
Salmonella Shedding in Slaughter Pigs and the Use of Esterified Formic Acid in the Drinking Water as a Potential Abattoir-Based Mitigation Measure

 , and
, and
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Study 1 Prevalence Survey and Salmonella Characterization
2.1.1. Herd and Animal Selection and Sample Collection
2.1.2. Salmonella Isolation, Serotyping and Antimicrobial Susceptibility Testing
2.2. Study 2 Assessment of the Efficacy of Esterified Formic Acid in Abattoir Drinking Water
2.2.1. Farm and Animal Selection and Sample Collection
2.2.2. Organic Acids
2.2.3. Salmonella Isolation
2.3. Statistical Analyses
3. Results
3.1. Study 1 Prevalence Survey and Salmonella Characterization
3.1.1. Prevalence of Salmonella Shedding
3.1.2. Antimicrobial Susceptibility Testing
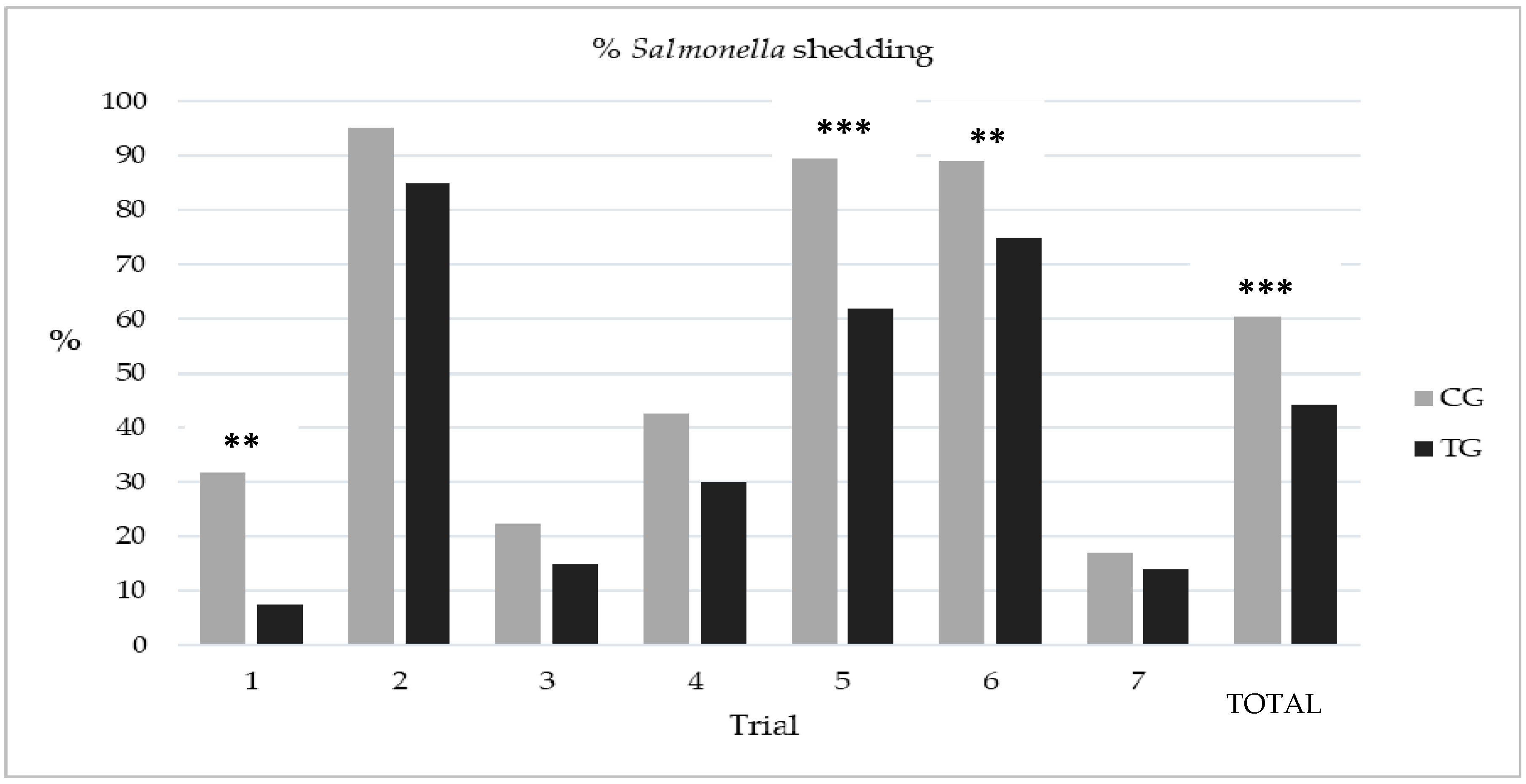
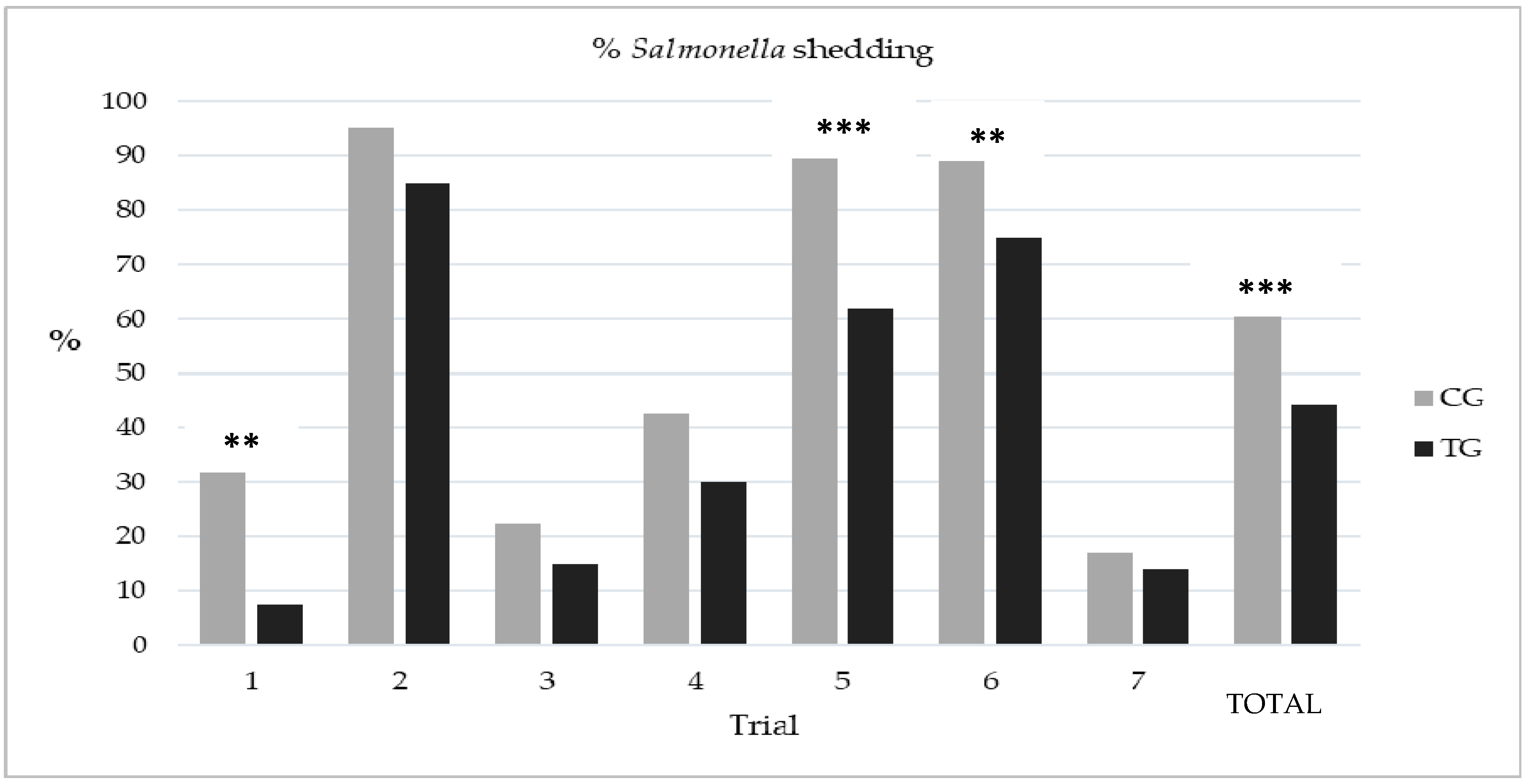
3.2. Study 2 Assessment of the Efficacy of Esterified Formic Acid in Abattoir Drinking Water
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- European Food Safety Authority (EFSA) and European Centre for Disease Prevention and Control (ECDC). The European Union One Health 2020 Zoonoses Report. EFSA J. 2021, 19, 6971. [Google Scholar]
- Correia-Gomes, C.; Leonard, F.; Graham, D. Description of control programmes for Salmonella in pigs in Europe. Progress to date? J. Food Saf. 2021, 41, e12916. [Google Scholar] [CrossRef]
- Bonardi, S. Salmonella in the pork production chain and its impact on human health in the European Union. Epidemiol. Infect. 2017, 145, 1513–1526. [Google Scholar] [CrossRef] [Green Version]
- European Food Safety Authority (EFSA) and European Centre for Disease Prevention and Control (ECDC). Report of the Task Force on Zoonoses Data Collection on the analysis of the baseline survey on the prevalence of Salmonella in slaughter pigs, Part A. EFSA J. 2008, 135, 1–111. [Google Scholar]
- Hurd, H.S.; McKean, J.D.; Griffith, R.D.; Rostagno, M.H. Estimation of the Salmonella enterica prevalence in finishing swine. Epidemiol. Infect. 2004, 132, 127–135. [Google Scholar] [CrossRef]
- European Food Safety Authority (EFSA) and European Centre for Disease Prevention and Control (ECDC). Analysis of the baseline survey on the prevalence of Salmonella in holdings with breeding pigs, in the EU, 2008, Part A: Salmonella prevalence estimates. EFSA J. 2009, 7, 93. [Google Scholar]
- Vico, J.P.; Rol, I.; Garrido, V.; San Román, B.; Grilló, M.J.; Mainar-Jaime, R.C. Salmonellosis in finishing pigs in Spain: Prevalence, antimicrobial agent susceptibilities, and risk factor analysis. J. Food Prot. 2011, 74, 1070–1078. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Massacci, F.R.; Morelli, A.; Cucco, L.; Castinel, A.; Ortenzi, R.; Tofani, S.; Pezzotti, G.; Estellé, J.; Paniccià, M.; Magistrali, C.F. Transport to the Slaughterhouse Affects the Salmonella Shedding and Modifies the Fecal Microbiota of Finishing Pigs. Animals 2020, 10, 676. [Google Scholar] [CrossRef]
- Hurd, H.S.; McKean, J.D.; Griffith, R.W.; Wesley, I.V.; Rostagno, M.H. Salmonella enterica infections in market swine with and without transport and holding. Appl. Environ. Microbiol. 2002, 68, 2376–2381. [Google Scholar] [CrossRef] [Green Version]
- Beloeil, P.A.; Chauvin, C.; Proux, K.; Madec, F.; Fravalo, P.; Alioum, A. Impact of the Salmonella status of market-age pigs and the pre-slaughter process on Salmonella caecal contamination at slaughter. Vet. Res. 2004, 35, 513–530. [Google Scholar] [CrossRef] [Green Version]
- Casanova-Higes, A.; Andrés-Barranco, S.; Mainar-Jaime, R.C. Influence of On-farm pig Salmonella status on Salmonella Shedding at Slaughter. Zoonoses Public Health 2017, 64, 328–336. [Google Scholar] [CrossRef] [PubMed]
- McDowell, S.W.; Porter, R.; Madden, R.; Cooper, B.; Neill, S.D. Salmonella in slaughter pigs in Northern Ireland: Prevalence and use of statistical modelling to investigate sample and abattoir effects. Int. J. Food Microbiol. 2007, 118, 116–125. [Google Scholar] [CrossRef] [PubMed]
- Powell, L.F.; Cheney, T.E.A.; Williamson, S.; Guy, E.; Smith, R.P.; Davies, R.H. A prevalence study of Salmonella spp., Yersinia spp., Toxoplasma gondii and porcine reproductive and respiratory syndrome virus in UK pigs at slaughter. Epidemiol. Infect. 2016, 44, 1538–1549. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Martelli, F.; Oastler, C.; Barker, A.; Jackson, G.; Smith, R.P.; Davies, R. Abattoir-based study of Salmonella prevalence in pigs at slaughter in Great Britain. Epidemiol. Infect. 2021, 149, e218. [Google Scholar] [CrossRef]
- Rostagno, M.H.; Hurd, H.S.; McKean, J.D.; Ziemer, C.J.; Gailey, J.K.; Leite, R.C. Preslaughter holding environment in pork plants is highly contaminated with Salmonella enterica. Appl. Environ. Microbiol. 2003, 69, 4489–4494. [Google Scholar] [CrossRef] [Green Version]
- Scherer, K.; Szabo, I.; Rösler, U.; Appel, B.; Hensel, A.; Nöckler, K. Time course of infection with Salmonella Typhimurium and its influence on fecal shedding, distribution in inner organs, and antibody response in fattening pigs. J. Food Prot. 2008, 71, 699–705. [Google Scholar] [CrossRef]
- Pesciaroli, M.; Cucco, L.; De Luca, S.; Massacci, F.R.; Maresca, C.; Medici, L.; Paniccià, M.; Scoccia, E.; Staffolani, M.; Pezzotti, G.; et al. Association between pigs with high caecal Salmonella loads and carcass contamination. Int. J. Food Microbiol. 2017, 242, 82–86. [Google Scholar] [CrossRef]
- Cantini. Compositions Containing C1 to C7 Organic Acid Monoglycerides and Glycerol, Their Preparation and Use as Antibacterials and Anti-Mould Agents. 2015. Available online: https://patents.google.com/patent/WO2010106488A2/en (accessed on 21 April 2022).
- Gomez-Osorio, L.M.; Yepes-Medina, V.; Ballou, A.; Parini, M.; Angel, R. Short and Medium Chain Fatty Acids and Their Derivatives as a Natural Strategy in the Control of Necrotic Enteritis and Microbial Homeostasis in Broiler Chickens. Front. Vet. Sci. 2021, 8, 773372. [Google Scholar] [CrossRef]
- Tennant, S.M.; Diallo, S.; Levy, H.; Livio, S.; Sow, S.O.; Tapia, M.; Fields, P.I.; Mikoleit, M.; Tamboura, B.; Kotloff, K.L.; et al. Identification by PCR of non-typhoidal Salmonella enterica serovars associated with invasive infections among febrile patients in Mali. PLoS Negl. Trop. Dis. 2010, 4, e621. [Google Scholar] [CrossRef] [Green Version]
- Barco, L.; Lettini, A.A.; Ramon, E.; Longo, A.; Saccardin, C.; Pozza, M.C.; Ricci, A. A rapid and sensitive method to identify and differentiate Salmonella enterica serotype Typhimurium and Salmonella enterica serotype 4,[5],12:i:- by combining traditional serotyping and multiplex polymerase chain reaction. Foodborne Pathog. Dis. 2011, 8, 741–743. [Google Scholar] [CrossRef] [Green Version]
- European Committee on Antimicrobial Susceptibility Testing (EUCAST). Clinical Breakpoints and Dosing of Antibiotics. Version 12.0. Available online: https://www.eucast.org/clinical_breakpoints/ (accessed on 21 April 2022).
- Magiorakos, A.P.; Srinivasan, A.; Carey, R.B.; Carmeli, Y.; Falagas, M.E.; Giske, C.G.; Harbarth, S.; Hindler, J.F.; Kahlmeter, G.; Olsson-Liljequist, B.; et al. Multidrug-resistant, extensively drug-resistant and pandrug-resistant bacteria: An international expert proposal for interim standard definitions for acquired resistance. Clin. Microbiol. Infect. 2012, 18, 268–281. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Luise, D.; Correa, F.; Bosi, P.; Trevisi, P. A Review of the Effect of Formic Acid and Its Salts on the Gastrointestinal Microbiota and Performance of Pigs. Animals 2020, 10, 887. [Google Scholar] [CrossRef] [PubMed]
- Poppe, C.; Ayroud, M.; Ollis, G.; Chirino-Trejo, M.; Smart, N.; Quessy, S.; Michel, P. Trends in antimicrobial resistance of Salmonella isolated from animals, foods of animal origin, and the environment of animal production in Canada, 1994–1997. Microb. Drug. Resist. 2001, 7, 197–212. [Google Scholar] [CrossRef] [PubMed]
- Dohoo, I.R.; Martin, W.; Stryhn, H.E. Veterinary Epidemiologic Research; University of Prince Edward Island: Charlottetown, PE, Canada, 2003; p. 127. [Google Scholar]
- European Food Safety Authority (EFSA) and European Centre for Disease Prevention and Control (ECDC). The European Union One Health 2019 Zoonoses Report. EFSA J. 2021, 19, 6406. [Google Scholar]
- Mainar-Jaime, R.C.; Casanova-Higes, A.; Andrés-Barranco, S.; Vico, J.P. Looking for new approaches for the use of serology in the context of control programmes against pig salmonellosis. Zoonoses Public Health. 2018, 65, e222–e228. [Google Scholar] [CrossRef] [Green Version]
- Boughton, C.; Egan, J.; Kelly, G.; Markey, B.; Leonard, N. Rapid infection of pigs following exposure to environments contaminated with different levels of Salmonella Typhimurium. Foodborne Pathog. Dis. 2007, 4, 33–40. [Google Scholar] [CrossRef]
- Marín, C.; Chinillac, M.C.; Cerdà-Cuéllar, M.; Montoro-Dasi, L.; Sevilla-Navarro, S.; Ayats, T.; Marco-Jimenez, F.; Vega, S. Contamination of pig carcass with Salmonella enterica serovar Typhimurium monophasic variant 1,4[5],12:i:- originates mainly in live animals. Sci. Total Environ. 2020, 703, 134609. [Google Scholar] [CrossRef]
- San Román, B.; Garrido, V.; Sánchez, S.; Martínez-Ballesteros, I.; Garaizar, J.; Mainar-Jaime, R.C.; Migura-Garcia, L.; Grilló, M.J. Relationship between Salmonella infection, shedding and serology in fattening pigs in low-moderate prevalence areas. Zoonoses Public Health 2018, 65, 481–489. [Google Scholar] [CrossRef]
- Mueller-Doblies, D.; Speed, K.C.R.; Kidd, S.; Davies, R.H. Salmonella Typhimurium in livestock in Great Britain—trends observed over a 32-year period. Epidemiol. Infect. 2018, 146, 409–422. [Google Scholar] [CrossRef] [Green Version]
- Teng, K.T.; Martinez-Avilés, M.; Ugarte-Ruiz, M.; Barcena, C.; de la Torre, A.; Lopez, G.; Moreno, M.A.; Dominguez, L.; Alvarez, J. Spatial Trends in Salmonella Infection in Pigs in Spain. Front. Vet. Sci. 2020, 7, 345. [Google Scholar] [CrossRef]
- D′Incau, M.; Salogni, C.; Giovannini, S.; Ruggeri, J.; Scali, F.; Tonni, M.; Formenti, N.; Guarneri, F.; Pasquali, P.; Alborali, G.L. Occurrence of Salmonella Typhimurium and its monophasic variant (4, [5],12:i:-) in healthy and clinically ill pigs in northern Italy. Porcine Health Manag. 2021, 7, 34. [Google Scholar] [CrossRef] [PubMed]
- Mandilara, G.; Sideroglou, T.; Chrysostomou, A.; Rentifis, I.; Papadopoulos, T.; Polemis, M.; Tzani, M.; Tryfinopoulou, K.; Mellou, K. The Rising Burden of Salmonellosis Caused by Monophasic Salmonella Typhimurium (1,4,[5],12:i:-) in Greece and New Food Vehicles. Antibiotics 2021, 10, 185. [Google Scholar] [CrossRef] [PubMed]
- Deane, A.; Murphy, D.; Leonard, F.C.; Byrne, W.; Clegg, T.; Madigan, G.; Griffin, M.; Egan, J.; Prendergast, D.M. Prevalence of Salmonella spp. in slaughter pigs and carcasses in Irish abattoirs and their antimicrobial resistance. Ir. Vet. J. 2022, 75, 4. [Google Scholar] [CrossRef] [PubMed]
- McLure, A.; Shadbolt, C.; Desmarchelier, P.M.; Kirk, M.D.; Glass, K. Source attribution of salmonellosis by time and geography in New South Wales, Australia. BMC Infect. Dis. 2022, 22, 14. [Google Scholar] [CrossRef] [PubMed]
- Ingle, D.J.; Ambrose, R.L.; Baines, S.L.; Duchene, S.; Gonçalves da Silva, A.; Lee, D.Y.J.; Jones, M.; Valcanis, M.; Taiaroa, G.; Ballard, S.A.; et al. Evolutionary dynamics of multidrug resistant Salmonella enterica serovar 4,[5],12:i:- in Australia. Nat. Commun. 2021, 12, 4786. [Google Scholar] [CrossRef]
- Win, A.T.; Supa-Amornkul, S.; Orsi, R.H.; Carey, J.H.; Wolfgang, W.J.; Chaturongakul, S. Sequence Analyses and Phenotypic Characterization Revealed Multidrug Resistant Gene Insertions in the Genomic Region Encompassing Phase 2 Flagellin Encoding fljAB Genes in Monophasic Variant Salmonella enterica Serovar 4,5,12:i:- Isolates From Various Sources in Thailand. Front. Microbiol. 2021, 12, 720604. [Google Scholar]
- Yang, X.; Wu, Q.; Zhang, J.; Huang, J.; Guo, W.; Cai, S. Prevalence and Characterization of Monophasic Salmonella Serovar 1,4,[5],12:i:- of Food Origin in China. PLoS ONE 2015, 10, e0137967. [Google Scholar] [CrossRef] [Green Version]
- Wasyl, D.; Hoszowski, A. Occurrence and characterization of monophasic Salmonella enterica serovar Typhimurium (1,4,[5],12:i:-) of non-human origin in Poland. Foodborne Pathog. Dis. 2012, 9, 1037–1043. [Google Scholar] [CrossRef]
- Andrés-Barranco, S.; Vico, J.P.; Marín, C.M.; Herrera-León, S.; Mainar-Jaime, R.C. Characterization of Salmonella enterica Serovar Typhimurium Isolates from Pigs and Pig Environment-Related Sources and Evidence of New Circulating Monophasic Strains in Spain. J. Food Prot. 2016, 79, 407–412. [Google Scholar] [CrossRef]
- Wang, Z.; Xu, H.; Tang, Y.; Li, Q.; Jiao, X. A Multidrug-resistant Monophasic Salmonella Typhimurium Co-harboring mcr-1, fosA3, blaCTX-M-14 in a Transferable IncHI2 Plasmid from a Healthy Catering Worker in China. Infect. Drug Resist. 2020, 13, 3569–3574. [Google Scholar] [CrossRef]
- Vázquez, X.; García, V.; Fernández, J.; Bances, M.; de Toro, M.; Ladero, V.; Rodicio, R.; Rodicio, M.R. Colistin Resistance in Monophasic Isolates of Salmonella enterica ST34 Collected from Meat-Derived Products in Spain, with or without CMY-2 Co-production. Front. Microbiol. 2022, 12, 735364. [Google Scholar] [CrossRef] [PubMed]
- Qin, X.; Yang, M.; Cai, H.; Liu, Y.; Gorris, L.; Aslam, M.Z.; Jia, K.; Sun, T.; Wang, X.; Dong, Q. Antibiotic Resistance of Salmonella Typhimurium Monophasic Variant 1,4,[5],12:i:-in China: A Systematic Review and Meta-Analysis. Antibiotics 2022, 11, 532. [Google Scholar] [CrossRef] [PubMed]
- European Food Safety Authority (EFSA) and European Centre for Disease Prevention and Control (ECDC). The European Union Summary Report on Antimicrobial Resistance in zoonotic and indicator bacteria from humans, animals and food in 2019–2020. EFSA J. 2022, 20, 7209. [Google Scholar]
- Sevilla, E.; Vico, J.P.; Delgado-Blas, J.F.; González-Zorn, B.; Marín, C.M.; Uruén, C.; Martín-Burriel, I.; Bolea, R.; Mainar-Jaime, R.C. Resistance to colistin and production of extended-spectrum β-lactamases and/or AmpC enzymes in Salmonella isolates collected from healthy pigs in Northwest Spain in two periods: 2008-2009 and 2018. Int. J. Food Microbiol. 2021, 338, 108967. [Google Scholar] [CrossRef] [PubMed]
- Agencia Española de Medicamentos y Productos Sanitarios (AEMPS). Plan Nacional Frente a la Resistencia a los Antibióticos 2019–2021. Available online: http://www.resistenciaantibioticos.es/es/system/files/field/files/pran_2019-2021_0.pdf?file=1&type=node&id=497&force=0 (accessed on 25 April 2022).
- Wang, Y.; Xu, C.; Zhang, R.; Chen, Y.; Shen, Y.; Hu, F.; Liu, D.; Lu, J.; Guo, Y.; Xia, X.; et al. Changes in colistin resistance and mcr-1 abundance in Escherichia coli of animal and human origins following the ban of colistin-positive additives in China: An epidemiological comparative study. Lancet Infect. Dis. 2020, 20, 1161–1171. [Google Scholar] [CrossRef]
- Miguela-Villoldo, P.; Moreno, M.A.; Rodríguez-Lázaro, D.; Gallardo, A.; Hernández, M.; Serrano, T.; Sáez, J.L.; de Frutos, C.; Agüero, M.; Quesada, A.; et al. Longitudinal study of the mcr-1 gene prevalence in Spanish food-producing pigs from 1998 to 2021 and its relationship with the use of polymyxins. Porcine Health Manag. 2022, 8, 12. [Google Scholar] [CrossRef]
- European Food Safety Authority (EFSA) and European Centre for Disease Prevention and Control (ECDC). The European Union summary report on antimicrobial resistance in zoonotic and indicator bacteria from humans, animals and food in 2017. EFSA J. 2019, 17, 5598. [Google Scholar]
- Cameron-Veas, K.; Fraile, L.; Napp, S.; Garrido, V.; Grilló, M.J.; Migura-Garcia, L. Multidrug resistant Salmonella enterica isolated from conventional pig farms using antimicrobial agents in preventative medicine programmes. Vet. J. 2018, 234, 36–42. [Google Scholar] [CrossRef]
- Alessiani, A.; Goffredo, E.; Mancini, M.; Occhiochiuso, G.; Faleo, S.; Didonna, A.; Fischetto, R.; Suglia, F.; De Vito, D.; Stallone, A.; et al. Evaluation of Antimicrobial Resistance in Salmonella Strains Isolated from Food, Animal and Human Samples between 2017 and 2021 in Southern Italy. Microorganisms 2022, 10, 812. [Google Scholar] [CrossRef]
- European Centre for Disease Prevention and Control (ECDC); European Food Safety Authority (EFSA); European Medicines Agency (EMA). Third Joint Inter-Agency Report on Integrated Analysis of Consumption of Antimicrobial Agents and Occurrence of Antimicrobial Resistance in Bacteria from Humans and Food-Producing Animals in the EU/EEA, JIACRA III 2016–2018. 2021. Available online: https://www.ema.europa.eu/en/documents/report/ema/ecdc/efsa-third-joint-report-integrated-analysis-consumption-antimicrobial-agents-occurrence_en.pdf (accessed on 28 April 2022).
- Yin, X.; Dudley, E.G.; Pinto, C.N.; M′ikanatha, N.M. Fluoroquinolone sales in food animals and quinolone resistance in non-typhoidal Salmonella from retail meats: United States, 2009-2018. J. Glob. Antimicrob. Resist. 2022, 29, 163–167. [Google Scholar] [CrossRef]
- European Medicines Agency (EMA); European Surveillance of Veterinary Antimicrobial Consumption (ESVAC). Sales of Veterinary Antimicrobial Agents in 31 European Countries in 2019 and 2020 (EMA/58183/2021). 2021. Available online: https://www.ema.europa.eu/en/documents/report/sales-veterinary-antimicrobial-agents-31-european-countries-2019-2020-trends-2010-2020-eleventh_en.pdf (accessed on 28 April 2022).
- Capoor, M.R.; Nair, D.; Posti, J.; Singhal, S.; Deb, M.; Aggarwal, P.; Pillai, P. Minimum inhibitory concentration of carbapenems and tigecycline against Salmonella spp. J. Med. Microbiol. 2009, 58, 337–341. [Google Scholar] [CrossRef] [PubMed]
- Sun, C.; Cui, M.; Zhang, S.; Wang, H.; Song, L.; Zhang, C.; Zhao, Q.; Liu, D.; Wang, Y.; Shen, J.; et al. Plasmid-mediated tigecycline-resistant gene tet(X4) in Escherichia coli from food-producing animals, China, 2008–2018. Emerg. Microbes Infect. 2019, 8, 1524–1527. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sun, J.; Chen, C.; Cui, C.Y.; Zhang, Y.; Liu, X.; Cui, Z.H.; Ma, X.Y.; Feng, Y.; Fang, L.X.; Lian, X.L.; et al. Plasmid-encoded tet(X) genes that confer high-level tigecycline resistance in Escherichia coli. Nat. Microbiol. 2019, 4, 1457–1464. [Google Scholar] [CrossRef] [PubMed]
- Snary, E.L.; Swart, A.N.; Simons, R.R.; Domingues, A.R.; Vigre, H.; Evers, E.G.; Hald, T.; Hill, A.A. A Quantitative Microbiological Risk Assessment for Salmonella in Pigs for the European Union. Risk Anal. 2016, 36, 437–449. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Canibe, N.; Højberg, O.; Højsgaard, S.; Jensen, B.B. Feed physical form and formic acid addition to the feed affect the gastrointestinal ecology and growth performance of growing pigs. J. Anim. Sci. 2005, 83, 1287–1302. [Google Scholar] [CrossRef] [Green Version]
- Papenbrock, S.; Stemme, K.; Amtsberg, G.; Verspohl, J.; Kamphues, J. Investigations on prophylactic effects of coarse feed structure and/or potassium diformate on the microflora in the digestive tract of weaned piglets experimentally infected with Salmonella Derby. J. Anim. Physiol. Anim. Nutr. 2005, 89, 84–87. [Google Scholar] [CrossRef]
- Creus, E.; Pérez, J.F.; Peralta, B.; Baucells, F.; Mateu, E. Effect of acidified feed on the prevalence of Salmonella in market-age pigs. Zoonoses Public Health 2007, 54, 314–319. [Google Scholar] [CrossRef]
- Lynch, H.; Leonard, F.C.; Walia, K.; Lawlor, P.G.; Duffy, G.; Fanning, S.; Markey, B.K.; Brady, C.; Gardiner, G.E.; Argüello, H. Investigation of in-feed organic acids as a low cost strategy to combat Salmonella in grower pigs. Prev. Vet. Med. 2017, 139, 50–57. [Google Scholar] [CrossRef]
- Walia, K.; Argüello, H.; Lynch, H.; Leonard, F.C.; Grant, J.; Yearsley, D.; Kelly, S.; Duffy, G.; Gardiner, G.E.; Lawlor, P.G. Effect of strategic administration of an encapsulated blend of formic acid, citric acid, and essential oils on Salmonella carriage, seroprevalence, and growth of finishing pigs. Prev. Vet. Med. 2017, 137, 28–35. [Google Scholar] [CrossRef]
- Van der Wolf, P.J.; Wolbers, W.B.; Elbers, A.R.; van der Heijden, H.M.; Koppen, J.M.; Hunneman, W.A.; van Schie, F.W.; Tielen, M.J. Herd level husbandry factors associated with the serological Salmonella prevalence in finishing pig herds in The Netherlands. Vet. Microbiol. 2001, 78, 205–219. [Google Scholar] [CrossRef]
- Boyen, F.; Haesebrouck, F.; Vanparys, A.; Volf, J.; Mahu, M.; Van Immerseel, F.; Rychlik, I.; Dewulf, J.; Ducatelle, R.; Pasmans, F. Coated fatty acids alter virulence properties of Salmonella Typhimurium and decrease intestinal colonization of pigs. Vet. Microbiol. 2008, 132, 319–327. [Google Scholar] [CrossRef] [PubMed]
- Taube, V.A.; Neu, M.E.; Hassan, Y.; Verspohl, J.; Beyerbach, M.; Kamphues, J. Effects of dietary additives (potassium diformate/organic acids) as well as influences of grinding intensity (coarse/fine) of diets for weaned piglets experimentally infected with Salmonella Derby or Escherichia coli. J. Anim. Physiol. Anim. Nutr. 2009, 93, 350–358. [Google Scholar] [CrossRef] [PubMed]
- Visscher, C.F.; Winter, P.; Verspohl, J.; Stratmann-Selke, J.; Upmann, M.; Beyerbach, M.; Kamphues, J. Effects of feed particle size at dietary presence of added organic acids on caecal parameters and the prevalence of Salmonella in fattening pigs on farm and at slaughter. J. Anim. Physiol. Anim. Nutr. 2009, 93, 423–430. [Google Scholar] [CrossRef]
- Argüello, H.; Carvajal, A.; Costillas, S.; Rubio, P. Effect of the addition of organic acids in drinking water or feed during part of the finishing period on the prevalence of Salmonella in finishing pigs. Foodborne Pathog. Dis. 2013, 10, 842–849. [Google Scholar] [CrossRef] [PubMed]
- Casanova-Higes, A.; Andrés-Barranco, S.; Mainar-Jaime, R.C. Effect of the addition of protected sodium butyrate to the feed on Salmonella spp. infection dynamics in fattening pigs. Anim. Feed Sci. Technol. 2017, 231, 12–18. [Google Scholar] [CrossRef] [Green Version]
- Casanova-Higes, A.; Andrés-Barranco, S.; Mainar Jaime, R.C. Use of a New Form of Protected Sodium Butyrate to Control Salmonella Infection in Fattening Pigs. Span. J. Agric. Res. 2019, 16, e05SC02. [Google Scholar] [CrossRef] [Green Version]
- De Busser, E.V.; Dewulf, J.; Nollet, N.; Houf, K.; Schwarzer, K.; De Sadeleer, L.; De Zutter, L.; Maes, D. Effect of organic acids in drinking water during the last 2 weeks prior to slaughter on Salmonella shedding by slaughter pigs and contamination of carcasses. Zoonoses Public Health. 2009, 56, 129–136. [Google Scholar] [CrossRef]
- Martín-Peláez, S.; Costabile, A.; Hoyles, L.; Rastall, R.A.; Gibson, G.R.; La Ragione, R.M.; Woodward, M.J.; Mateu, E.; Martín-Orúe, S.M. Evaluation of the inclusion of a mixture of organic acids or lactulose into the feed of pigs experimentally challenged with Salmonella Typhimurium. Vet. Microbiol. 2010, 142, 337–345. [Google Scholar] [CrossRef]
- De Ridder, L.; Maes, D.; Dewulf, J.; Pasmans, F.; Boyen, F.; Haesebrouck, F.; Méroc, E.; Butaye, P.; Van der Stede, Y. Evaluation of three intervention strategies to reduce the transmission of Salmonella Typhimurium in pigs. Vet. J. 2013, 197, 613–618. [Google Scholar] [CrossRef] [Green Version]
- Yoon, B.K.; Jackman, J.A.; Valle-González, E.R.; Cho, N.J. Antibacterial Free Fatty Acids and Monoglycerides: Biological Activities, Experimental Testing, and Therapeutic Applications. Int. J. Mol. Sci. 2018, 19, 1114. [Google Scholar] [CrossRef] [Green Version]
- Bonardi, S.; Alpigiani, I.; Bruini, I.; Barilli, E.; Brindani, F.; Morganti, M.; Cavallini, P.; Bolzoni, L.; Pongolini, S. Detection of Salmonella enterica in pigs at slaughter and comparison with human isolates in Italy. Int. J. Food Microbiol. 2016, 218, 44–50. [Google Scholar] [CrossRef] [PubMed]
- De la Cruz, M.L.; Conrado, I.; Nault, A.; Perez, A.; Dominguez, L.; Alvarez, J. Vaccination as a control strategy against Salmonella infection in pigs: A systematic review and meta-analysis of the literature. Res. Vet. Sci. 2017, 114, 86–94. [Google Scholar] [CrossRef] [PubMed]
- Cargnel, M.; Maes, D.; Peeters, L.; Dispas, M. Combining quantitative and qualitative approaches to determine viability of a potential Salmonella Typhimurium vaccination program in pigs in Belgium. Prev. Vet. Med. 2020, 184, 105132. [Google Scholar] [CrossRef] [PubMed]

{kind=link}
| Farm ID | N | No. + (%) | Serotypes Involved (No. of Strains) |
|---|---|---|---|
| 1 | 43 | 0 (0.00) | - |
| 2 | 46 | 19 (41.30) | S. Typhimurium (3), mST (16) |
| 3 | 45 | 1 (2.22) | other (1) |
| 4 | 49 | 9 (18.37) | S. Typhimurium (1), mST (8) |
| 5 | 46 | 35 (76.09) | S. Typhimurium (20), mST (5), other (10) |
| 6 | 48 | 27 (56.25) | S. Typhimurium (7), mST (18), other (2) |
| 7 | 25 | 18 (72.00) | other (18) |
| 8 | 48 | 33 (68.75) | S. Typhimurium (11), mST (5), other (17) |
| 9 | 46 | 15 (32.61) | S. Typhimurium (1), mST (7), other (7) |
| 10 | 45 | 39 (86.67) | S. Typhimurium (4), mST (34), other (1) |
| 11 | 50 | 4 (8.00) | mST (2), other (2) |
| 12 | 51 | 1 (1.96) | S. Typhimurium (1) |
| 13 | 44 | 8 (18.18) | mST (8) |
| 14 | 48 | 9 (18.75) | other (9) |
| 15 | 48 | 7 (14.58) | mST (7) |
| 16 | 47 | 5 (11.36) | other (5) |
| 17 | 46 | 4 (8.70) | other (4) |
| 18 | 39 | 1 (2.56) | other (1) |
| 19 | 34 | 27 (79.41) | mST (25), other (2) |
| 20 | 46 | 1 (2.17) | other (1) |
| 21 | 44 | 2 (4.55) | S. Typhimurium (1), other (1) |
| 22 | 44 | 19 (43.18) | S. Typhimurium (3), mST (2), other (14) |
| 23 | 44 | 0 (0.00) | - |
| 24 | 42 | 8 (19.05) | S. Typhimurium (3), other (5) |
| All | 1068 | 292 (27.34) | S. Typhimurium (55), mST (137), other (100) |
| Antimicrobial Class | Antimicrobial Agent * | No. ST (%) n = 20 | No. mST (%) n = 28 | No. “Other” Serotypes (%) n = 32 | Farm ID |
|---|---|---|---|---|---|
| Penicillins | AMP | 14 (70.0) | 26 (92.9) | 19 (59.4) | 2, 4, 5, 6, 8, 9, 10, 11, 12, 13, 15, 16, 17, 19, 20, 21, 22, 24 |
| AMC | 0 (0.0) | 3 (10.7) | 3 (9.4) | 11, 13 | |
| TZP | 0 (0.0) | 2 (7.1) | 0 (0.0) | 11 | |
| Cephalosporins | CXM | 1 (5.0) | 1 (3.6) | 3 (9.4) | 13, 22 |
| FOX | 0 (0.0) | 0 (0.0) | 0 (0.0) | - | |
| CTX | 0 (0.0) | 1 (3.6) | 3 (9.4) | 13, 22 | |
| CAZ | 0 (0.0) | 0 (0.0) | 3 (9.4) | 22 | |
| FEP | 0 (0.0) | 0 (0.0) | 0 (0.0) | - | |
| Carbapenems | ETP | 0 (0.0) | 0 (0.0) | 0 (0.0) | - |
| IPM | 0 (0.0) | 0 (0.0) | 0 (0.0) | - | |
| Polymyxins | CST | 0 (0.0) | 0 (0.0) | 0 (0.0) | - |
| Aminoglycosides | AMK | 0 (0.0) | 0 (0.0) | 0 (0.0) | - |
| GEN | 0 (0.0) | 3 (10.7) | 2 (6.3) | 8, 11, 13 | |
| Tetracyclines | TGC | 4 (20.0) | 4 (14.3) | 4 (12.5) | 2, 11, 16, 17, 22 |
| Quinolones | NAL | 2 (10.0) | 7 (25.0) | 7 (21.9) | 8, 9, 11, 12, 13, 16, 19, 20, 22, 24 |
| CIP | 0 (0.0) | 0 (0.0) | 1 (3.1) | 16 | |
| Sulphonamides dihydrofolate reductase inhibitors | SXT | 2 (10.0) | 3 (10.7) | 11 (34.4) | 8, 9, 11, 12, 13, 16, 17, 19 |
| Estimated Water Consumption Per Pig | ||||
|---|---|---|---|---|
| <0.9 L | ≥0.9 L * | |||
| No. (%) | No. (%) | Total | ||
| Salmonella shedding | Yes | 96 (76.8) | 29 (23.2) | 125 |
| No | 66 (42.0) | 91 (58.0) | 157 | |
| OR = 4.56 (95% CI: 2.70–7.69; p < 0.0001) | ||||
| Logistic Regression Parameters | |||||||
|---|---|---|---|---|---|---|---|
| Variable | N | No. + (%) | β | SE (β) | p | OR | 95% CI OR |
| Group | |||||||
| Treatment 1 | 282 | 125 (44.3) | 1 | ||||
| Control | 280 | 170 (60.7) | 1.01 | 0.21 | <0.001 | 2.75 | 1.80–4.21 |
| Season | |||||||
| Spring 1 | 240 | 156 (65) | 1 | ||||
| Summer | 80 | 31 (38.7) | −3.32 | 2.06 | 0.107 | 0.036 | 0.0006–2.05 |
| Autumn | 242 | 108 (44.6) | −2.45 | 1.42 | 0.085 | 0.086 | 0.005–1.39 |
| Lairage time | |||||||
| <15 h | 320 | 171 (53.4) | |||||
| ≥15 h | 242 | 124 (51.2) | 1.94 | 1.50 | 0.195 | 7.01 | 0.36–132 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bernad-Roche, M.; Casanova-Higes, A.; Marín-Alcalá, C.M.; Mainar-Jaime, R.C. Salmonella Shedding in Slaughter Pigs and the Use of Esterified Formic Acid in the Drinking Water as a Potential Abattoir-Based Mitigation Measure. Animals 2022, 12, 1620. https://doi.org/10.3390/ani12131620
Bernad-Roche M, Casanova-Higes A, Marín-Alcalá CM, Mainar-Jaime RC. Salmonella Shedding in Slaughter Pigs and the Use of Esterified Formic Acid in the Drinking Water as a Potential Abattoir-Based Mitigation Measure. Animals. 2022; 12(13):1620. https://doi.org/10.3390/ani12131620
Chicago/Turabian StyleBernad-Roche, María, Alejandro Casanova-Higes, Clara María Marín-Alcalá, and Raúl Carlos Mainar-Jaime. 2022. "Salmonella Shedding in Slaughter Pigs and the Use of Esterified Formic Acid in the Drinking Water as a Potential Abattoir-Based Mitigation Measure" Animals 12, no. 13: 1620. https://doi.org/10.3390/ani12131620
APA StyleBernad-Roche, M., Casanova-Higes, A., Marín-Alcalá, C. M., & Mainar-Jaime, R. C. (2022). Salmonella Shedding in Slaughter Pigs and the Use of Esterified Formic Acid in the Drinking Water as a Potential Abattoir-Based Mitigation Measure. Animals, 12(13), 1620. https://doi.org/10.3390/ani12131620