
Morphology of Starch Particles along the Passage through the Gastrointestinal Tract in Laboratory Mice Fed Extruded and Pelleted Diets

 , ,
, ,  and
and
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Animals and Diets
2.2. Analyses
2.2.1. Diet
2.2.2. Stereomicroscopy
2.2.3. Scanning Electron Microscopy
3. Results
3.1. Diets
3.2. Macroscopic Presentation
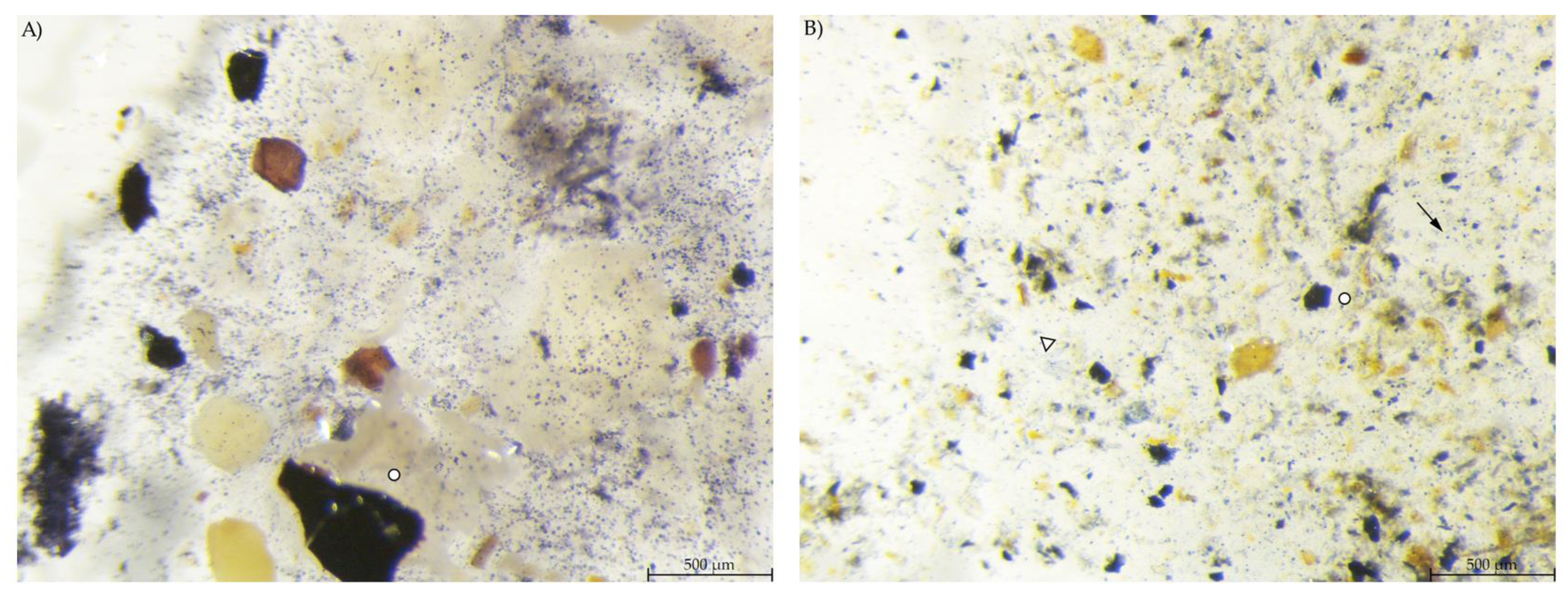
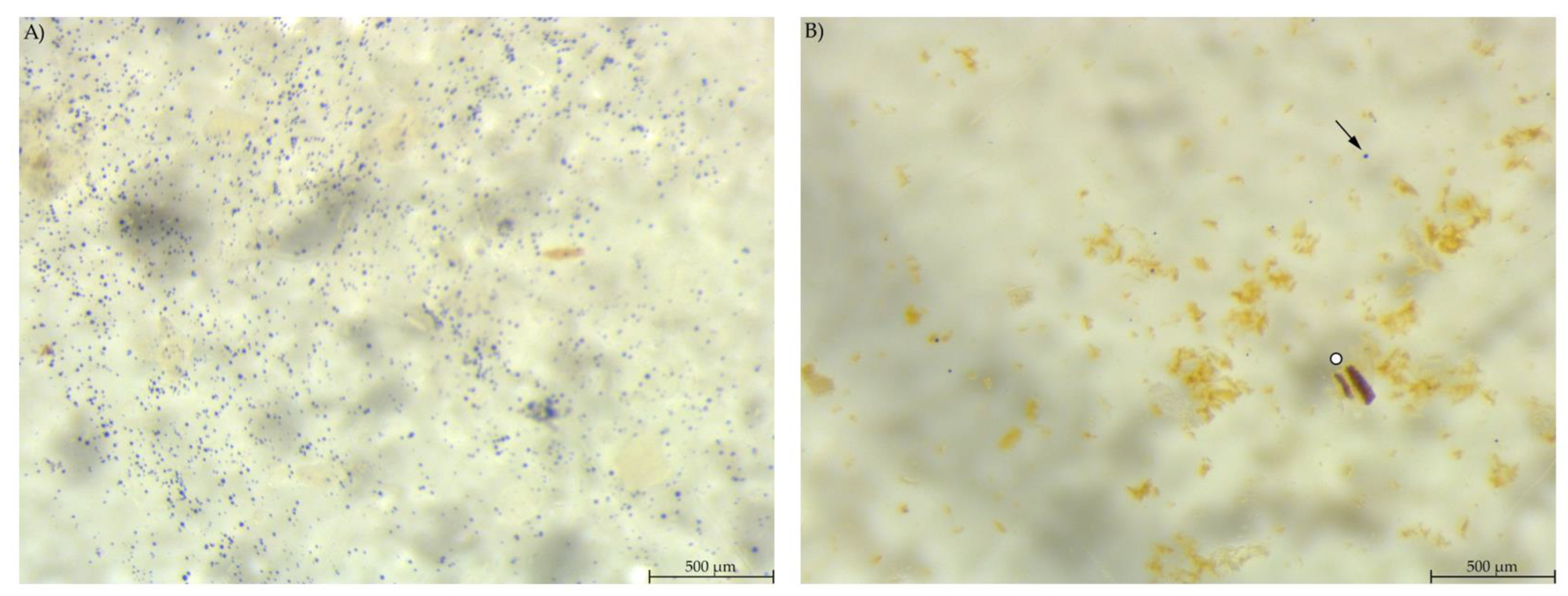
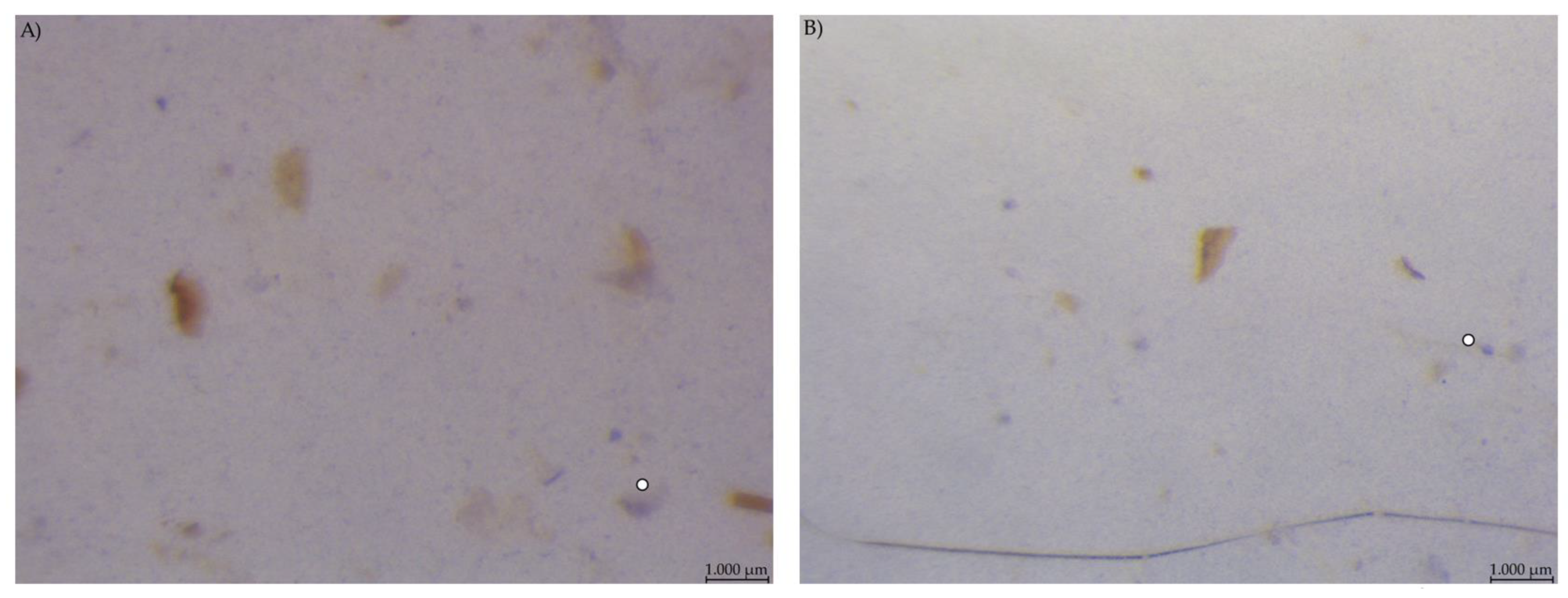
3.3. Stereomicroscopy
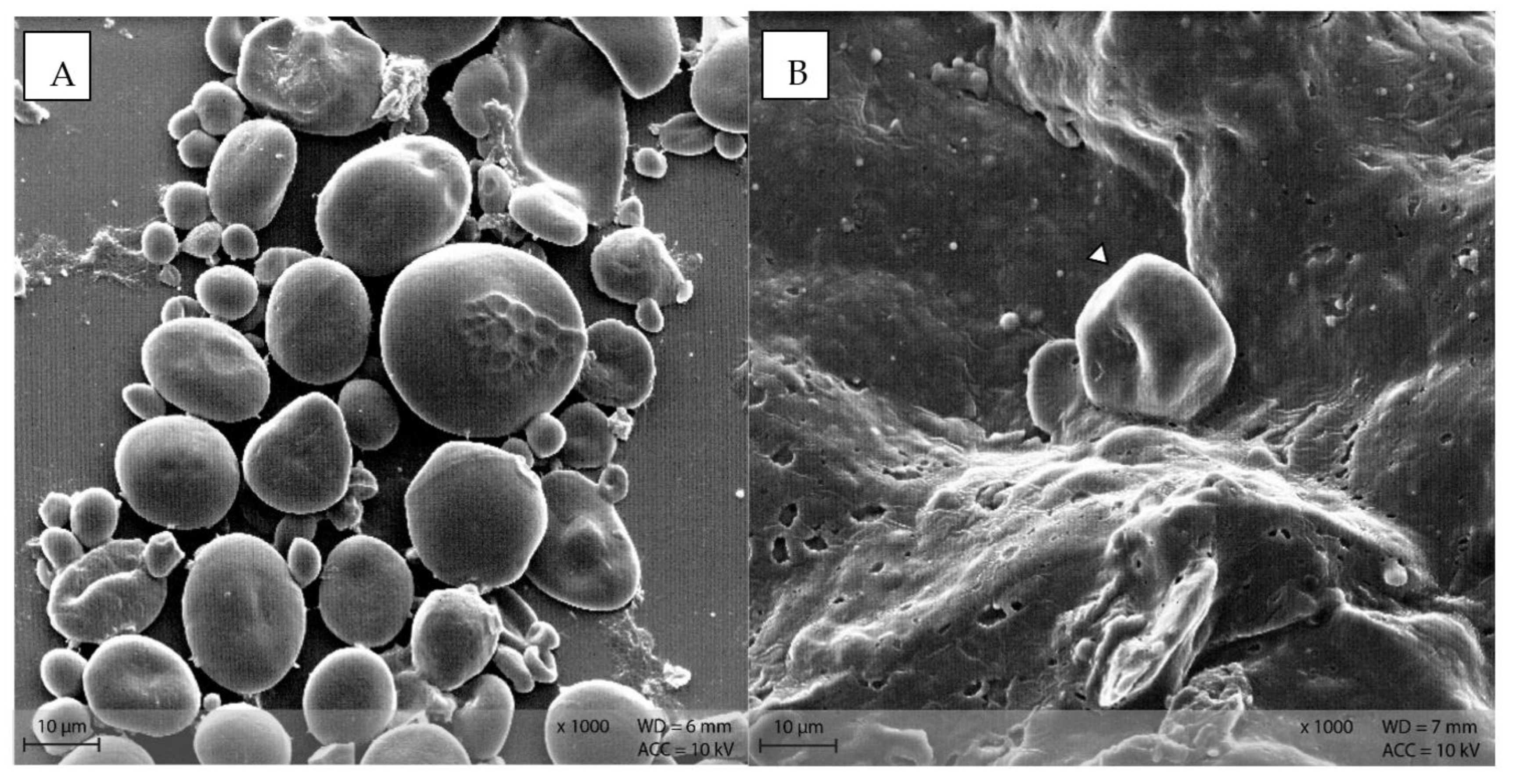
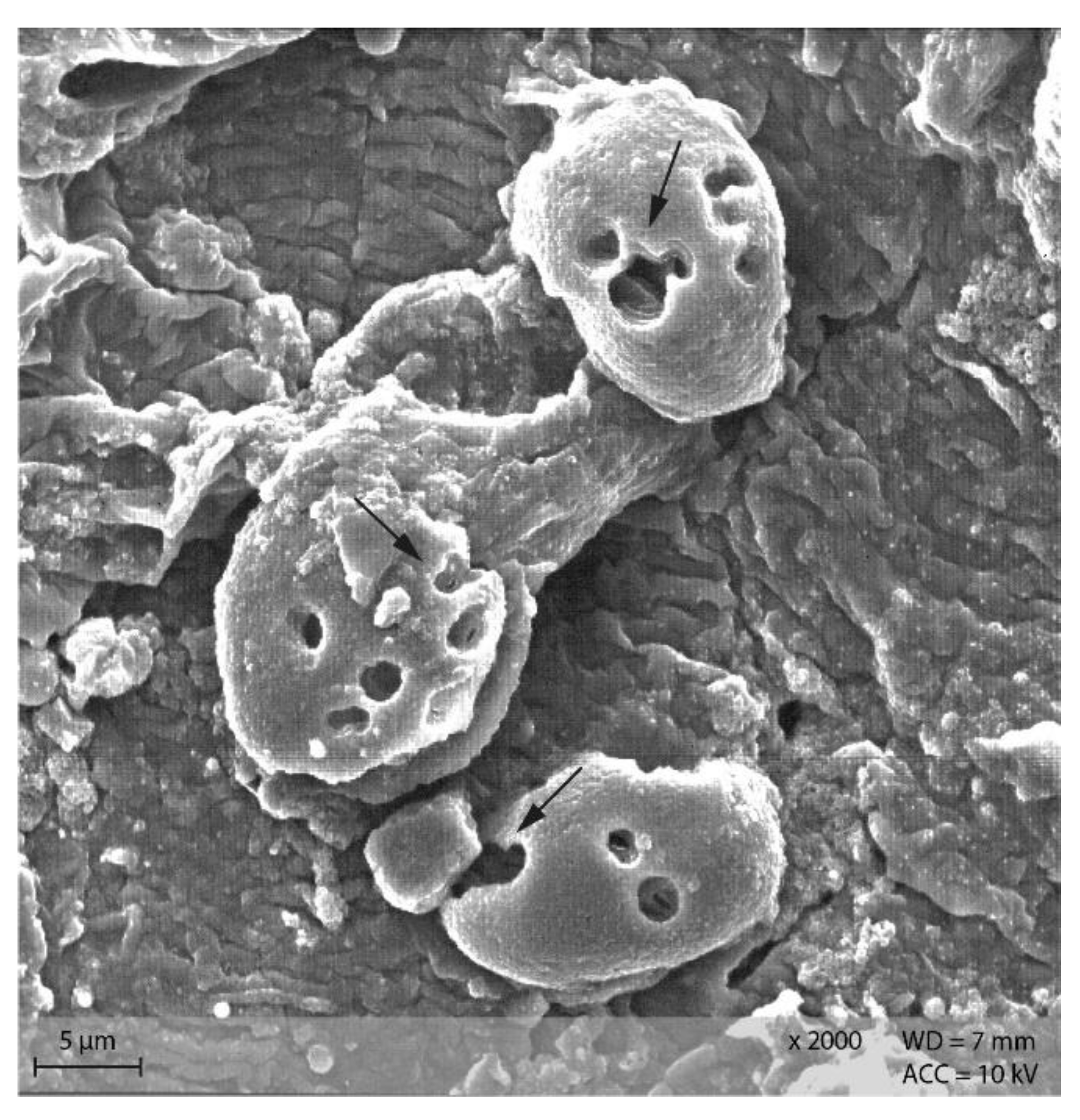
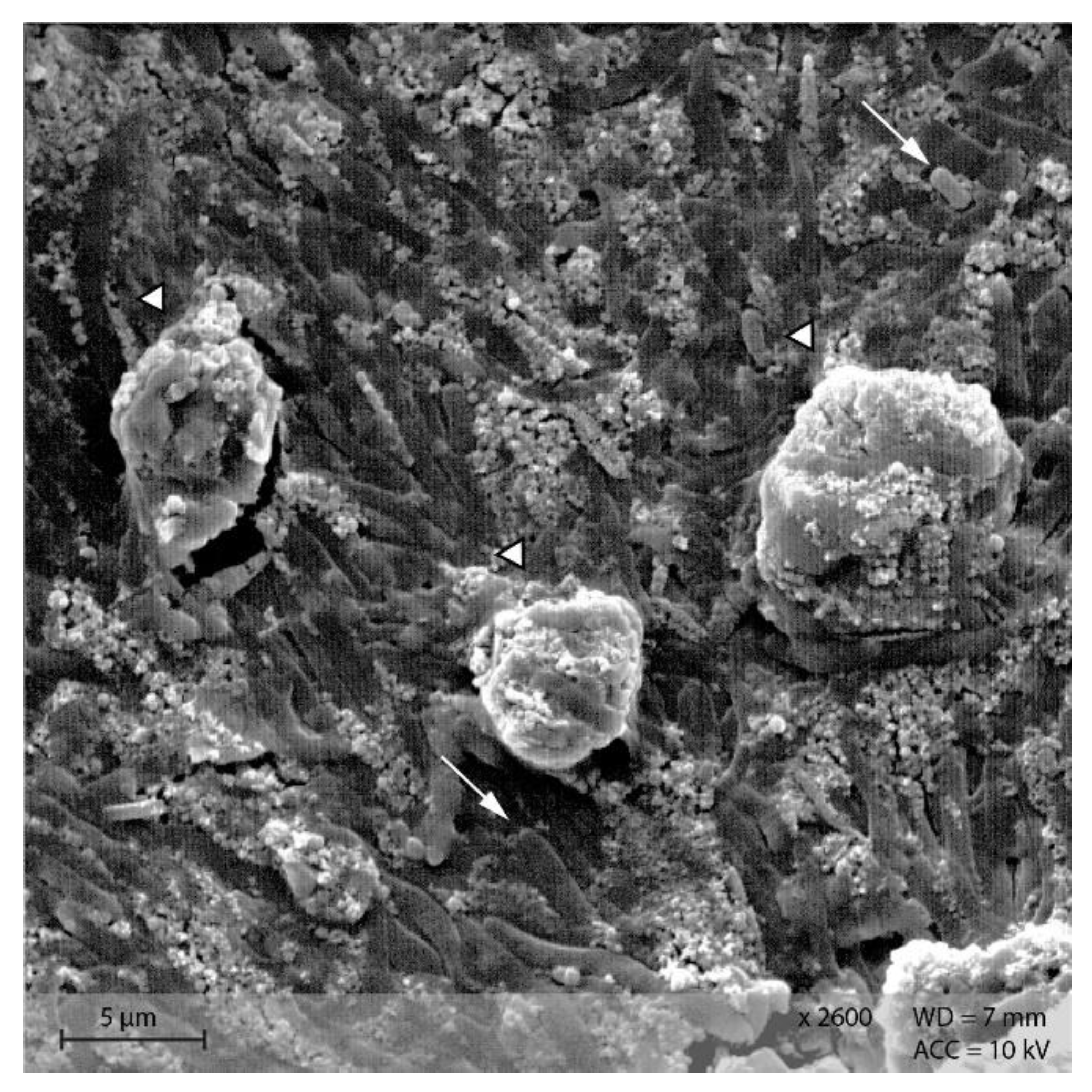
3.4. Scanning Electron Microscopy
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Conflicts of Interest
References
- Stevens, C.E.; Hume, I.D. Comparative Physiology of the Vertebrate Digestive System; Cambridge University Press: Cambridge, UK, 2004. [Google Scholar]
- Snipes, R.L. Anatomy of the cecum of the laboratory mouse and rat. Anat. Embryol. 1981, 162, 455–474. [Google Scholar] [CrossRef] [PubMed]
- Sakaguchi, E. Digestive strategies of small hindgut fermenters. Anim. Sci. J. 2003, 74, 327–337. [Google Scholar] [CrossRef]
- Svihus, B.; Uhlen, A.K.; Harstad, O.M. Effect of starch granule structure, associated components and processing on nutritive value of cereal starch: A review. Anim. Feed. Sci. Technol. 2005, 122, 303–320. [Google Scholar] [CrossRef]
- Gidley, M.; Flanagan, B.; Sharpe, K.; Sopade, P. Starch digestion in monogastrics–mechanisms and opportunities. Recent Adv. Anim. Nutr. 2011, 18, 207–213. [Google Scholar]
- Skoch, E.; Binder, S.; Deyoe, C.; Allee, G.; Behnke, K. Effects of steam pelleting conditions and extrusion cooking on a swine diet containing wheat middlings. J. Anim. Sci. 1983, 57, 929–935. [Google Scholar] [CrossRef]
- Asp, N.; Bjorck, I. Nutritional properties of extruded foods. Extrus. Cook. 1983, 57, 929–935. [Google Scholar]
- Böswald, L.; Kienzle, E. Differences in starch gelatinisation between differently processed laboratory rodent maintenance diets labelled as identical. In Proceedings of the 73rd Society of Nutrition Physiology (GfE) Conference, Goettingen, Germany, 13–15 March 2019. [Google Scholar]
- Leszczyñski, W.A. Resistant starch–classification, structure, production. Pol. J. Food Nutr. Sci. 2004, 13, 37–50. [Google Scholar]
- Asp, N.-G.; Van Amelsvoort, J.; Hautvast, J. Nutritional implications of resistant starch. Nutr. Res. Rev. 1996, 9, 1–31. [Google Scholar] [CrossRef] [Green Version]
- Ottenhof, M.-A.; Farhat, I.A. Starch retrogradation. Biotechnol. Genet. Eng. Rev. 2004, 21, 215–228. [Google Scholar] [CrossRef] [Green Version]
- Medel, P.; Latorre, M.; De Blas, C.; Lázaro, R.; Mateos, G. Heat processing of cereals in mash or pellet diets for young pigs. Anim. Feed. Sci. Technol. 2004, 113, 127–140. [Google Scholar] [CrossRef]
- Böswald, L.F.; Wenderlein, J.; Straubinger, R.K.; Ulrich, S.; Kienzle, E. Processing Matters in Nutrient-Matched Laboratory Diets for Mice—Energy and Nutrient Digestibility. Animals 2021, 11, 523. [Google Scholar] [CrossRef]
- Kienzle, E.; Pohlenz, J.; Radicke, S. Morphology of starch digestion in the horse. J. Vet. Med. Ser. A 1997, 44, 207–221. [Google Scholar] [CrossRef]
- Meyer, H.; Radicke, S.; Kienzle, E.; Wilke, S.; Kleffken, D.; Illenseer, M. Investigations on preileal digestion of starch from grain, potato and manioc in horses. J. Vet. Med. Ser. A 1995, 42, 371–381. [Google Scholar] [CrossRef]
- Champ, M.; Szylit, O.; Gallant, D. The influence of microflora on the breakdown of maize starch granules in the digestive tract of chicken. Poult. Sci. 1981, 60, 179–187. [Google Scholar] [CrossRef]
- Davis, A.B. Scanning Electron Microscopy of Monogastric and Ruminant Digestion of Sorghum Grain. Master’s Thesis, Kansas State University, Manhattan, KS, USA, 1973. [Google Scholar]
- Martens, B.M.; Gerrits, W.J.; Bruininx, E.M.; Schols, H.A. Amylopectin structure and crystallinity explains variation in digestion kinetics of starches across botanic sources in an in vitro pig model. J. Anim. Sci. Biotechnol. 2018, 9, 91. [Google Scholar] [CrossRef] [Green Version]
- Lee, T.-T.; Huang, Y.-F.; Chiang, C.-C.; Chung, T.-K.; Chiou, P.W.-S.; Yu, B. Starch characteristics and their influences on in vitro and pig prececal starch digestion. J. Agric. Food Chem. 2011, 59, 7353–7359. [Google Scholar] [CrossRef]
- Bochnia, M.; Walther, S.; Schenkel, H.; Romanowski, K.; Zeyner, A. Comparison of scanning electron microscopic examination of oats, barley and maize grains with the analysed degree of starch breakdown and glycamic responses in horse. Int. J. Sci. Res. Sci. Technol. 2015, 1, 81–84. [Google Scholar]
- Wenderlein, J.; Böswald, L.F.; Ulrich, S.; Kienzle, E.; Neuhaus, K.; Lagkouvardos, I.; Zenner, C.; Straubinger, R.K. Processing Matters in Nutrient-Matched Laboratory Diets for Mice—Microbiome. Animals 2021, 11, 862. [Google Scholar] [CrossRef]
- Naumann, C.; Bassler, R. Die Chemische Untersuchung von Futtermitteln. Band III: Methodenbuch; Verlag, J., Ed.; Naumann: Neudamm, Germany, 1986. [Google Scholar]
- Treuting, P.M.; Dintzis, S.M.; Montine, K.S. Comparative Anatomy and Histology: A Mouse, Rat, and Human Atlas; Academic Press: Cambridge, MA, USA, 2017. [Google Scholar]
- Ghoshal, N.; Bal, H. Comparative morphology of the stomach of some laboratory mammals. Lab. Anim. 1989, 23, 21–29. [Google Scholar] [CrossRef]
- Alexander, F.; Davies, M.E. Production and fermentation of lactate by bacteria in the. 5 alimentary canal of the horse and pig. J. Comp. Pathol. 1963, 73, 1–8. [Google Scholar] [CrossRef]
- Hagen, K.B.; Müller, D.W.; Ortmann, S.; Kreuzer, M.; Clauss, M. Digesta kinetics in two arvicoline rodents, the field vole (Microtus agrestis) and the steppe lemming (Lagurus lagurus). Mamm. Biol. 2018, 89, 71–78. [Google Scholar] [CrossRef]
- Cotta, M.; Forster, R. The family Lachnospiraceae, including the genera Butyrivibrio, Lachnospira and Roseburia. Prokaryotes 2006, 4, 1002–1021. [Google Scholar]
- Vacca, M.; Celano, G.; Calabrese, F.M.; Portincasa, P.; Gobbetti, M.; De Angelis, M. The controversial role of human gut lachnospiraceae. Microorganisms 2020, 8, 573. [Google Scholar] [CrossRef]
- Lagkouvardos, I.; Lesker, T.R.; Hitch, T.C.; Gálvez, E.J.; Smit, N.; Neuhaus, K.; Wang, J.; Baines, J.F.; Abt, B.; Stecher, B. Sequence and cultivation study of Muribaculaceae reveals novel species, host preference, and functional potential of this yet undescribed family. Microbiome 2019, 7, 28. [Google Scholar] [CrossRef] [Green Version]
- Xu, S.S.; Xiang, Z.J.; Bin, L.; Jing, L.; Bin, Z.; Jiao, Y.J.; Kun, S.R. Preparation and physical characteristics of resistant starch (type 4) in acetylated indica rice. Food Chem. 2012, 134, 149–154. [Google Scholar]
- Zhang, H.; Jin, Z. Preparation of products rich in resistant starch from maize starch by an enzymatic method. Carbohydr. Polym. 2011, 86, 1610–1614. [Google Scholar] [CrossRef]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Diet PEL | Diet EXT | |
|---|---|---|
| Gross energy (MJ/kg) | 16.6 | 17.3 |
| Dry matter (%) | 89.5 | 88.5 |
| Crude protein (%) | 18.7 | 22.5 |
| Crude fat (%) | 2.5 | 5.1 |
| Crude ash (%) | 4.8 | 6.1 |
| Crude fiber (%) | 5.2 | 5.8 |
| Starch (%) | 43.0 | 28.0 |
| Degree of starch gelatinization (% of starch) | 17.0 | 70.0 |
| Total dietary fiber (%) | 15.9 | 23.3 |
| Soluble dietary fiber (%) | 2.0 | 3.9 |
| Insoluble dietary fiber (%) | 13.9 | 19.4 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wenderlein, J.; Kienzle, E.; Straubinger, R.K.; Schöl, H.; Ulrich, S.; Böswald, L.F. Morphology of Starch Particles along the Passage through the Gastrointestinal Tract in Laboratory Mice Fed Extruded and Pelleted Diets. Animals 2022, 12, 952. https://doi.org/10.3390/ani12080952
Wenderlein J, Kienzle E, Straubinger RK, Schöl H, Ulrich S, Böswald LF. Morphology of Starch Particles along the Passage through the Gastrointestinal Tract in Laboratory Mice Fed Extruded and Pelleted Diets. Animals. 2022; 12(8):952. https://doi.org/10.3390/ani12080952
Chicago/Turabian StyleWenderlein, Jasmin, Ellen Kienzle, Reinhard K. Straubinger, Heidrun Schöl, Sebastian Ulrich, and Linda Franziska Böswald. 2022. "Morphology of Starch Particles along the Passage through the Gastrointestinal Tract in Laboratory Mice Fed Extruded and Pelleted Diets" Animals 12, no. 8: 952. https://doi.org/10.3390/ani12080952
APA StyleWenderlein, J., Kienzle, E., Straubinger, R. K., Schöl, H., Ulrich, S., & Böswald, L. F. (2022). Morphology of Starch Particles along the Passage through the Gastrointestinal Tract in Laboratory Mice Fed Extruded and Pelleted Diets. Animals, 12(8), 952. https://doi.org/10.3390/ani12080952