second intermediate and definitive hosts in Teletskoye Lake. Based on results of five years of studies (2017; 2019–2022), we found statistically insignificant interannual variation in the prevalence of T. nodulosus in their second intermediate hosts. Our results were also confirmed by a previous study from water bodies in northern Finland, where stability in the infestation level of T. nodulosus in different second intermediate hosts was also noted [17]. At the same time, the interannual variation in the prevalence of T. crassus in the “normal” form of whitefish Cor. l. pidschian was significantly different. For the definitive host (E. lucius), the interannual levels of prevalence, intensity, and abundance for T. crassus have shown insignificant variations, whereas for T. nodulosus, the values were more stable. We assume that the stability in infection levels found in pike of both Triaenophorus species is determined by the relative stability of infection levels of their main fish prey (Cot. sibiricus, Pe. fluviatilis, and whitefishes, as the second intermediate hosts for T. nodulosus and T. crassus, respectively).
Moreover, with increasing length and weight of pike, the proportion of T. crassus in relation to T. nodulosus was also increased and indicated the higher importance of whitefishes in the diet of E. lucius with growth. The increased proportion of T. crassus in relation to T. nodulosus was also reported by other researchers [17,29] from Lesser Slave Lake in Canada and Bothnian Bay in northern Finland.
In order to study ecological differences among infested fish, we estimated the total, and average weight of the Triaenophorus species themselves as well as in relation to the weight of infected organ and body weight of fish. We found that the highest individual weight of cysts and highest ratio of T. nodulosus weight to weight of infected organ (liver) was found for Cot. sibiricus (0.024 ± 0.003 g and 22.8 ± 4.6%, respectively), whereas the minimum value was for Pe. fluviatilis (0.014 ± 0.005 g and 1.9 ± 0.1%, respectively). According to Kuperman [6], the ratio of T. nodulosus weight from liver of perch fry ranged from 5.0 to 60.0%. These differences could be related to different immune status of various fish species as well as different levels of infestion. Indeed, the level of T. nodulosus intensity in the liver of Pe. fluviatilis from Rybinsk Reservoir was much higher (51.8%) when compared to Pe. fluviatilis from Teletskoye Lake (26.1% in 2022).
Moreover, we found a strong positive correlation between the number of worms and their total weight for the majority of studied fish species (Cot. sibiricus, L. lota, Pe. fluviatilis, both forms/species of whitefishes). A positive significant correlation was also found between the average individual weight of cyst (in case of L. lota, Pe. fluviatilis) and their number. In another study in lakes on Vancouver Island, a positive correlation between the mean size of Schistocephalus solidus Muller, 1776 and the parasite’s abundance in threespine stickleback Gasterosteus aculeatus Linnaeus, 1758 was also found [30]. It should be considered that the weight of parasites is strongly dependent on their lifespan. Indeed, in cases of Triaenophorus species, the cysts in the second intermediate hosts (fishes) could be “young” (from the present year) and “old” (from the previous year), which is reflected in their weight. It can also be related to, for example, different sampling times and characteristics of infestation (how long the fish could be infested by parasites during a year or season) and immune and/or physiological status of the fish. All these parameters are difficult to estimate in nature, hence a similar study but under laboratory conditions is needed where one of the most important parameters—time of infestation—will be controlled.
A typical localization of T. crassus plerocercoids is intramuscular, but they may occasionally occur in other organs such as the stomach and pyloric caeca [29]. According to our data, in five cases out of 396 studied whitefishes (2019–2022), the capsules with plerocercoids were located on the liver and the surface of the stomach. In addition, we found that a higher number of cysts in muscles of the ‘‘normal” whitefish Cor. l. pidschian was found on the right side of the body when compared to the left one. Similar results were shown for ciscoes Coregonus artedi Lesueur, 1818, Cor. zenithicus Jordan and Evermann, 1909, Cor. nipigon Koelz, 1925, Cor. nigripinnis Miller, 1874 from Lesser Slave Lake and Square Lake, Alberta [31]. The author suggested that the greater number of cysts on the right side of the body of coregonids might be explained by the more right-handed location of the stomach and intestine. In a study of asymmetry in the occurrence of eye flukes in Pe. fluviatilis Linnaeus, 1758 and Rutilus rutilus Linnaeus, 1758 the asymmetrical locations were also seasonal and possibly influenced by the amount of rainfall (likely influencing prey abundance/availability) [32]. The authors attributed the altered left–right distribution to variations in the vasculature of the eye, which is not in disagreement with the idea posited herein regarding the positioning of the stomach and the asymmetry of the helminth infections. Further, Newton [31] studied the distribution of T. crassus plerocercoids in whitefish from the same lakes. He found that 80 percent of the cysts were in the epaxial muscles between the head and the dorsal fin. Kuperman [6] also confirmed these data and showed that 82.6% of T. crassus plerocercoids infecting C. albula Linnaeus, 1758 from Lake Ladoga and Rybinsk Reservoir were in the dorsal muscles, 4.4% on the abdominal wall muscles and 13.0% in the area of anal fin. Similar to Newton [31], we also noted that T. crassus plerocercoids in the tail musculature were rare or absent in the case of “dwarf” and ‘‘normal” forms, respectively. This effect can potentially be explained by the fact that penetration of the parasite to tail musculature is restricted by the length of the fish intestine and the parasites’ movement. Indeed, Rosen and Dick [33] also showed that the penetration of parasites more regularly occur through the stomach, pyloric caeca, or anterior intestine.
5. Conclusions
Our study revealed the range of the hosts and levels of infestation of two different Triaenophorus species (T. nodulosus and T. crassus) from a deep, oligotrophic mountain lake in Siberia. Due to the relatively wide range of hosts, we conclude that both Triaenophorus species are well integrated into the aquatic system of the lake. Based on the infestation level, the most important hosts in the lifecycle of T. crassus and T. nodulosus in Teletskoye Lake are Cor. l. pravdinellus and Cot. sibiricus, respectively. For these fishes, the direct route of parasite transmission to the definitive host was shown. Despite the high level of infestation, L. lota in Teletskoye Lake is an evolutionary dead-end host in the lifecycle of T. nodulosus due to it occupying the niche of the largest carnivorous fish. An indirect route of Triaenophorus spp. transmission in Teletskoye Lake has been shown for the “normal” Cor. l. pidschian, Th. arcticus, and facultative carnivorous Pe. fluviatilis. Infestation of these fishes may occur as a result of accidental ingestion of copepods or reinvasion by Triaenophorus spp. This assumption confirms our first hypothesis. Based on results from five years of sampling, we found stability of T. nodulosus in their second intermediate hosts, whereas for T. crassus infestations, they show significant interannual variation. At the same time, for the definitive host (E. lucius), the interannual levels of prevalence, intensity, and abundance for T. nodulosus were more stable, whereas for T. crassus there were only insignificant variations to confirm our second hypothesis. Differences in feeding habits and physiology of various fishes probably may affect on the mass of parasites through their abundance, supporting our third hypothesis but needs more extensive studies in the future. At the same time, the high rates of invasion by Triaenophorus plerocercoids can potentially affect the locomotor activity of fish. The lower activity of the host under parasitic invasion makes the fish more accessible to the definitive host, which, as a result, increases the success of implementation of the parasite lifecycle.
Supplementary Materials
The following supporting information can be downloaded at: https://www.mdpi.com/article/10.3390/ani13193122/s1, Figure S1: Size of cestodes (white bar) and their relationship with eviscerated body weight of fish (blue bar).
Author Contributions
Conceptualization, Methodology, Writing—original draft, E.N.K. and M.M.S. Investigation, E.N.K., P.G.V., T.V.K., G.V.I., A.V.S., G.A.R., G.N.M. and M.M.S. Writing—review and editing, E.N.K., M.M.S. and K.B.A. Visualization, E.N.K. and A.V.S. Funding acquisition, E.N.K. and M.M.S. All authors have read and agreed to the published version of the manuscript.
Funding
The research of 2020 was partially supported by the Russian Foundation for Basic Research (grant number 19-34-60028), the data curation in 2021 was partially supported by the Russian Science Foundation (project No. 19-74-10054), and for 2022 was partially supported by the Russian international scientific collaboration program Mega-grant (mega-grant No. 075-15-2022-1134).
Institutional Review Board Statement
The present research meets the requirements guided by the order of the High and Middle Education Ministry (care for vertebrate animal included in scientific experiments, #742 from 13 November 1984) and additionally by the Federal Law of the Russian Federation #498 FL (from 19 December 2018) with regard to the humane treatment of animals. The present study was based on field research and involved the collection of fishes from natural water bodies. Adult fish used in the current study were collected based on permeation number (№ 542019030188, 542020030123, 542021030077, and 542022030074) obtained from the Russian Federal Fisheries Agency (https://fish.gov.ru/territorial_adm/verkhneobskoe/, accessed on 18 August 2023). The permit allowed us to collect fish in the specified water bodies. No other permits were required to carry out the present study in designated areas under Russian Federation laws.
Informed Consent Statement
Not applicable.
Data Availability Statement
All data generated or analyzed during this study are included in this published article.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Knudsen, R.; Rikardsen, A.H.; Dempson, J.B.; BjØrn, P.A.; Finstad, B.; Holm, M.; Amundsen, P.-A. Trophically transmitted parasites in wild Atlantic salmon post-smolts from Norwegian fjords. J. Fish Biol. 2005, 66, 758–772. [Google Scholar] [CrossRef]
- Knudsen, R.; Amundsen, P.A.; Nilsen, R.; Kristoffersen, R.; Klemetsen, A. Food borne parasites as indicators of trophic segregation between Arctic charr and brown trout. Environ. Biol. Fishes 2008, 83, 107–116. [Google Scholar] [CrossRef]
- Busarova, O.; Markevich, G.; Esin, E. Trophic differentiation of the nosed charr Salvelinus schmidti Viktorovsky, 1978 in Lake Kronotskoe (Kamchatka). Russ. J. Mar. Biol. 2017, 43, 57–64. [Google Scholar] [CrossRef]
- Bonato, K.O.; Silva, P.C.; Malabarba, L.R. Unrevealing parasitic trophic interactions—A molecular approach for fluid-feeding fishes. Front. Ecol. Evol. 2018, 6, 22. [Google Scholar] [CrossRef]
- Vlasenko, P.G.; Sokolov, S.G.; Ieshko, E.P.; Frolov, E.V.; Kalmykov, A.P.; Parshukov, A.N.; Chugunova, Y.K.; Kashinskaya, E.N.; Shokurova, A.V.; Bochkarev, N.A.; et al. A re-evaluation of conflicting taxonomic structures of Eurasian Triaenophorus spp. (Cestoda, Bothriocephalidea: Triaenophoridae) based on partial cox1 mtDNA and 28S rRNA gene sequences. Can. J. Zool. 2022, 100, 323–333. [Google Scholar] [CrossRef]
- Kuperman, B.I. Tapeworms of the Genus Triaenophorus, Parasites of Fish. Experimental Systematics, Ecology; Nauka: Leningrad, Russia, 1973; p. 208. (In Russian) [Google Scholar]
- Kuchta, R.; Vlckova, R.; Poddubnaya, L.G.; Gustinelli, A.; Dzika, E.; Scholz, T. Invalidity of three Palaearctic species of Triaenophorus tapeworms (Cestoda: Pseudophyllidea): Evidence from morphometric analysis of scolex hooks. Folia Parasitol. 2007, 54, 34–42. [Google Scholar] [CrossRef] [PubMed]
- Titova, S.D. Parasites of fish from Teletskoye Lake. Tr. Probl. I Temat. Soveshhanij Zin. Vii Soveshhanie Po Parazitol. Probl. 1954, 4, 79–84. (In Russian) [Google Scholar]
- Kashinskaya, E.N.; Vlasenko, P.G.; Bochkarev, N.A.; Andree, K.B.; Solovyev, M.M. Feeding habits shape infection levels by plerocercoids of the tapeworm Triaenophorus crassus in muscle of a sympatric pair of whitefish in an oligotrophic lake. J. Helminthol. 2021, 95, e8. [Google Scholar] [CrossRef]
- Kuperman, B.I. New species of the genus Triaenophorus Rud. (Cestoda, Pseudophyllidea). Parazitologiya 1968, 2, 495–501. (In Russian) [Google Scholar]
- Bush, A.O.; Lafferty, K.D.; Lotz, J.M.; Shostak, A.W. Parasitology meets ecology on its own terms: Margolis et al. Revisited. J. Parasitol. Res. 1997, 83, 575–583. [Google Scholar] [CrossRef]
- Hammer, Ø.; Harper, D.A.T.; Ryan, P.D. PAST: Paleontological statistics software package for education and data analysis. Palaeontol. Electron. 2001, 4, 1–9. Available online: http://palaeo-electronica.org/2001_1/past/issue1_01.html (accessed on 1 April 2023).
- Wickham, H. ggplot2: Elegant Graphics for Data Analysis, 2nd ed.; Springer: New York, NY, USA, 2016; 368p. [Google Scholar] [CrossRef]
- Reiczigel, J.; Marozzi, M.; Fábián, I.; Rózsa, L. Biostatistics for parasitologists—A primer to quantitative parasitology. Trends Parasitol. 2019, 35, 277–281. [Google Scholar] [CrossRef]
- Wayland, M.T.; Chubb, J.C. A new R package and web application for detecting bilateral asymmetry in parasitic infections. Folia Parasitol. 2016, 63, 1–5. [Google Scholar] [CrossRef] [PubMed]
- Bondarenko, A.V. (Ed.) Red Book of the Altai Republic. Animals, 3rd ed.; GAGU: Gorno-Altaisk, Russia, 2017; 368p. (In Russian) [Google Scholar]
- Valtonen, E.T.; Rintamäki, P.; Lappalainen, M. Triaenophorus nodulosus and T. crassus in fish from Northern Finland. Folia Parasitol. 1989, 36, 351–370. [Google Scholar]
- Andersen, K.I.; Valtonen, E.T. Segregation and co-occurrence of larval cestodes in freshwater fishes in the Bothnian Bay, Finland. Parasitology 1992, 104, 161–168. [Google Scholar] [CrossRef]
- Rusinek, O.T. Fish Parasites of Lake Baikal (Fauna, Communities, Zoogeography and Historical Background); KMK Scientific Press Ltd.: Moscow, Russia, 2007; p. 571. (In Russian) [Google Scholar]
- Dugarov, Z.N.; Pronin, N.M.; Batueva, M.D.; Burdukovskaya, T.G.; Matveev, A.N.; Pronina, S.V.; Sondueva, L.D. Fauna of parasites of burbot Lota lota (L.) (Gadiformes: Lotidae) from reservoirs of Baikal Rift Zone. Inland Water Biol. 2016, 9, 341–349. [Google Scholar] [CrossRef]
- Bauer, O.N. Ecology of parasites of freshwater fish (Relationship of the parasite with environment). Izv. GosNIORH 1959, 49, 5–206. (In Russian) [Google Scholar]
- Protasova, E.N. Cestodes of Fish—Bothriocephalata. Principles of Cestodology; Ryzhikov, K.M., Ed.; Nauka: Moscow, Russia, 1977; Volume 8, 298p. (In Russian) [Google Scholar]
- Masson, G.; Vanacker, M.; Fox, M.G.; Beisel, J.N. Impact of the cestode Triaenophorus nodulosus on the exotic Lepomis gibbosus and the autochthonous Perca fluviatilis. Parasitology 2015, 142, 745–755. [Google Scholar] [CrossRef]
- Morley, N.J.; Lewis, J.W. Influence of Triaenophorus nodulosus plerocercoids (Cestoda: Pseudophyllidea) on the occurrence of intestinal helminths in the perch (Perca fluviatilis). J. Helminthol. 2017, 91, 711–717. [Google Scholar] [CrossRef]
- Busarova, O.; Koval, M.; Esin, E.; Markevich, G. Trophic segregation in multispecies community of salmonids in the Penzhina river lower course (Kamchatsky krai, Russia). Nat. Conserv. Res. 2019, 4, 83–94. [Google Scholar] [CrossRef]
- Pospekhov, V.V.; Atrashkevich, G.I.; Orlovskaya, O.M. Parasites of graylings (Thymallidae: Thymallus) from the northern continental coast of the Okhotsk Sea. Izv. TINRO 2020, 200, 965–977. [Google Scholar] [CrossRef]
- Bochkarev, N.A.; Zuikova, E.I. Morphological, biological, and ecological differentiation of sympatric Coregonus species from Teletskoe Lake. Russ. J. Zool. 2006, 85, 950–958. (In Russian) [Google Scholar]
- Bochkarev, N.A.; Gafina, T.E. Comparative characteristics of whitefish from Teletskoye Lake (Altai Republic). Sib. Biol. J. 1993, 2, 64–69. (In Russian) [Google Scholar]
- Miller, R.B. A Review of the Triaenophorus Problem in Canadian Lakes, 1st ed.; Fisheries Research Board of Canada: Ottawa, ON, Canada, 1952; Bulletin 95; 42p. [Google Scholar]
- Weber, J.N.; Steinel, N.C.; Shim, K.C.; Bolnick, D.I. Recent evolution of extreme cestode growth suppression by a vertebrate host. Proc. Natl. Acad. Sci. USA 2022, 114, 6575–6580. [Google Scholar] [CrossRef] [PubMed]
- Newton, M.V.B. The biology of Triaenophorus tricuspidatus, (Bloch 1779), in western Canada. Contrib. Can. Biol. Fish 1932, 7, 341–360. [Google Scholar] [CrossRef]
- Morley, N.J.; Lewis, J.W. Influence of an extreme climatic event on bilateral asymmetry and occurrence of eye flukes in perch (Perca fluviatilis) and roach (Rutilus rutilus) from a lake in southeast England. J. Helminthol. 2020, 94, e57. [Google Scholar] [CrossRef]
- Rosen, R.; Dick, T.A. Development and infectivity of the procercoid of Triaenophorus crassus Forel and mortality of the first intermediate host. Can. J. Zool. 1983, 61, 2120–2128. [Google Scholar] [CrossRef]
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).