
Effects of Dietary Supplementation with Glutamine on the Immunity and Intestinal Barrier Gene Expression in Broiler Chickens Infected with Salmonella Enteritidis


Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Salmonella Enteritidis, Gln
2.2. Broilers, Management, Experimental Diets, and Experimental Design
2.3. Sample Collection and Procedures
2.4. Enzyme Activity Assay and Detection of Serum Immunoglobulin Populations
2.5. Intestinal Mucosa Sample RNA Extraction and qRT–PCR Analysis
2.6. Statistical Methods
3. Results
3.1. Growth Performance
3.2. LZM, IgA, IgG, and IgM Concentrations in Serum
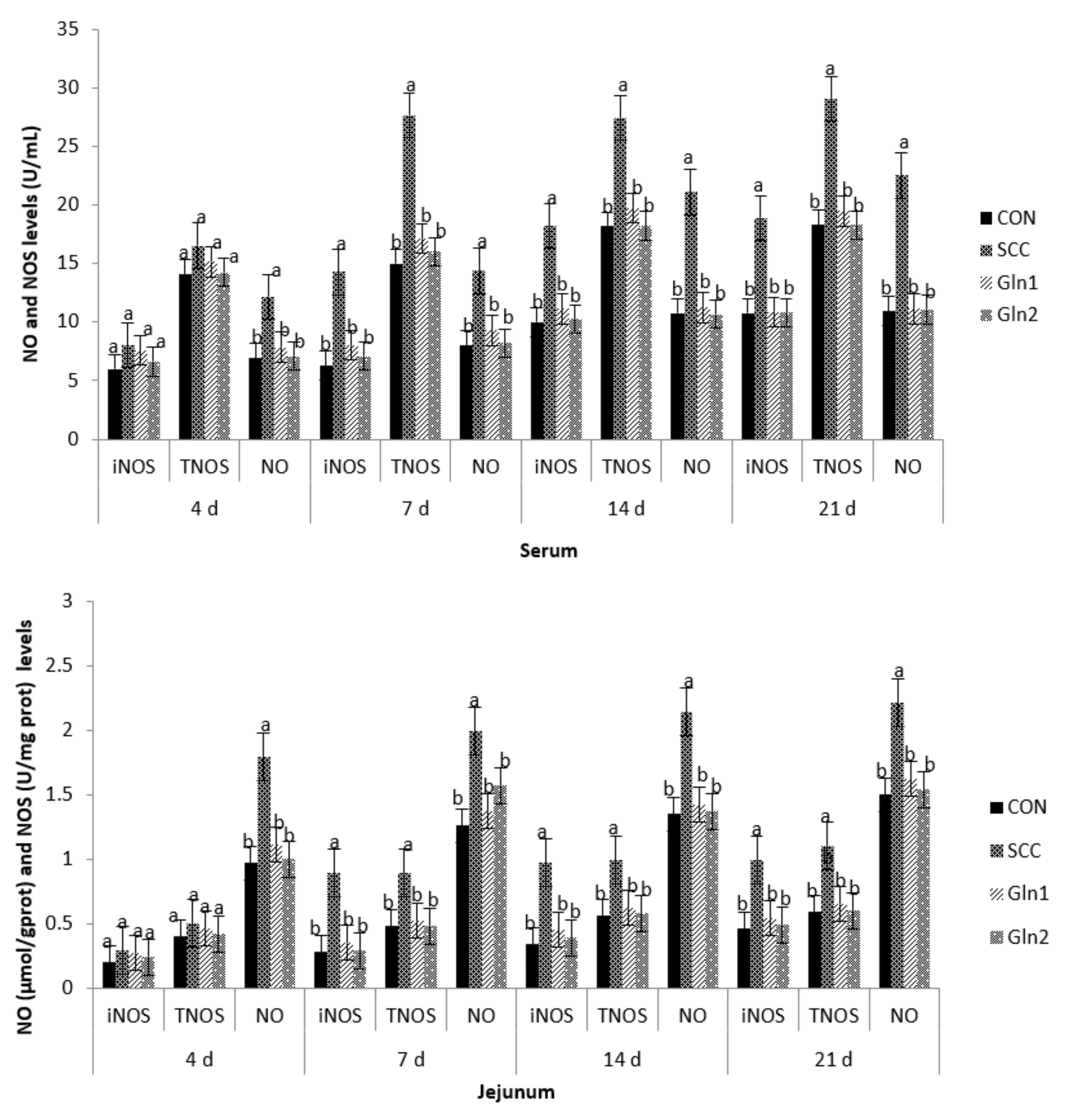
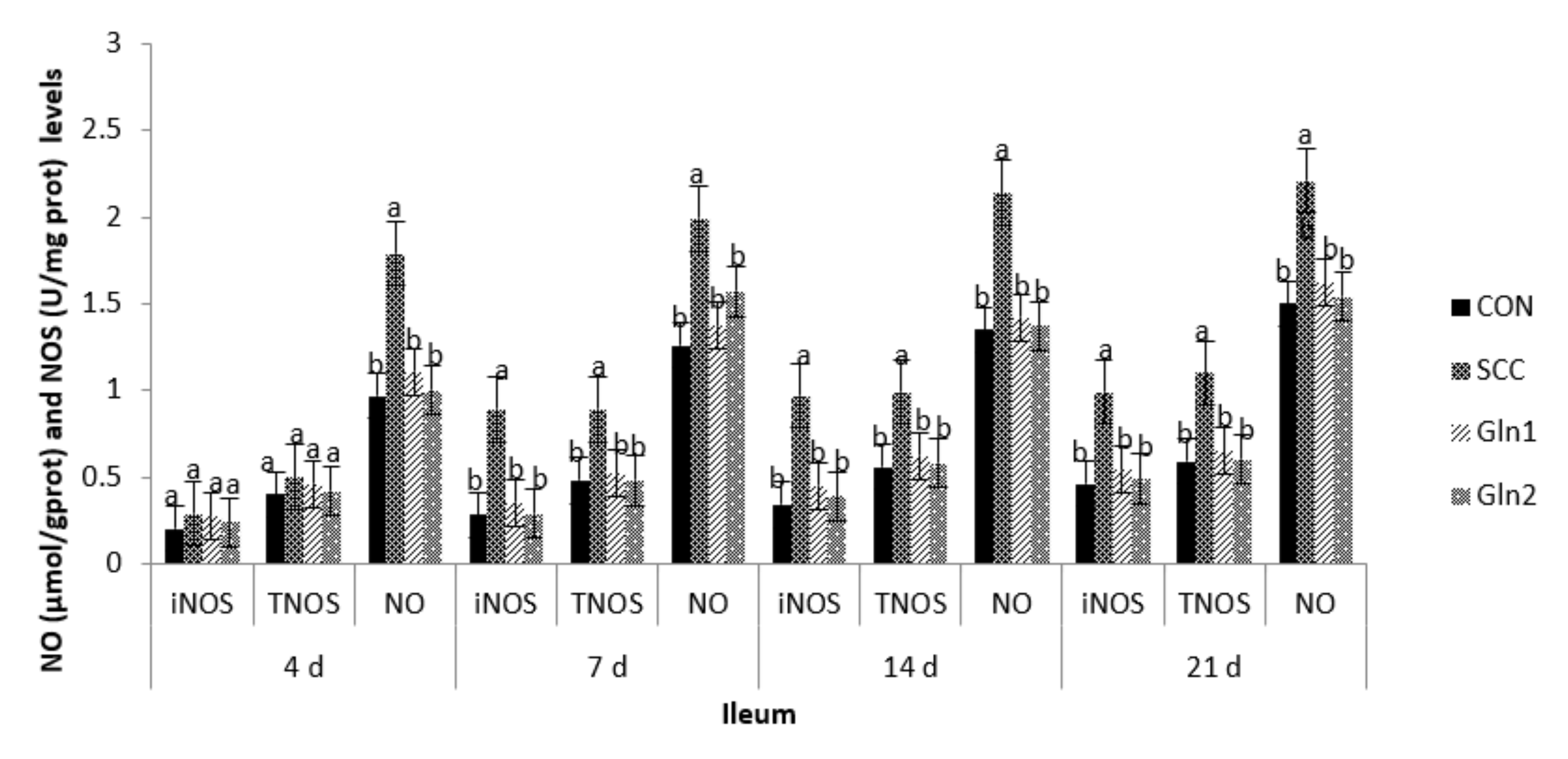
3.3. NO and NOS Levels in Serum and Intestinal Mucosa
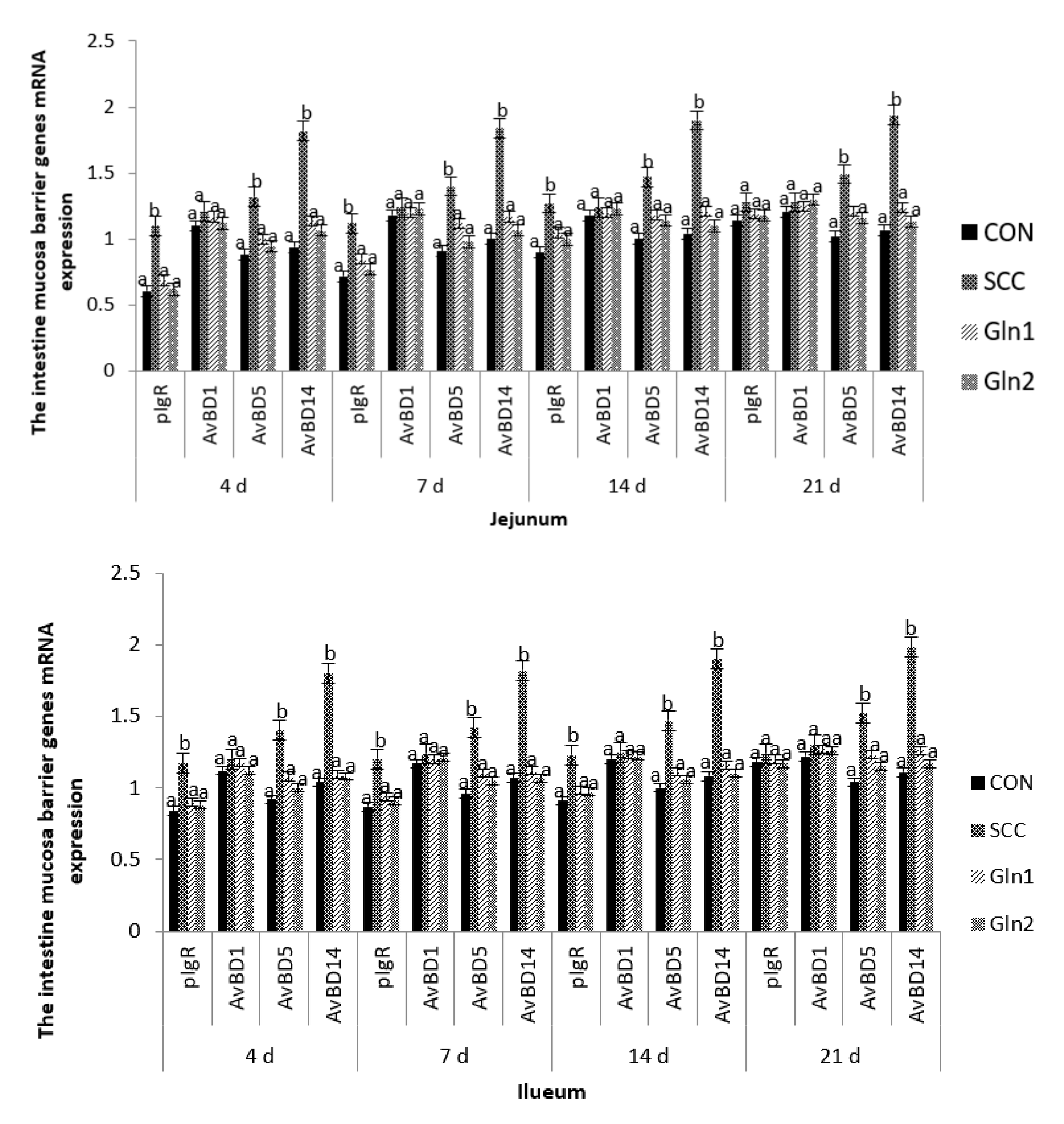
3.4. The Intestinal Mucosa Barrier Genes’ mRNA Expression Levels
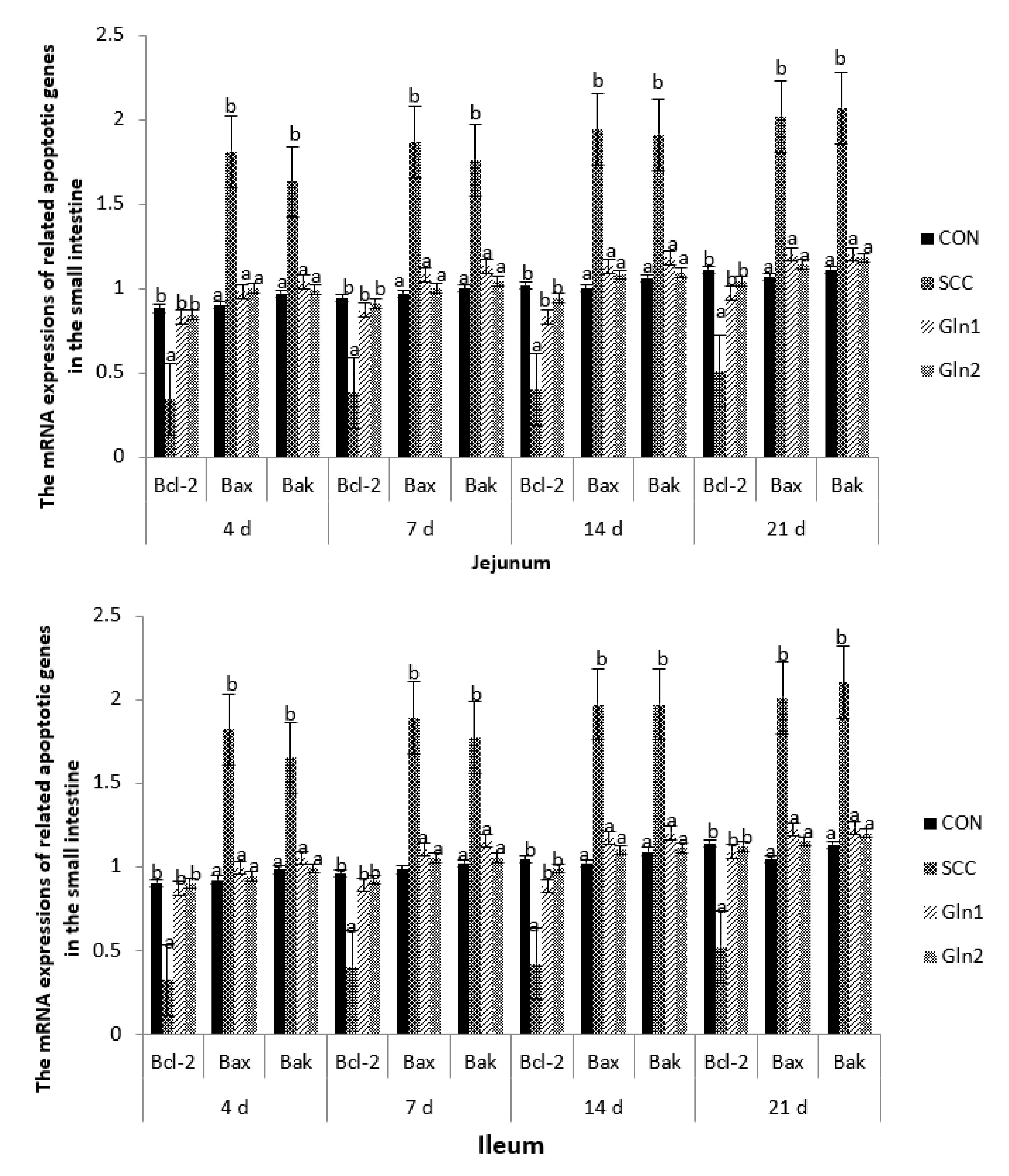
3.5. The Related Apoptotic Genes’ mRNA Expression Levels in the Intestinal Mucosa
4. Discussion
4.1. Effect of Dietary Gln on the Growth Performance of Broilers Infected with Salmonella Enteritidis
4.2. Effect of Dietary Gln on LZM and Antibody Levels in the Serum of Broilers Infected with Salmonella Enteritidis
4.3. Effect of Dietary Gln on the NO and NOS Levels in the Serum and Intestinal Mucosa of Broilers Infected with Salmonella Enteritidis
4.4. Effect of Dietary Gln on the Intestine Mucosa Barrier Genes’ mRNA Expressions of Broilers Infected with Salmonella Enteritidis
4.5. Effect of Dietary Gln on the mRNA Expressions of Related Apoptotic Genes in the Small Intestine of Broilers Infected with Salmonella Enteritidis
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Khin, K.Z.M.; Perot, S.; Michelle, M.H.; Ganrea, C.; Chang, H.C.; Lydia, G.; Elizabeth, A.M.; Zhou, H.J. Salmonella enter-ica Serovars Enteritidis Infection Alters the Indigenous Microbiota Diversity in Young Layer Chicks. Front. Vet. Sci. 2015, 2, 61. [Google Scholar]
- Guo, X.H.; Li, D.F.; Lu, W.Q.; Piao, X.S.; Chen, X.L. Salmonella: A review on pathogenesis, epidemiology and antibiotic re-sistance. Front. Life Sci. 2015, 8, 284–293. [Google Scholar]
- Westrom, B.; ArevaloSureda, E.; Pierzynowska, K.; Pierzynowski, S.G.; Perez-Cano, F.J. New insights in intestinal oxidative stress damage and the health intervention effects of nutrients: A review. J. Funct. Foods 2020, 75, 104248. [Google Scholar]
- Hou, Q.; Ye, L.; Liu, H.; Huang, L.; Yang, Q.; Turner, J.R.; Yu, Q. The research progress on intestinal stem cells and itts relationship with intestinal microbiota. Front. Immunol. 2017, 8, 599. [Google Scholar] [CrossRef] [Green Version]
- Xi, P.; Jiang, Z.; Dai, Z.; Li, X.; Yao, K.; Zheng, C.; Lin, Y.; Wang, J.; Wu, G. Regulation of protein turnover by l-glutamine in porcine intestinal epithelial cells. J. Nutr. Biochem. 2012, 23, 1012–1017. [Google Scholar] [CrossRef]
- Lash, L.H.; Hagen, T.M.; Jones, D.P. Exogenous glutathione protects intestinal epithelial cells from oxidative injury. Proc. Natl. Acad. Sci. USA 1986, 83, 4641–4645. [Google Scholar] [CrossRef] [Green Version]
- Rao, R.K.; Li, L.; Baker, R.D.; Baker, S.S.; Gupta, A. Glutathione oxidation and PTPase inhibition by hydrogen peroxide in Caco-2 cell monolayer. Am. J. Physiol.-Gastrointest. Liver Physiol. 2000, 279, G332–G340. [Google Scholar] [CrossRef]
- Kelly, N.; Friend, K.; Boyle, P.; Zhang, X.R.; Wong, C.; Hackam, D.J.; Zamora, R.; Ford, H.R.; Upperman, J.S. The role of the glutathione antioxidant system in gut barrier failure in a rodent model of experimental necrotizing enterocolitis. Surgery 2004, 136, 557–566. [Google Scholar] [CrossRef]
- Wu, Q.J.; Zheng, X.C.; Wang, T.; Zhang, T.Y. Effects of oridonin on immune cells, Th1/Th2 balance and the expression of BLys in the spleens of broiler chickens challenged with Salmonella Pullorum. Res. Vet. Sci. 2018, 119, 262–267. [Google Scholar] [CrossRef]
- Soderholm, J.D.; Yates, D.A.; Gareau, M.G.; Yang, P.C.; MacQueen, G.; Perdue, M.H. Neonatal maternal separation predis-poses adult rats to colonice barrier dysfunction in response to mild stress. Am. J. Physiol. Gastrointese Liver Physiol. 2002, 283, 1257–1263. [Google Scholar] [CrossRef] [Green Version]
- Jiang, Z.Y.; Sun, L.H.; Lin, Y.C.; Ma, X.Y.; Zheng, C.T.; Zhou, G.L.; Chen, F.; Zou, S.T. Effects of dietary glycyl-glutamine on growth performance, small intestinal integrity, and immune responses of weaning piglets challenged with lipopolysaccharide1. J. Anim. Sci. 2009, 87, 4050–4056. [Google Scholar] [CrossRef] [Green Version]
- Wijtten, P.J.A.; Van Der Meulen, J.; Verstegen, M.W.A. Intestinal barrier function and absorption in pigs after weaning: A review. Br. J. Nutr. 2011, 105, 967–981. [Google Scholar] [CrossRef]
- Sukhotnik, I.; Yulia, P.; Coran, A.G.; Janna, P.; Jacob, B.; Mogilner, J.G.; Berkowitz, D. Glutamine attenuates the inhibitory effect of methotrexate on TLR signaling during intestinal chemotherapy-induced mucositis in a rat. Nutr. Metab. 2014, 11, 17. [Google Scholar] [CrossRef] [Green Version]
- Shakeri, M.; Zulkifli, I.; Oskoueian, E.; Shakeri, M.; Oskoueian, A.; Ebrahimi, M. Response to Dietary Supplementation of Glutamine in Broiler Chickens Subjected to Transportation Stress. J. Faculty Veter. Med. 2016, 42, 122–131. [Google Scholar] [CrossRef]
- Groschwitz, K.R.; Ahrens, R.; Osterfeld, H.; Gurish, M.F.; Han, X.; Abrink, M. Mast cells regulate homeostatic intestinal epi-thelial migration and barrier function by a chymase/Mcpt-4-dependent mechanism. Proc. Natl. Acad. Sci. USA 2009, 106, 22381–22386. [Google Scholar] [CrossRef] [Green Version]
- Smith, F.; Clark, J.E.; Overman, B.L.; Tozel, C.C.; Huang, J.H.; Rivier, J.E.F.; Blisklager, A.T.; Moeser, A.J. Early weaning stress impairs development of mucosal barrier function in the porcine intestine. Am. J. Physiol. Liver Physiol. 2010, 298, G352–G363. [Google Scholar] [CrossRef] [Green Version]
- Wu, G.; Bazer, F.W.; Johnson, G.A.; Knabe, D.A.; Burghardt, R.C.; Spencer, T.E.; Li, X.L.; Wang, J.J. Triennial Growth Sym-posium: Important roles for L-glutamine in swine nutrition and production. J. Anim. Sci. 2011, 89, 2017–2030. [Google Scholar] [CrossRef] [Green Version]
- José, M.M.; Matés, J.M.; Segura, J.A.; Alonso, F.J.; Márquez, J. Pathways from glutamine to apoptosis. Front. Biosci. 2006, 11, 3164–3180. [Google Scholar] [CrossRef] [Green Version]
- Sandri, M. FOXOphagy path to inducing stress resistance and cell survival. Nat. Cell Biol. 2012, 14, 786–788. [Google Scholar] [CrossRef]
- Sakiyama, T.; Musch, M.W.; Ropeleski, M.J.; Tsubouchi, H.; Chang, E.B. Glutamine Increases Autophagy Under Basal and Stressed Conditions in Intestinal Epithelial Cells. Gastroenterology 2009, 136, 924–932.e2. [Google Scholar] [CrossRef] [Green Version]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2(-Delta Delta C(T)) Method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef]
- Chalghoumi, R.; Théwis, A.; Beckers, Y.; Marcq, C.; Portetelle, D.; Schneider, Y.-J. Adhesion and Growth Inhibitory Effect of Chicken Egg Yolk Antibody (IgY) onSalmonella entericaSerovars Enteritidis and TyphimuriumIn Vitro. Foodborne Pathog. Dis. 2009, 6, 593–604. [Google Scholar] [CrossRef] [PubMed]
- Barrow, P.A.; Freintas, N.O.C. Pullorum disease and fowl typhoid-new thoughts on old diseases: A review. Avian Pathol. 2011, 40, 1–13. [Google Scholar] [CrossRef] [PubMed]
- Bohez, L.; Gantois, I.; Ducatelle, R.; Pasmans, F.; Dewulf, J.; Haesebrouck, F.; Van Immerseel, F. The Salmonella Pathogenicity Island 2 regulator ssrA promotes reproductive tract but not intestinal colonization in chickens. Veter- Microbiol. 2008, 126, 216–224. [Google Scholar] [CrossRef]
- Fasina, Y.O.; Bowers, J.B.; Hess, J.B.; McKee, S.R. Effect of dietary glutamine supplementation on Salmonella colonization in the ceca of young broiler chicks. Poult. Sci. 2010, 89, 1042–1048. [Google Scholar] [CrossRef] [PubMed]
- Zou, T.D.; Deng, C.X.; Wang, Z.R.; Ye, Y.L.; You, J.M. Dietary alanyl-glutamine improves growth performance of weaned piglets through maintaining intestinal morphology and digestion–absorption function. Animal 2019, 13, 1826–1833. [Google Scholar] [CrossRef]
- Wu, Q.J.; Zhu, D.D.; Wang, D.D.; Zhang, B.B.; Ren, A.; Bin Zhang, Z. Effects of dietary supplementation with glutamine on the lymphocyte proliferation and intestinal immune gene expression in broiler chickens infected with Salmonella Enteritidis. Res. Veter- Sci. 2021, 139, 18–24. [Google Scholar] [CrossRef]
- Cheng, Z.; Buentello, A.; Gatlin, D.M. Effects of dietary arginine and glutamine on growth performance, immune responses and intestinal structure of red drum, Sciaenops ocellatus. Aquaculture 2011, 319, 247–252. [Google Scholar] [CrossRef]
- Murakami, A.E.; Sakamoto, M.I.; Natali, M.R.M.; Souza, L.M.G.; Franco, J.R.G. Supplementation of Glutamine and Vitamin E on the Morphometry of the Intestinal Mucosa in Broiler Chickens. Poult. Sci. 2007, 86, 488–495. [Google Scholar] [CrossRef]
- Bartell, S.M.; Batal, A.B. The effect of supplemental glutamine on growth performance, development of the gastrointestinal tract and humoral immune response of broilers. J. Poult. Sci. 2007, 86, 1940–1947. [Google Scholar] [CrossRef]
- Soltan, M.A. Influence dietary supplementation on growth performance, small intestinal morphology, immune response and some blood parameters broiler chickens. Int. Poult. Sci. 2009, 1, 60–68. [Google Scholar] [CrossRef] [Green Version]
- Ayazi, M. The effect of dietary Glutamine supplementation on performance and blood parameter, carcass characteristics, quality and characteristics meat of broiler chickens under continuous heat stress condition. Int. J. Farm. Alli. Sci. 2014, 3, 1234–1242. [Google Scholar]
- Saurabh, S.; Sahoo, P.K. Lysozyme: An important defence molecule of fish innate immune system. Aquac. Res. 2008, 39, 223–239. [Google Scholar] [CrossRef]
- Demers, N.E.; Bayne, C.J. The immediate effects of stress on hormones and plasma lysozyme in rainbow trout. Dev. Comp. Immunol. 1997, 21, 363–373. [Google Scholar] [CrossRef]
- Jia, R.; Liu, B.-L.; Feng, W.-R.; Han, C.; Huang, B.; Lei, J.-L. Stress and immune responses in skin of turbot (Scophthalmus maximus) under different stocking densities. Fish Shellfish Immunol. 2016, 55, 131–139. [Google Scholar] [CrossRef]
- Li, X.; Ji, L.; Wu, L.; Gao, X.; Li, X.; Li, J.; Liu, Y. Effect of flow velocity on the growth, stress and immune responses of turbot (Scophthalmus maximus) in recirculating aquaculture systems. Fish Shellfish Immunol. 2018, 86, 1169–1176. [Google Scholar] [CrossRef] [PubMed]
- Mock, A.; Peters, G. Lysozyme activity in rainbow trout, Oncorhynchus mykiss (Walbaum) stressed by handling, transport, and water pollution. J. Fish. Biol. 1990, 37, 873–885. [Google Scholar]
- Caruso, C.; Buffa, S.; Candore, G.; Colonna-Romano, G.; Dunn-Walters, D.; Kipling, D.; Pawelec, G. Mechanisms of immunosenescence. Immun. Ageing 2009, 22, 6–10. [Google Scholar]
- Sahoo, P.K.; Mukherjee, S.C. The Effect of Dietary Glucan, Levamisole, Ascorbic Acid and a-Tocopherol on Disease Resistance and Non-Speci¢c Immunity Level in a£atoxin B1-induced immunocompromised rohu (Labeo rohita). In The Fifth Indian Fisheries Forum Proceedings; Ayyappan, S., Jena, J.K., Mohan Joseph, M., Eds.; AFSIB Mangalore and AoA: Bhuba-neswar, India, 2002; pp. 119–126. [Google Scholar]
- Sahoo, P.K.; Mukherjee, S.C. Immunomodulation by dietary vitamin C in healthy and a£atoxin B1-induced immunocom-promised rohu (Labeo rohita). Comp. Immunol. Microbiol. Infect. Dis. 2003, 26, 65–76. [Google Scholar]
- Sahoo, P.K.; Mukherjee, S.C. In£uence of high dietary a-tocopherol intakes on specific immune response, non-specific resistance factors and disease resistance of healthy and a£atoxin B1-induced immunocompromised Indian major carp, Labeo rohita (Hamilton). Aquacult. Nutr. 2002, 8, 159–167. [Google Scholar] [CrossRef]
- Kumari, J.; Sahoo, P.K. Dietary b-1,3 glucan potentiates innate immunity and disease resistance of Asian cat¢sh, Clarias ba-trachus (L.). J. Fish Dis. 2006, 29, 95–101. [Google Scholar] [CrossRef] [PubMed]
- Liu, G.; Ren, W.; Fang, J.; Hu, C.-A.A.; Guan, G.; Al-Dhabi, N.A.; Yin, J.; Duraipandiyan, V.; Chen, S.; Peng, Y.; et al. l-Glutamine and l-arginine protect against enterotoxigenic Escherichia coli infection via intestinal innate immunity in mice. Amino Acids 2017, 49, 1945–1954. [Google Scholar] [CrossRef] [PubMed]
- Ren, W.; Wang, K.; Yin, J.; Chen, S.; Liu, G.; Tan, B.; Wu, G.; Bazer, F.W.; Peng, Y.; Yin, Y. Glutamine-Induced Secretion of Intestinal Secretory Immunoglobulin A: A Mechanistic Perspective. Front. Immunol. 2016, 7, 503. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wieland, W.H.; Orzaez, D.; Lammers, A.; Parmentier, H.K.; Verstegen, M.W.A.; Schots, A. A functional polymeric immunoglobulin receptor in chicken (Gallus gallus) indicates ancient role of secretory IgA in mucosal immunity. Biochem. J. 2004, 380, 669–676. [Google Scholar] [CrossRef] [PubMed]
- Schroeder, H.W., Jr.; Cavacini, L. Structure and function of immunoglobulins. J. Allergy Clin. Immunol. 2010, 125, 41–52. [Google Scholar] [CrossRef] [Green Version]
- Tian, J.; Hao, L.; Chandra, P.; Jones, D.P.; Willams, I.R.; Gewirtz, A.T.; Ziegler, T.R. Dietary glutamine and oral antibiotics each improve indexes of gut barrier function in rat short bowel syndrome. Am. J. Physiol. Liver Physiol. 2009, 296, G348–G355. [Google Scholar] [CrossRef]
- Fan, J.; Li, G.; Wu, L.; Tao, S.; Wang, W.; Sheng, Z.; Meng, Q. Parenteral glutamine supplementation in combination with enteral nutrition improves intestinal immunity in septic rats. Nutrition 2015, 31, 766–774. [Google Scholar] [CrossRef]
- Wu, M.; Xiao, H.; Liu, G.; Chen, S.; Tan, B.; Ren, W.; Bazer, F.W.; Wu, G.; Yin, Y. Glutamine promotes intestinal SIgA secretion through intestinal microbiota and IL-13. Mol. Nutr. Food Res. 2016, 60, 1637–1648. [Google Scholar] [CrossRef]
- Dphil, P.Y.; Calder, P.C. Glutamine requirement of proliferating T lymphocytes. Nutrition 1997, 13, 646–651. [Google Scholar]
- Coleman, J.W. Nitric oxide in immunity and inflammation. Int. Immunopharmacol. 2001, 1, 1397–1406. [Google Scholar] [CrossRef]
- Cristina, M.C.; Hovsepian, E.; Sarnacki, S.H.; Goren, N.B. Salmonella enterica serovar Enteritidis dam mutant induces low NOS-2 and COX-2 expression in macrophages via attenuation of MAPK and NF-κB pathways. Microbes. Infect. 2008, 10, 1431–1439. [Google Scholar] [CrossRef] [PubMed]
- Meng, Q.; Choudry, H.; Souba, W.; Karinch, A.; Huang, J.; Lin, C.; Vary, T.; Pan, M. Regulation of Amino Acid Arginine Transport by Lipopolysaccharide and Nitric Oxide in Intestinal Epithelial IEC-6 Cells. J. Gastrointest. Surg. 2005, 9, 1276–1285. [Google Scholar] [CrossRef] [PubMed]
- Xiao, Y.; Liu, F.; Zhang, Z.; Tang, J. Gut-Colonizing Bacteria Promote Celegans Innate Immunity by Producing Nitric Oxide. Cell Rep. 2016, 15, 1123. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Alican, I.; Kubes, P. A critical role for nitric oxide in intestinal barrier function and dysfunction. Am. J. Physiol. Liver Physiol. 1996, 270, G225–G237. [Google Scholar] [CrossRef]
- Wu, G. Intestinal Mucosal Amino Acid Catabolism. J. Nutr. 1998, 128, 1249–1252. [Google Scholar] [CrossRef] [Green Version]
- Johansen, F.-E.; Schjerven, H.; Norderhaug, I.N.; Brandtzaeg, P. Regulation of the formation and external transport of secretory immunoglobulins. Crit. Rev. Immunol. 1999, 19, 28. [Google Scholar] [CrossRef]
- Kaetzel, C.S.; Bruno, M.E.C. Epithelial Transport of IgA by the Polymeric Immunoglobulin Receptor. In Mucosal Immune De-fense: Immunoglobulin A; Kaetzel, C.S., Ed.; Springer: New York, NY, USA, 2007; pp. 43–89. [Google Scholar]
- Bruno, M.E.C.; Frantz, A.L.; Rogier, E.W.; Johansen, F.E.; Kaetzel, C.S. Regulation of the polymeric immunoglobulin receptor by the classical and alternative NF-κB pathways in intestinal epithelial cells. Mucosal Immunol. 2011, 4, 468–478. [Google Scholar] [CrossRef] [Green Version]
- Chorny, A.; Puga, I.; Cerutti, A. Innate Signaling Networks in Mucosal IgA Class Switching. Adv. Immunol. 2010, 107, 31–69. [Google Scholar] [CrossRef] [Green Version]
- Sugiarto, H.; Yu, P.L. Avian Antimicrobial Peptides: The Defense Role of Beta-defensins. Biochem. Biophys. Res. Commun. 2004, 323, 721–727. [Google Scholar] [CrossRef]
- van Dijk, A.; Veldhuizen, E.J.; Haagsman, H.P. Avian defensins. Vet. Immunol. Immunop. 2008, 124, 1–18. [Google Scholar] [CrossRef]
- Ma, D.; Zhang, M.; Zhang, K.; Liu, X.; Han, Z.; Shao, Y.; Liu, S. Identification of three novel avian beta-defensins from goose and their significance in the pathogenesis of Salmonella. Mol. Immunol. 2013, 56, 521–529. [Google Scholar] [CrossRef] [PubMed]
- Wang, R.Q.; Ma, D.Y.; Lin, L.J.; Zhou, C.Y.; Han, Z.X.; Shao, Y.H.; Liao, W.Y.; Liu, S.W. Identification and characterization of an avian-defensin orthologue, avian-defensin 9, from quails. Appl. Micro. Biot. 2010, 87, 1395–1405. [Google Scholar] [CrossRef]
- Milona, P.; Townes, C.L.; Bevan, R.M.; Hall, J. The chicken host peptides, gallinacins 4, 7, and 9 have antimicrobial activity against Salmonella serovars. Biochem. Biophys. Res. Commun. 2007, 356, 169–174. [Google Scholar] [CrossRef] [PubMed]
- Kramer, J.; Malek, M.; Lamont, S.J. Association of Twelve Candidate Gene Polymorphisms And response to Challenge with Salmonella Enteritidis in Poultry. Anim. Genet. 2003, 34, 339–348. [Google Scholar] [CrossRef] [Green Version]
- Hasenstein, J.R.; Lamont, S.J. Chicken Gallinacin Gene Cluster Associated with Salmonella Response in Advanced Intercross Line. Avian Dis. 2007, 51, 561–567. [Google Scholar] [CrossRef]
- Hasenstein, J.R.; Zhang, G.; Lamont, S.J. Analyses of Five Gallinacin Genes and the Salmonella Enterica Serovar Enteritidis Re-sponse in Poultry. Infect. Immun. 2006, 74, 3375–3380. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Anastasiadou, M.; Avdi, M.; Michailidis, G. Expression of avian β-defensins and Toll-like receptor genes in the rooster epidi-dymis during growth and Salmonella infection. Anim. Reprod. Sci. 2013, 140, 224–231. [Google Scholar] [CrossRef]
- Pérez-Bárcena, J.; Regueiro, V.; Marsé, P.; Raurich, J.M.; Rodríguez, A.; Ibánez, J.; Mateos, A.G.D.L.; Bengoechea, J.A. Glu-tamine as a modulator of the immune system of critical care patients: Effect on Toll-like receptor expression. Nutrition 2008, 24, 522–527. [Google Scholar] [CrossRef]
- Audisio, M.C.; Oliver, G.; Apella, M.C. Protective Effect of Enterococcus faecium J96, a Potential Probiotic Strain, on Chicks Infected with Salmonella Pullorum. J. Food Prot. 2000, 63, 1333–1337. [Google Scholar] [CrossRef]
- Bartchewsky, W.; Martini, M.R.; Squassoni, A.C.; Alvarez, M.C.; Ladeira, M.S.; Salvatore, D.M.; Trevisan, M.A.; Pedrazzoli, J.; Ribeiro, M.L. Effects of Helicobacter pylori Infection on the Expressions of Bax and Bcl-2 in Patients with Chronic Gastritis and Gastric Cancer. Am. J. Dig. Dis. 2009, 55, 111–116. [Google Scholar] [CrossRef]
- Chang, W.-K.; Yang, K.D.; Chuang, H.; Jan, J.-T.; Shaio, M.-F. Glutamine Protects Activated Human T Cells from Apoptosis by Up-Regulating Glutathione and Bcl-2 Levels. Clin. Immunol. 2002, 104, 151–160. [Google Scholar] [CrossRef] [PubMed]

{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Nutrient Ingredients (g/kg) | 1–21 days |
|---|---|
| Corn ground | 428 |
| Soybean meal, toasted | 365 |
| Wheat | 130 |
| Soybean oil | 17 |
| Corn gluten meal | 20 |
| Canola meal | 0 |
| Na chloride | 2.3 |
| Dicalcium phosphate | 15 |
| Na bicarbonate | 2.4 |
| Ca carbonate | 10.8 |
| DL-Methionine | 2.7 |
| L-Lysine·HCl | 2.2 |
| Premix a | 2 |
| Multi-enzyme | 0.3 |
| Phytase | 0.3 |
| Bentonite | 0.0 |
| Prebiotics | 2 |
| Total | 1000 |
| Calculation of nutrients (g/kg) | |
| Apparent metabolism energy (MJ/kg) | 12.5 |
| Crude protein | 222 |
| Calcium | 9.7 |
| Available phosphorus | 4.7 |
| Lysine | 13.8 |
| Methionine | 6.0 |
| Methionine + cysteine | 8.1 |
| Primer | Sequence (5′→3′) | Length |
|---|---|---|
| pIgR | GGATCTGGAAGCCAGCAAT | 123 bp |
| GAGCCAGAGCTTTGCTCAGA | ||
| AvBD1 | GGATGCACGCTGTTCTTGGT | 100 bp |
| TCCGCATGGTTTACGTCTGTC | ||
| AvBD5 | AGCCGATGGTATTCCTGATGG | 107 bp |
| TGGTGATTGTTGCCTCTGGTG | ||
| AvBD14 | TGCCGAAGATTAAGGGCAA | 83 bp |
| GCTAGTCCATGGTAGVAGGT | ||
| Bcl-2 | ATCCTCGCCTCCTTCGAGTT | 191 bp |
| ATCGCATCCTTCGTTGTCCT | ||
| Bax | GTGCTGGCATGGGACATAGCTA | 119 bp |
| TGGAGTAGACCTTGCGGATAC | ||
| Bak | ACCCGGAGATCATGGAGA | 209 bp |
| GATGCCTTGCTGGTAGACG | ||
| GAPDH | GTGGTGGCCATCAATGATCC | 230 bp |
| ACTTGTGATCAATGGGCACG |
| Item 2 | Treatment 1 | |||
|---|---|---|---|---|
| CON | SCC | Gln1 | Gln2 | |
| 4 d | ||||
| ADFI (g/bird/d) | 20.17 ± 0.12 | 20.03 ± 0.23 | 20.54 ± 1.01 | 20.12 ± 0.81 |
| ADG (g/bird/d) | 15.40 ± 0.14 | 14.84 ± 0.14 | 15.33 ± 0.62 | 15.48 ± 0.51 |
| FCR (g/g) | 1.31 ± 0.03 | 1.35 ± 0.09 | 1.34 ± 0.10 | 1.30 ± 0.07 |
| 7 d | ||||
| ADFI (g/bird/d) | 27.48 ± 0.16 a | 28.29 ± 0.13 b | 27.64 ± 0.14 a | 27.24 ± 0.08 a |
| ADG (g/bird/d) | 19.77 ± 0.14 b | 18.86 ± 0.10 a | 19.33 ± 0.05 b | 19.74 ± 0.13 b |
| FCR (g/g) | 1.39 ± 0.01 a | 1.50 ± 0.11 b | 1.43 ± 0.05 a | 1.38 ± 0.09 a |
| 14 d | ||||
| ADFI (g/bird/d) | 31.71 ± 1.29 | 32.73 ± 0.89 | 32.33 ± 0.52 | 30.20 ± 0.91 |
| ADG (g/bird/d) | 21.57 ± 0.64 | 21.39 ± 0.80 | 21.22 ± 0.77 | 21.57 ± 0.74 |
| FCR (g/g) | 1.47 ± 0.09 | 1.53 ± 0.06 | 1.50 ± 0.08 | 1.45 ± 0.08 |
| 21 d | ||||
| ADFI (g/bird/d) | 40.65 ± 0.72 | 38.50 ± 0.48 | 44.54 ± 0.81 | 43.52 ± 0.27 |
| ADG (g/bird/d) | 24.64 ± 0.58 | 21.88 ± 0.76 | 26.20 ± 0.47 | 25.90 ± 0.61 |
| FCR (g/g) | 1.65 ± 0.08 | 1.76 ± 0.06 | 1.70 ± 0.09 | 1.68 ± 0.03 |
| Items 2 | Treatment 1 | |||
|---|---|---|---|---|
| CON | SCC | Gln1 | Gln2 | |
| 4 days | ||||
| LZM (U/mL) | 112.92 ± 20.42 a | 187.28 ± 19.87 b | 135.37 ± 20.86 a | 120.14 ± 18.24 a |
| IgG (g/L) | 3.28 ± 0.07 b | 1.33 ± 0.12 a | 3.04 ± 0.13 ab | 3.15 ± 0.14 b |
| IgM (g/L) | 0.63 ± 0.19 b | 0.36 ± 0.10 a | 0.60 ± 0.09 b | 0.63 ± 0.17 b |
| IgA (g/L) | 0.65 ± 0.16 b | 0.20 ± 0.15 a | 0.58 ± 0.14 b | 0.62 ± 0.14 b |
| 7 days | ||||
| LZM (U/mL) | 130.31 ± 20.14 a | 202.86 ± 18.91 b | 150.32 ± 21.30 a | 138.34 ± 20.41 a |
| IgG (g/L) | 4.41 ± 0.10 b | 2.39 ± 0.13 a | 4.32 ± 0.09 b | 4.35 ± 0.11 b |
| IgM (g/L) | 0.70 ± 0.11 b | 0.42 ± 0.18 a | 0.63 ± 0.17 b | 0.68 ± 0.15 b |
| IgA (g/L) | 0.75 ± 0.12 b | 0.46 ± 0.18 a | 0.66 ± 0.17 b | 0.72 ± 0.13 b |
| 14 days | ||||
| LZM (U/mL) | 142.34 ± 21.03 a | 214.18 ± 20.41 b | 168.32 ± 24.60 a | 147.31 ± 20.72 a |
| IgG (g/L) | 5.05 ± 0.19 b | 2.53 ± 0.15 a | 4.86 ± 0.16 b | 4.99 ± 0.09 b |
| IgM (g/L) | 0.84 ± 0.13 b | 0.48 ± 0.09 a | 0.75 ± 0.15 b | 0.80 ± 0.18 b |
| IgA (g/L) | 0.80 ± 0.15 b | 0.49 ± 0.10 a | 0.74 ± 0.14 b | 0.79 ± 0.11 b |
| 21 days | ||||
| LZM (U/mL) | 158.63 ± 24.62 a | 235.85 ± 19.68 b | 178.37 ± 29.67 a | 160.31 ± 30.52 a |
| IgG (g/L) | 5.84 ± 0.20 b | 4.97 ± 0.11 a | 5.53 ± 0.18 b | 5.68 ± 0.17 b |
| IgM (g/L) | 0.97 ± 0.08 b | 0.75 ± 0.13 a | 0.88 ± 0.18 b | 0.95 ± 0.17 b |
| IgA (g/L) | 0.85 ± 0.11 b | 0.51 ± 0.13 a | 0.79 ± 0.12 b | 0.83 ± 0.14 b |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wu, Q.; Wang, C.; Liao, J.; Hu, N.; Cheng, B.; Ma, Y.; Wang, Y. Effects of Dietary Supplementation with Glutamine on the Immunity and Intestinal Barrier Gene Expression in Broiler Chickens Infected with Salmonella Enteritidis. Animals 2022, 12, 2168. https://doi.org/10.3390/ani12172168
Wu Q, Wang C, Liao J, Hu N, Cheng B, Ma Y, Wang Y. Effects of Dietary Supplementation with Glutamine on the Immunity and Intestinal Barrier Gene Expression in Broiler Chickens Infected with Salmonella Enteritidis. Animals. 2022; 12(17):2168. https://doi.org/10.3390/ani12172168
Chicago/Turabian StyleWu, Qiujue, Cong Wang, Jiahui Liao, Naizhi Hu, Binyao Cheng, Yan Ma, and Yuqin Wang. 2022. "Effects of Dietary Supplementation with Glutamine on the Immunity and Intestinal Barrier Gene Expression in Broiler Chickens Infected with Salmonella Enteritidis" Animals 12, no. 17: 2168. https://doi.org/10.3390/ani12172168
APA StyleWu, Q., Wang, C., Liao, J., Hu, N., Cheng, B., Ma, Y., & Wang, Y. (2022). Effects of Dietary Supplementation with Glutamine on the Immunity and Intestinal Barrier Gene Expression in Broiler Chickens Infected with Salmonella Enteritidis. Animals, 12(17), 2168. https://doi.org/10.3390/ani12172168