
The Food Niche Overlap and Interspecific Relationship between the Sympatric Tibetan Macaque and Grey Snub-Nosed Monkey
 and
and
Abstract
Simple Summary
Abstract
1. Introduction
2. Materials and Methods
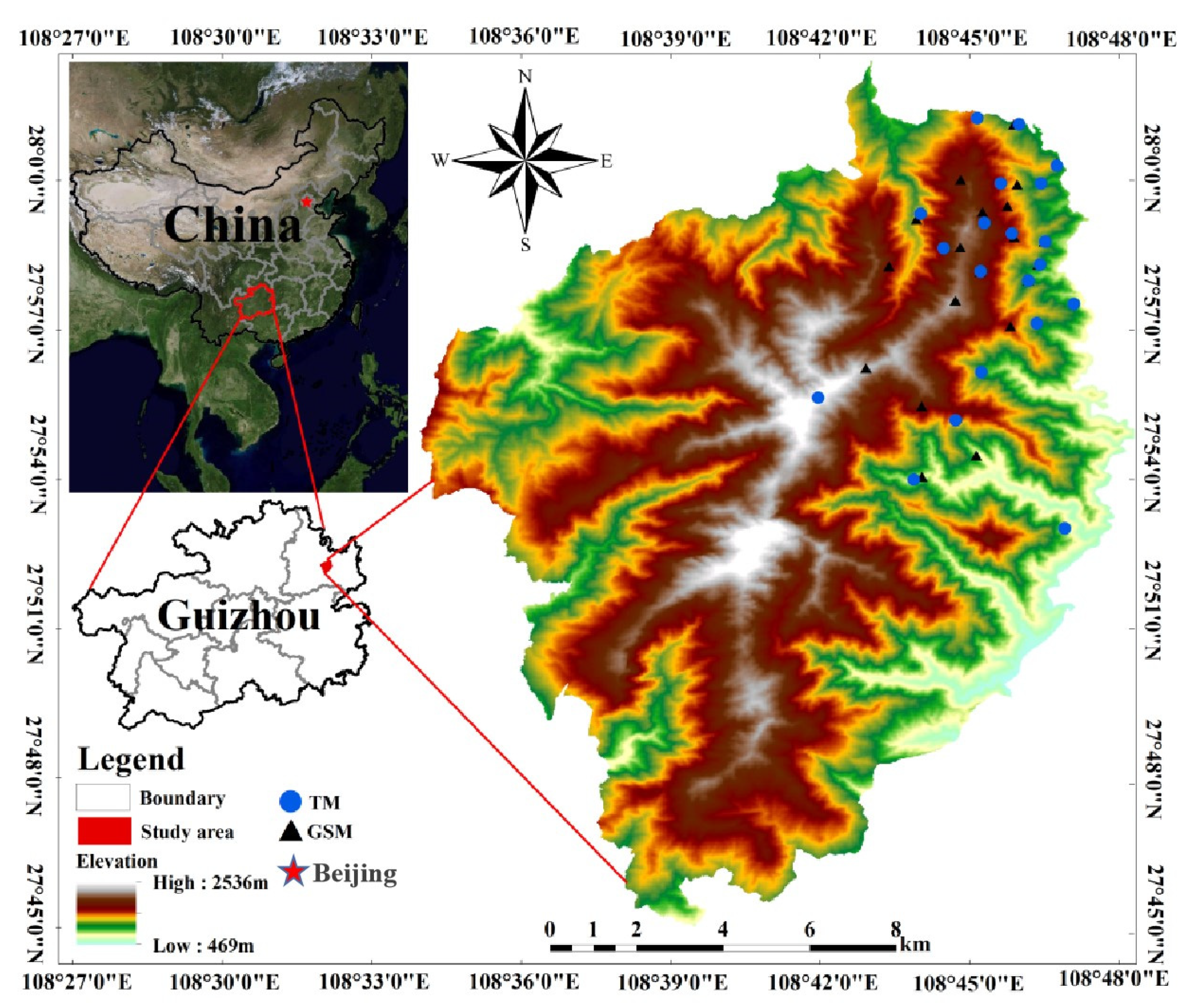
2.1. Study Area and Fecal Collection
2.2. DNA Extraction
2.3. PCR Amplification for Fecal Species Identification
2.4. Amplicon Sequencing
2.5. Food Niche Overlap Analysis
2.6. Statistical Analysis
3. Results
3.1. Food Composition of Tibetan Macaques and Grey Snub-Nosed Monkeys
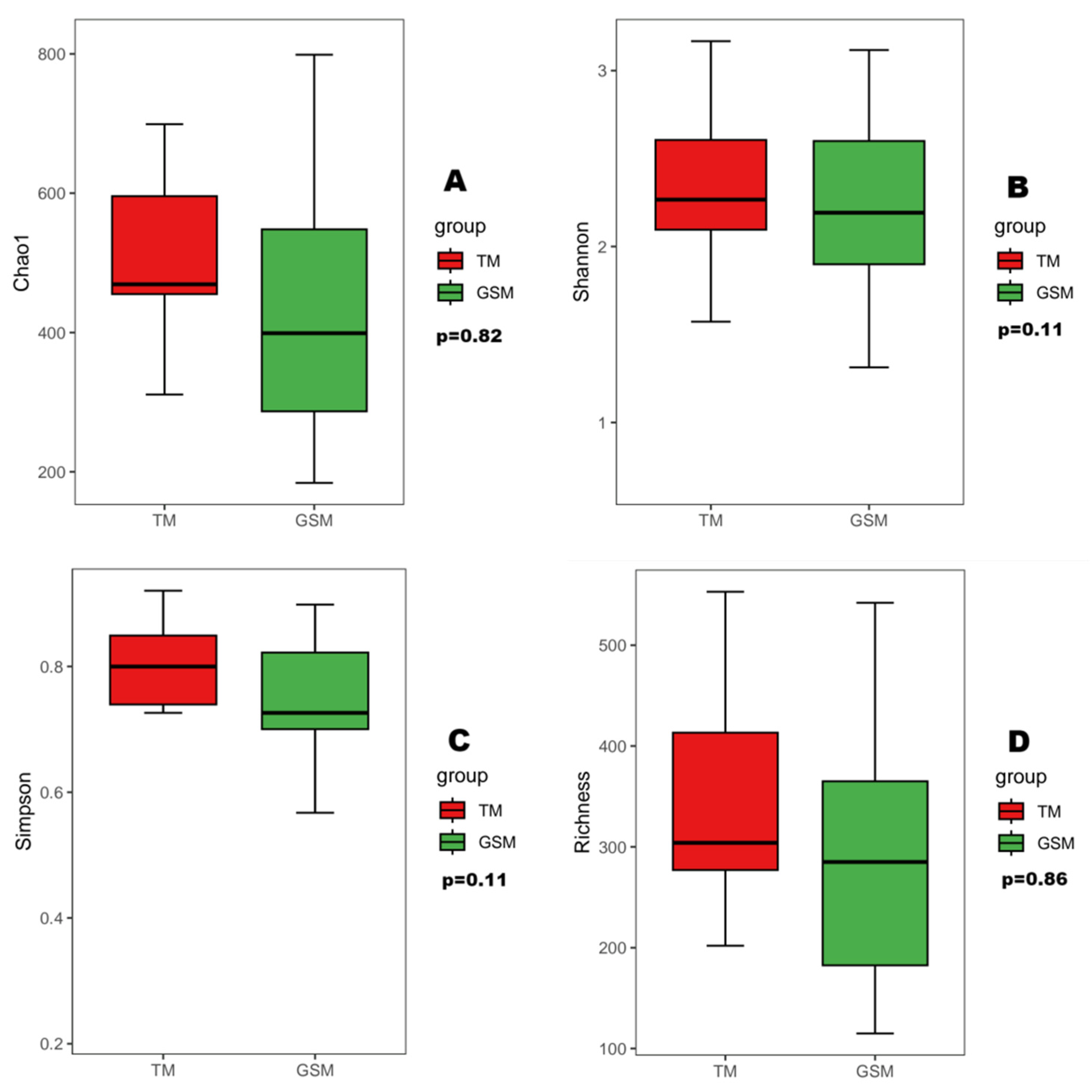
3.2. Analysis of Food Diversity in Tibetan Macaques and Grey Snub-Nosed Monkeys
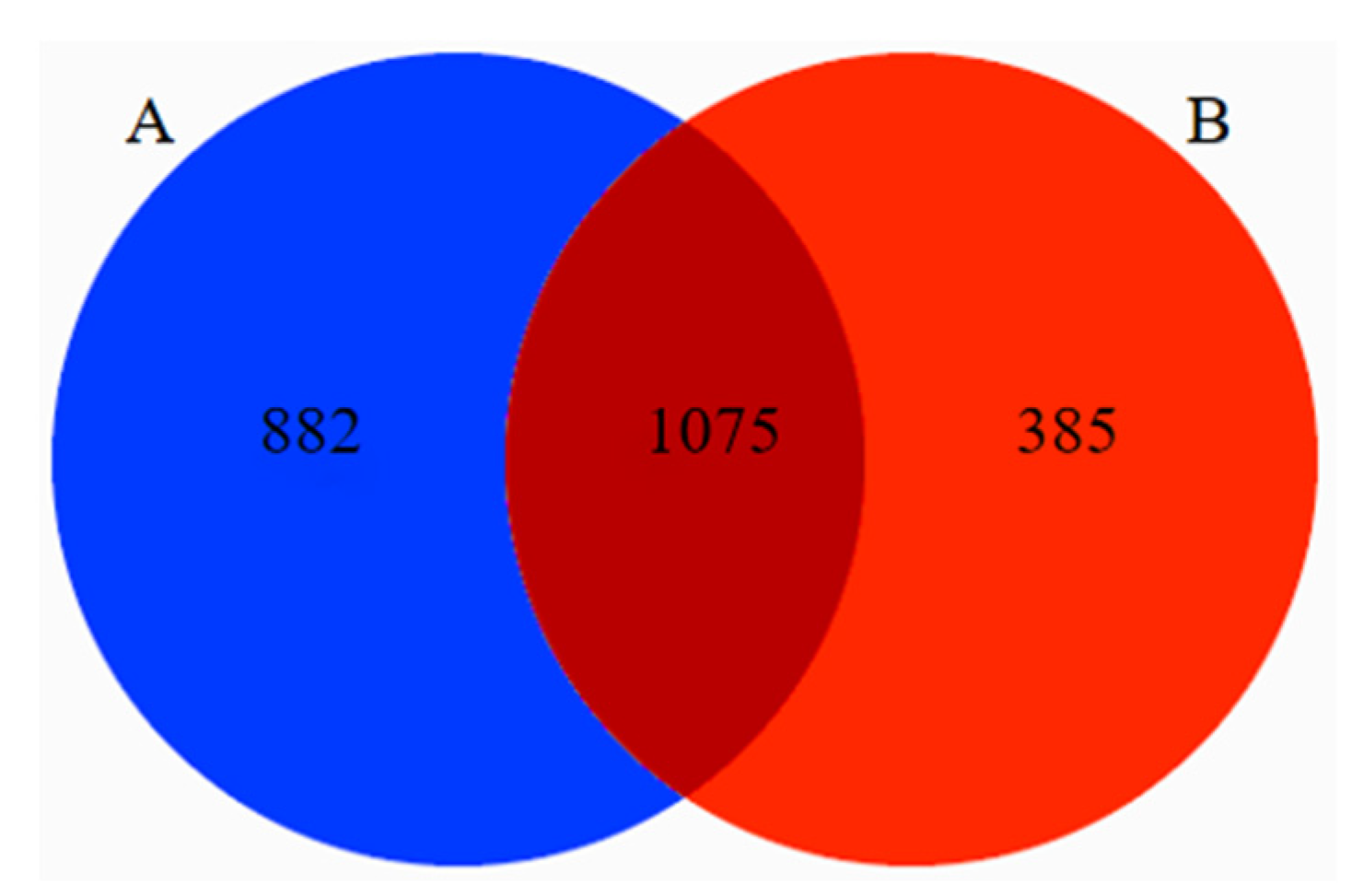
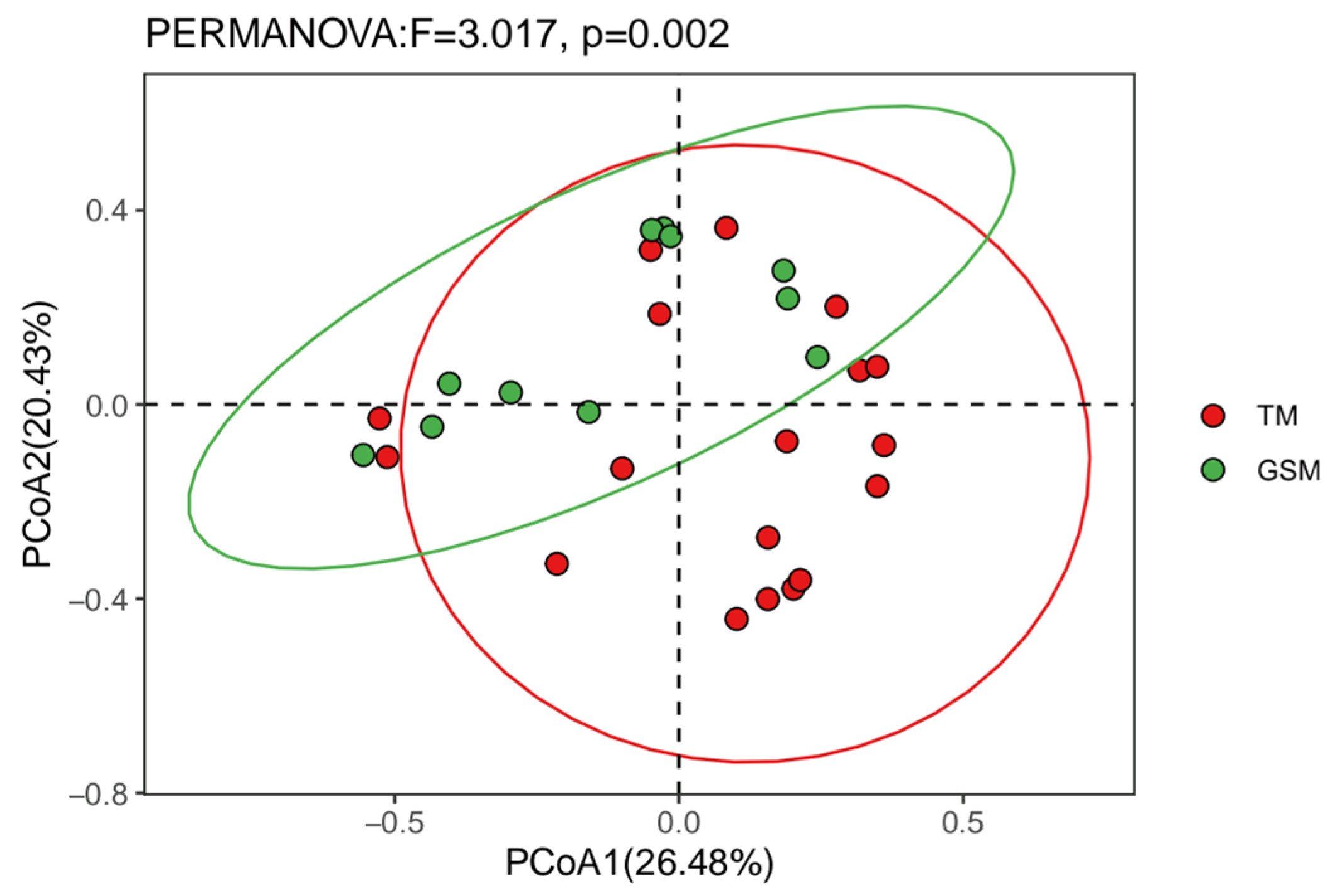
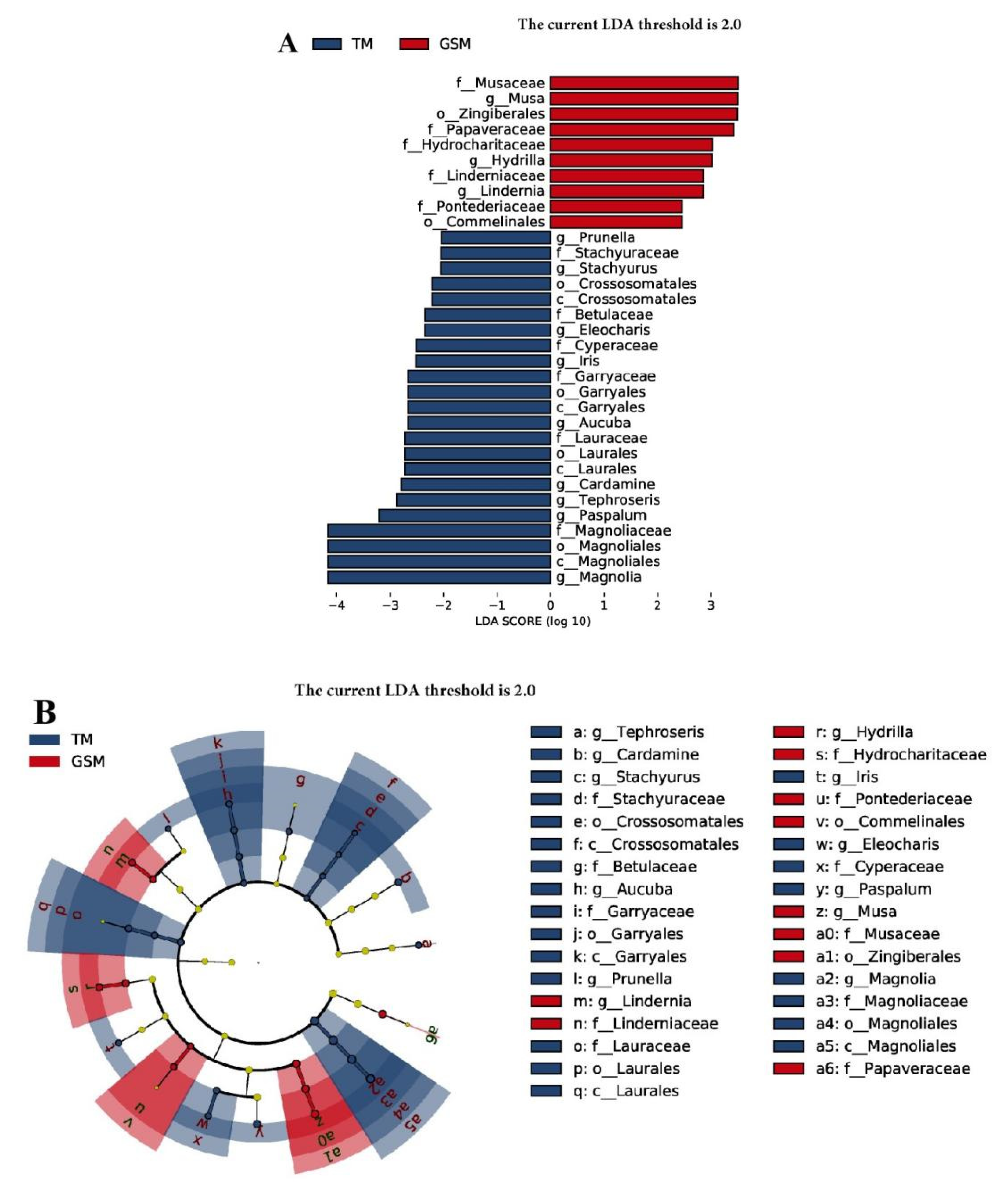
3.3. Difference Analysis of Food in Tibetan Macaques and Grey Snub-Nosed Monkeys
3.4. Food Niche Overlap of Tibetan Macaques and Grey Snub-Nosed Monkeys
4. Discussion
4.1. Diet of the Two Primates
4.2. Food Diversity in the Two Primates
4.3. Coexistence Mechanism of the Two Primates
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Wang, G.L.; Yin, H.B. Advances on foraging behavioral ecology in nonhuman primates. J. Biol. 2008, 5, 10–12. [Google Scholar]
- Hutchinson, G.E. Concluding Remarks. Cold Spring Harb. Lab. Press 1957, 22, 415–427. [Google Scholar] [CrossRef]
- Chillo, V.; Rodríguez, D.; Ojeda, R.A. Niche partitioning and coexistence between two mammalian herbivores in the Dry Chaco of Argentina. Acta Oecologica 2010, 36, 611–616. [Google Scholar] [CrossRef]
- Hardin, G. The Competitive Exclusion Principle. Science 1960, 131, 1292–1297. [Google Scholar] [CrossRef]
- Jonathan, M.; Levine, C.M.D. Elton revisited: A review of evidence linking diversity and invasibility. Oikos 1999, 87, 15–26. [Google Scholar] [CrossRef]
- Chu, C.J.; Wang, Y.S.; Liu, Y.; Jiang, L.; He, F.L. Advances in species coexistence theory. Biodivers. Sci. 2017, 25, 345–354. [Google Scholar] [CrossRef]
- Eloranta, A.P.; Finstad, A.G.; Sandlund, O.T.; Knudsen, R.; Kuparinen, A.; Amundsen, P.A. Species interactions, environmental gradients and body size shape population niche width. J. Anim. Ecol. 2022, 91, 154–169. [Google Scholar] [CrossRef]
- Lanszki, Z.; Horváth, G.F.; Bende, Z.; Lanszki, J. Differences in the diet and trophic niche of three sympatric carnivores in a marshland. Mammal Res. 2020, 65, 93–104. [Google Scholar] [CrossRef]
- Namgail, T.; Mishra, C.; de Jong, C.B.; van Wieren, S.E.; Prins, H.H.T. Effects of herbivore species richness on the niche dynamics and distribution of blue sheep in the Trans-Himalaya. Divers. Distrib. 2009, 15, 940–947. [Google Scholar] [CrossRef]
- Hadi, S.; Ziegler, T.; Waltert, M.; Syamsuri, F.; Mühlenberg, M.; Hodges, J.K. Habitat Use and Trophic Niche Overlap of Two Sympatric Colobines, Presbytis potenziani and Simias concolor, on Siberut Island, Indonesia. Int. J. Primatol. 2012, 33, 218–232. [Google Scholar] [CrossRef]
- Zhou, Q.; Huang, Z.; Wei, H.; Huang, C.; Zhou, Q.; Huang, Z.; Wei, H.; Huang, C. Variations in diet composition of sympatric Trachypithecus francoisi and Macaca assamensis in the limestone habitats of Nonggang, China. Zool. Res. 2018, 39, 284–290. [Google Scholar] [CrossRef] [PubMed]
- Liu, C.; Li, W.B.; Li, B.W.; Yang, P.P.; Xia, D.P.; Li, J.H. Food composition and nutrients of wild Tibetan macaques (Macaca thibetana) in winter. Acta Theriol. Sin. 2021, 41, 649–657. [Google Scholar]
- Ribeiro, L.B.; Freire, E.M.X. Trophic ecology and foraging behavior of Tropidurus hispidus and Tropidurus semitaeniatus (Squamata, Tropiduridae) in a caatinga area of northeastern Brazil. Série Zool. 2011, 101, 225–232. [Google Scholar] [CrossRef]
- Juri, G.L.; Naretto, S.; Mateos, A.C.; Chiaraviglio, M.; Cardozo, G. Influence of Life History Traits on Trophic Niche Segregation between Two Similar Sympatric Tupinambis Lizards. S. Am. J. Herpetol. 2015, 10, 132–142. [Google Scholar] [CrossRef]
- Pompanon, F.; Deagle, B.E.; Symondson, W.O.C.; Brown, D.S.; Jarman, S.N.; Taberlet, P. Who is eating what: Diet assessment using next generation sequencing. Mol. Ecol. 2012, 21, 1931–1950. [Google Scholar] [CrossRef] [PubMed]
- Guo, Y.P.; Zhang, H.; Zhao, X.G.; Luo, H.L.; Zhang, Y.J. Applications of DNA Metabarcoding in Diet Identification of Herbivores. Biotechnol. Bull. 2021, 37, 252–260. [Google Scholar] [CrossRef]
- Liu, G.; Ning, Y.; Xia, X.F.; Gong, M.H. The application of high-throughput sequencing technologies to wildlife diet analysis. Acta Ecol. Sin. 2018, 38, 3347–3356. [Google Scholar]
- Tillmar, A.O.; Dell’Amico, B.; Welander, J.; Holmlund, G. A Universal Method for Species Identification of Mammals Utilizing Next Generation Sequencing for the Analysis of DNA Mixtures. PLoS ONE 2013, 8, e83761. [Google Scholar] [CrossRef]
- Verkuil, Y.I.; Nicolaus, M.; Ubels, R.; Dietz, M.W.; Samplonius, J.M.; Galema, A.; Kiekebos, K.; de Knijff, P.; Both, C. DNA metabarcoding quantifies the relative biomass of arthropod taxa in songbird diets: Validation with camera-recorded diets. Ecol. Evol. 2022, 5, 12. [Google Scholar]
- Guillerault, N.; Bouletreau, S.; Iribar, A.; Valentini, A.; Santoul, F. Application of DNA metabarcoding on faeces to identify European catfishSilurus glanis diet. J. Fish Biol. 2017, 90, 2214–2219. [Google Scholar] [CrossRef]
- Brun, L.; Schneider, J.; Carrió, E.M.; Dongre, P.; Taberlet, P.; Waal, V.D.; Fumagalli, L. Focal vs. fecal: Seasonal variation in the diet of wild vervet monkeys from observational and DNA metabarcoding data. Ecol. Evol. 2022, 10, 12. [Google Scholar] [CrossRef] [PubMed]
- Pereira, A.; Samlali, M.A.; Khifa, A.S.; Slimani, T.; Harris, D.J. A pilot study on the use of DNA metabarcoding for diet analysis in a montane amphibian population from North Africa. Afr. J. Herpetol. 2021, 70, 68–74. [Google Scholar] [CrossRef]
- Norconk, M.A.; Boinski, S.; Forget, P. Primates in 21st century ecosystems: Does primate conservation promote ecosystem conservation? Am. J. Primatol. 2011, 73, 3–8. [Google Scholar] [CrossRef]
- Osman, N.A.; Abdul-Latiff, M.A.B.; Mohd-Ridwan, A.R.; Yaakop, S.; Karuppannan, K.V.; Md-Zain, B.M.; Yaakop, K.V.K.B. Metabarcoding data analysis revealed the plant dietary variation of long-tailed macaque Macaca fascicularis (Cercopithecidae, Cercopithecinae) living in disturbed habitats in Peninsular Malaysia. Biodivers. Data J. 2022, 10, e89617. [Google Scholar] [CrossRef]
- Srivathsan, A.; Ang, A.; Vogler, A.P.; Meier, R. Fecal metagenomics for the simultaneous assessment of diet, parasites, and population genetics of an understudied primate. Front. Zool. 2016, 13, 17. [Google Scholar] [CrossRef]
- Rowe, A.K.; Donohue, M.E.; Clare, E.L.; Drinkwater, R.; Koenig, A.; Ridgway, Z.M.; Martin, L.D.; Nomenjanahary, E.S.; Zakamanana, F.; Randriamanandaza, L.J.; et al. Exploratory analysis reveals arthropod consumption in 10 lemur species using DNA metabarcoding. Am. J. Primatol. 2021, 83, e23256. [Google Scholar] [CrossRef] [PubMed]
- Machado, F.F.; Jardim, L.; Dinnage, R.; Brito, D.; Cardillo, M. Diet disparity and diversity predict extinction risk in primates. Anim. Conserv. 2023, 26, 331–339. [Google Scholar] [CrossRef]
- Xia, W.P.; Zhang, Y.Z. Primae Research and Conservation; China Forestry Publishing House: Beijing, China, 1995; pp. 150–151. [Google Scholar]
- Zhu, C.Q.; Li, D.Q.; Dong, K.; Wang, X.L.; Shi, P.L. Setting biodiversity conservation priorities in the Forests of the Upper Yangtze Ecoregion based on ecoregion conservation methodology. Biodivers. Sci. 2006, 2, 87–97. [Google Scholar]
- Zhou, X.Z.; Yang, Y.Q. Fanjingshan Research; Guizhou People’s Publishing House: Guiyang, China, 1990; pp. 3, 444–445. [Google Scholar]
- Luo, R. The Mammalian Fauna of Guizhou; Guizhou Science and Technology Publishing House: Guiyang, China, 1993; pp. 155–162. [Google Scholar]
- Yang, Y.Q.; Lei, X.P.; Yang, C.D. Fanjingshan Research; Guizhou Science and Technology Publishing House: Guiyang, China, 2002; p. 6. [Google Scholar]
- IUCN Red List of Threatened Species. Available online: https://www.iucnredlist.org/en (accessed on 12 February 2023).
- Cui, D.Y.; Niu, K.F.; Tan, C.L.; Yang, M.; Shi, L.; Zhang, W.Y.; Zhang, J.G.; Yang, Y.Q. Golden snub-nosed monkey distributed in Fanjing Mountain, Guizhou, China. Chin. J. Zool. 2014, 49, 792. [Google Scholar] [CrossRef]
- Guo, Y.Q.; Zhou, J.; Song, X.H.; Deng, H.Q.; Qiu, Y.; Shi, L.; Zhou, J. The population of Rhinopithecus brelichi in Fanjingshan National Nature Reserve, Guizhou, China. Acta Theriol. Sin. 2017, 37, 104–108. [Google Scholar] [CrossRef]
- Bleisch, W.; Cheng, A.S.; Ren, X.D.; Xie, J.H. Preliminary results from a field study of wild Guizhou snub-nosed monkeys (Rhinopithecus brelichi). Folia Primatol. 1993, 60, 72–82. [Google Scholar] [CrossRef] [PubMed]
- Bleisch, W.; Xie, J.H. Ecology and behavior of the Guizhou snub-nosed langur (Rhinopithecus brelichi). In The Natural History of the Doucs and Snub-Nosed Monkeys; World Scientific: Singapore, 1998; pp. 217–239. [Google Scholar] [CrossRef]
- Xiang, Z.; Liang, W.; Nie, S.; Li, M. Diet and Feeding Behavior of Rhinopithecus brelichi at Yangaoping, Guizhou. Am. J. Primatol. 2012, 74, 551–560. [Google Scholar] [CrossRef] [PubMed]
- Chang, Y.; Zhang, M.M.; Liu, Z.S.; Hu, T.H.; Li, Z.G. Summer diets of Sympatric Blue Sheep (Pseudois nayaur) and Red Deer (Cervus elaphus alxaicus) in the Helan Mountains, China. Acta Ecol. Sin. 2010, 30, 1486–1493. [Google Scholar]
- Zhou, Z.X.; Yang, Y.Q. Research on Fanjing Mountain; Guizhou People’s Publishing House: Guiyang, China, 1990; pp. 95–96. [Google Scholar]
- Vallet, D.; Petit, E.J.; Gatti, S.; Levréro, F.; Ménard, N. A new 2CTAB/PCI method improves DNA amplification success from faeces of Mediterranean (Barbary macaques) and tropical (lowland gorillas) primates. Conserv. Genet. 2008, 9, 677–680. [Google Scholar] [CrossRef]
- Xiong, M.Y.; Wang, D.J.; Bu, H.L.; Shao, X.N.; Zhang, D.; Li, S.; Wang, R.J.; Yao, M. Molecular dietary analysis of two sympatric felids in the Mountains of Southwest China biodiversity hotspot and conservation implications. Sci. Rep. 2017, 7, 41909. [Google Scholar] [CrossRef]
- Hofreiter, M.; Poinar, H.N.; Spaulding, W.G.; Bauer, K.; Martin, P.S.; Possnert, G.; Paabo, S. A molecular analysis of ground sloth diet through the last glaciation. Mol. Ecol. 2000, 9, 1975–1984. [Google Scholar] [CrossRef]
- Geller, J.; Meyer, C.; Parker, M.; Hawk, H. Redesign of PCR primers for mitochondrial cytochrome c oxidase subunit I for marine invertebrates and application inall-taxabioticsurveys. Wiley Online Libr. 2013, 13, 851–861. [Google Scholar] [CrossRef]
- Leray, M.; Yang, J.Y.; Meyer, C.P.; Mills, S.C.; Agudelo, N.; Ranwez, V.; Boehm, J.T.; Machida, R.J. A new versatile primer set targeting a short fragment of the mitochondrial COI region for metabarcoding metazoan diversity: Application for characterizing coral reef fish gut contents. Front. Zool. 2013, 10, 34. [Google Scholar] [CrossRef]
- Edgar, R.C. UPARSE: Highly accurate OTU sequences from microbial amplicon reads. Nat. Methods 2013, 10, 996–998. [Google Scholar] [CrossRef]
- Shao, X.N.; Song, D.Z.; Huang, Q.W.; Li, S.; Yao, M. Fast surveys and molecular diet analysis of carnivores based on fecal DNA and Fast surveys and molecular diet analysis of carnivores based on fecal DNA and metabarcoding. Biodivers. Sci. 2019, 27, 543–556. [Google Scholar]
- Cao, H.; Yang, X.; Peng, C.; Wang, Y.; Guo, Q.; Su, H.J. Winter Dietary Analysis Reveals the Foraging Differences of Wild Boar (Sus scrofa) in Different Regions of a Karst Mountainous Area. Animals 2023, 13, 727. [Google Scholar] [CrossRef] [PubMed]
- Deagle, B.E.; Thomas, A.C.; McInnes, J.C.; Clarke, L.J.; Vesterinen, E.J.; Clare, E.L.; Kartzinel, T.R.; Eveson, J.P. Counting with DNA in metabarcoding studies: How should we convert sequence reads to dietary data? Mol. Ecol. 2019, 28, 391–406. [Google Scholar] [CrossRef] [PubMed]
- Cui, D.Y.; Liu, Z.S.; Xiao, M.; Zhai, H.; Hu, T.H.; Li, Z.G. Winter Food-habits of Red Deer (Cervus elaphus alxaicus) in Helan Mountains, China. Zool. Res. 2007, 04, 383–388. [Google Scholar]
- Nie, S.G.; Xiang, Z.F.; Li, M. Preliminary report on the diet and social structure of gray snub-nosed monkeys (Rhinopithecus brelichi) at Yangaoping, Guizhou, China. Acta Theriol. Sin. 2009, 29, 326–331. [Google Scholar]
- Tilson, R.L.; Tenaza, R.R. lnterspecific Spacing Between Gibbons (Hylobates klossii) and Langurs (Presbytis potenziani) on Siberut Island, Indonesia. Am. J. Primatol. 1982, 2, 355–361. [Google Scholar] [CrossRef]
- Osman, N.A.; Abdul-Latiff, M.A.B.; Mohd-Ridwan, A.R.; Yaakop, S.; Nor, S.M.; Md-Zain, B.M. Diet Composition of the Wild Stump-Tailed Macaque (Macaca arctoides) in Perlis State Park, Peninsular Malaysia, Using a Chloroplast tRNL DNA Metabarcoding Approach: A Preliminary Study. Animals 2020, 10, 2215. [Google Scholar] [CrossRef]
- Guo, Y.; Zhou, J.; Xie, J.; Garber, P.A.; Bruford, M.; Ren, B.; Li, D.; Zhou, J. Altitudinal ranging of the Guizhou golden monkey (Rhinopithecus brelichi): Patterns of habitat selection and habitat use. Glob. Ecol. Conserv. 2018, 16, e473. [Google Scholar] [CrossRef]
- Mallott, E.K.; Garber, P.A.; Malhi, R.S. trnL outperforms rbcL as a DNA metabarcoding marker when compared with the observed plant component of the diet of wild white-faced capuchins (Cebus capucinus, Primates). PLoS ONE 2018, 13, e199556. [Google Scholar] [CrossRef]
- Sengupta, A.; Radhakrishna, S. Influence of Fruit Availability on Fruit Consumption in a Generalist Primate, the Rhesus Macaque Macaca mulatta. Int. J. Primatol. 2016, 37, 703–717. [Google Scholar] [CrossRef]
- Britt, A.; Randriamandratonirina, N.J.; Glasscock, K.D.; Iambana, B.R. Diet and Feeding Behaviour of Indri indri in a Low-Altitude Rain Forest. Folia Primatol. 2002, 73, 225–239. [Google Scholar] [CrossRef]
- Tsuji, Y.; Hanya, G.; Grueter, C.C. Feeding strategies of primates in temperate and alpine forests: Comparison of Asian macaques and colobines. Primates 2013, 54, 201–215. [Google Scholar] [CrossRef] [PubMed]
- Tuyisingize, D.; Eckardt, W.; Caillaud, D.; Kaplin, B.A. High flexibility in diet and ranging patterns in two golden monkey (Cercopithecus mitis kandti) populations in Rwanda. Am. J. Primatol. 2022, 84, e23347. [Google Scholar] [CrossRef]
- Wang, C.; Su, H.J.; Hu, C.S.; Shi, L.; Zhang, M.M. Spatial distribution of sympatric Rhinopithecus brelichi and Macaca thibetana in Fanjingshan National Nature Reserve, Guizhou, China. Acta Theriol. Sin. 2020, 40, 193–200. [Google Scholar] [CrossRef]
- Fleagle, J.G.; Meldrum, D.J. Locomotor behavior and skeletal morphology of two sympatric pitheciine monkeys, Pithecia pithecia and Chiropotes satanas. Am. J. Primatol. 1988, 16, 227–249. [Google Scholar] [CrossRef] [PubMed]
- Dagosto, M. Primate Origins and Evolution. A phylogenetic reconstruction. Int. J. Primatol. 1991, 12, 77–79. [Google Scholar] [CrossRef]
- Ford, S.M.; Davis, L.C. Systematics and body size: Implications for feeding adaptations in new world monkeys. Am. J. Phys. Anthropol. 1992, 88, 415–468. [Google Scholar] [CrossRef]
- Li, X.T. The World’s Endangered and Extinct Mammals; Knowledge Publishing House: Beijing, China, 2002; p. 4. [Google Scholar]
- Quan, G.Q.; Xie, J.H. Data on the Guizhou subspecies Rhinopithecus roxellanae brelichi Thomas. Acta Theriol. Sin. 1981, 2, 113–116. [Google Scholar] [CrossRef]
- Zhou, Q.H.; Huang, C.M. Advances in ecological research on the limestone langurs in China. Acta Theriol. Sin. 2021, 41, 59–70. [Google Scholar] [CrossRef]
- Hu, G. Distribution and conservation status of black langur resources in China. In Proceedings of the 9th Member Congress and 10th Academic Symposium of Sichuan Zoological Society; Sichuan Zoological Society: Chengdu, China, 2011. [Google Scholar]
- Anthony, R.E.; Sinclair, A.R. Wildlife Ecology, Conservation and Management; Wiley-Blackwel: New York, NY, USA, 2006; pp. 60–77. [Google Scholar]
- Li, J.Y. Abundance of Diet Plants and Seasonal Changes of Diet Nutrients of Rhinopithecus roxellana in Shennongjia Natural Reserve, China. Ph.D. Thesis, Beijing Forestry University, Beijing, China, 2015. [Google Scholar]
- Raubenheimer, D.; Simpson, S.J. Organismal Stoichiometry: Quantifying Non-Independence among Food Components. Ecology 2004, 85, 1203–1216. [Google Scholar] [CrossRef]
- Jiang, W.X.; Fu, X.C.; Tang, T.; Cai, Q.H. Community Structure and Niche of Macroinvertebrates in the Xiangxi River in Hubei, China. Chin. J. Appl. Environ. Biol. 2009, 03, 314–337. [Google Scholar] [CrossRef]
- Wang, C. Spatiotemporal Coexistence Pattern of Rhinopithecus brelichi and Macaca thibetana in Fanjingshan Nature Reserve based on Infrared Camera. Master’s Thesis, Gui Zhou University, Guizhou, China, 2021. [Google Scholar]
- Tutin, C.E.G.; Fernandez, M.; Rogers, M.E.; Williamson, E.A.; McGrew, W.C.; Altmann, S.A.; Southgate, D.A.T.; Crowe, I.; Tutin, C.E.G.; Whiten, A.; et al. Foraging profiles of sympatric lowland gorillas and chimpanzees in the Lopé Reserve, Gabon. Philos. Trans. Biol. Sci. 1991, 334, 179–186. [Google Scholar] [CrossRef]
- Heymann, E.W.; Knogge, C.; Tirado, H.E. Vertebrate predation by sympatric tamarins, Saguinus mystax and Saguinus fuscicollis. Am. J. Primatol. 2000, 51, 153–158. [Google Scholar] [CrossRef]
- Wu, F.; Zhu, D.; Wen, P.; Tang, Z.; Bao, L.; Guan, Y.; Ge, J.; Wang, H. Domestic Cattle in a National Park Restricting the Sika Deer Due to Diet Overlap. Animals 2023, 13, 561. [Google Scholar] [CrossRef] [PubMed]
- Schoener, T.W. Resource Partitioning in Ecological Communities. Science 1974, 185, 27–39. [Google Scholar] [CrossRef] [PubMed]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Group Information | Sample Number |
|---|---|
| Tibetan macaques | DH1, DH2, DH3, DH4, LJB1, LJB2, EB2, KZB2, KZB4, DH_a, HTP_a, KZB_a, KZB_c, EB_b, LJB_a, LJB_b, LJB_c, LJB_d |
| Grey snub-nosed monkeys | BP1, BP2, EB1, KZB1, KZB3, BP_c, BP_e, KZB_b, KZB_d, KZB_e, EB_a |
| B | Dij | |||
|---|---|---|---|---|
| Family | Genus | Family | Genus | |
| Tibetan macaques | 9.3865 | 10.2150 | 0.9998 | 0.9930 |
| Grey snub-nosed monkeys | 8.1951 | 9.2386 | ||
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yue, L.; Wang, C.; Meng, B.; Xie, B.; Cao, H.; Su, H.; Zhang, M. The Food Niche Overlap and Interspecific Relationship between the Sympatric Tibetan Macaque and Grey Snub-Nosed Monkey. Animals 2023, 13, 2536. https://doi.org/10.3390/ani13152536
Yue L, Wang C, Meng B, Xie B, Cao H, Su H, Zhang M. The Food Niche Overlap and Interspecific Relationship between the Sympatric Tibetan Macaque and Grey Snub-Nosed Monkey. Animals. 2023; 13(15):2536. https://doi.org/10.3390/ani13152536
Chicago/Turabian StyleYue, Li, Cheng Wang, Bingshun Meng, Bo Xie, Heqin Cao, Haijun Su, and Mingming Zhang. 2023. "The Food Niche Overlap and Interspecific Relationship between the Sympatric Tibetan Macaque and Grey Snub-Nosed Monkey" Animals 13, no. 15: 2536. https://doi.org/10.3390/ani13152536
APA StyleYue, L., Wang, C., Meng, B., Xie, B., Cao, H., Su, H., & Zhang, M. (2023). The Food Niche Overlap and Interspecific Relationship between the Sympatric Tibetan Macaque and Grey Snub-Nosed Monkey. Animals, 13(15), 2536. https://doi.org/10.3390/ani13152536