
Chitosan/Calcium–Alginate Encapsulated Flaxseed Oil on Dairy Cattle Diet: In Vitro Fermentation and Fatty Acid Biohydrogenation

 ,
,  ,
,  ,
,  ,
,  and
and 
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Preparation of Oil-Loaded Chitosan and Calcium Alginate Nanoparticles (NPs)
2.2. Measurement of Particle Size and Zeta Potential
2.3. Encapsulation Efficiency
2.4. Fourier Transform Infrared (FTIR) Characterization
2.5. In Vitro Gas Production Technique
2.6. In Vitro Digestibility
2.7. In Vitro Biohydrogenation
2.7.1. Preparation of Samples for Determination of Fatty Acid Profiles
Fatty Acid Extraction
Methylation Procedure
2.8. Statistical Analysis
3. Results
3.1. Capsulation efficiency
3.2. Zeta Potential
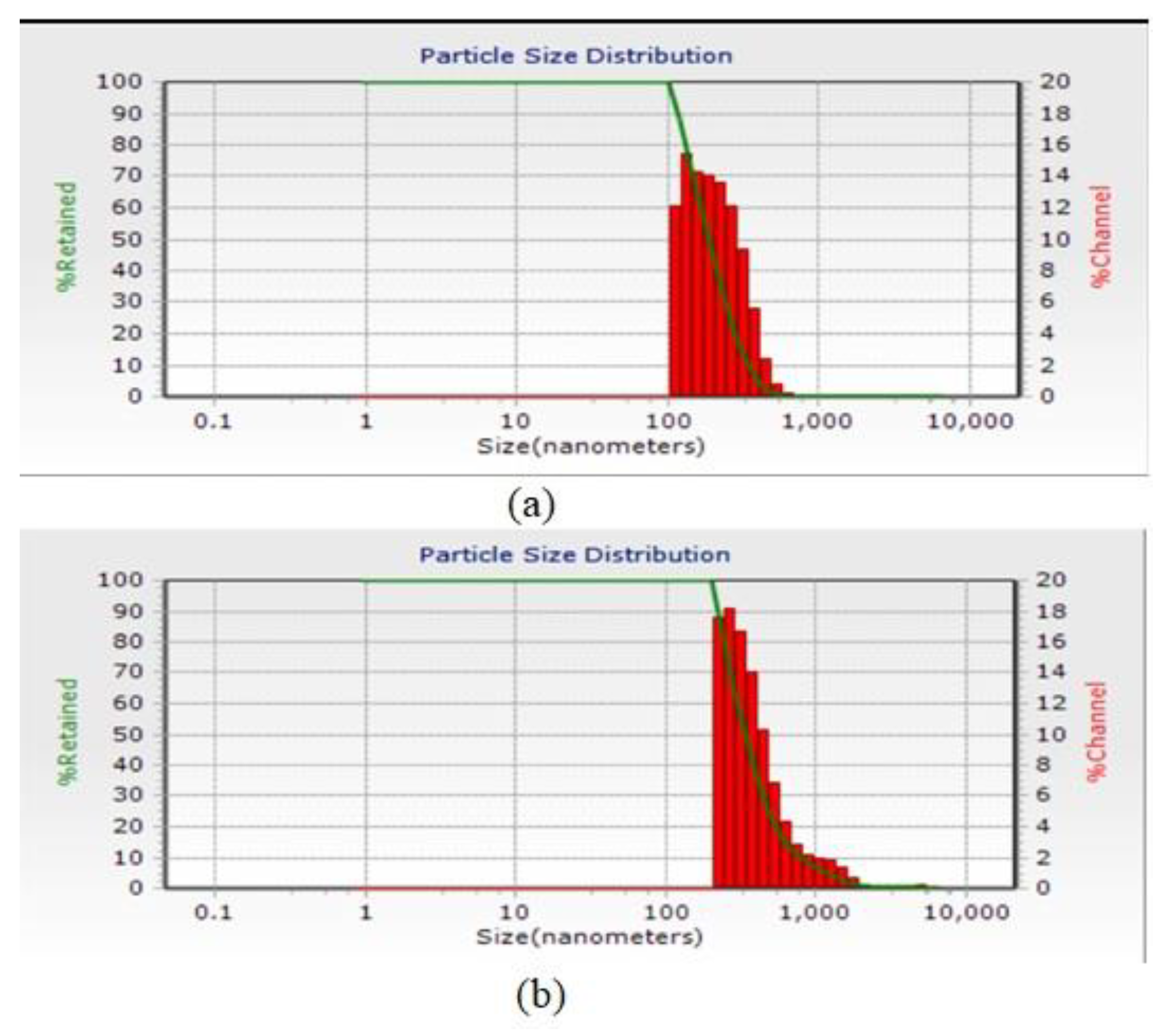
3.3. Particle Size Distribution
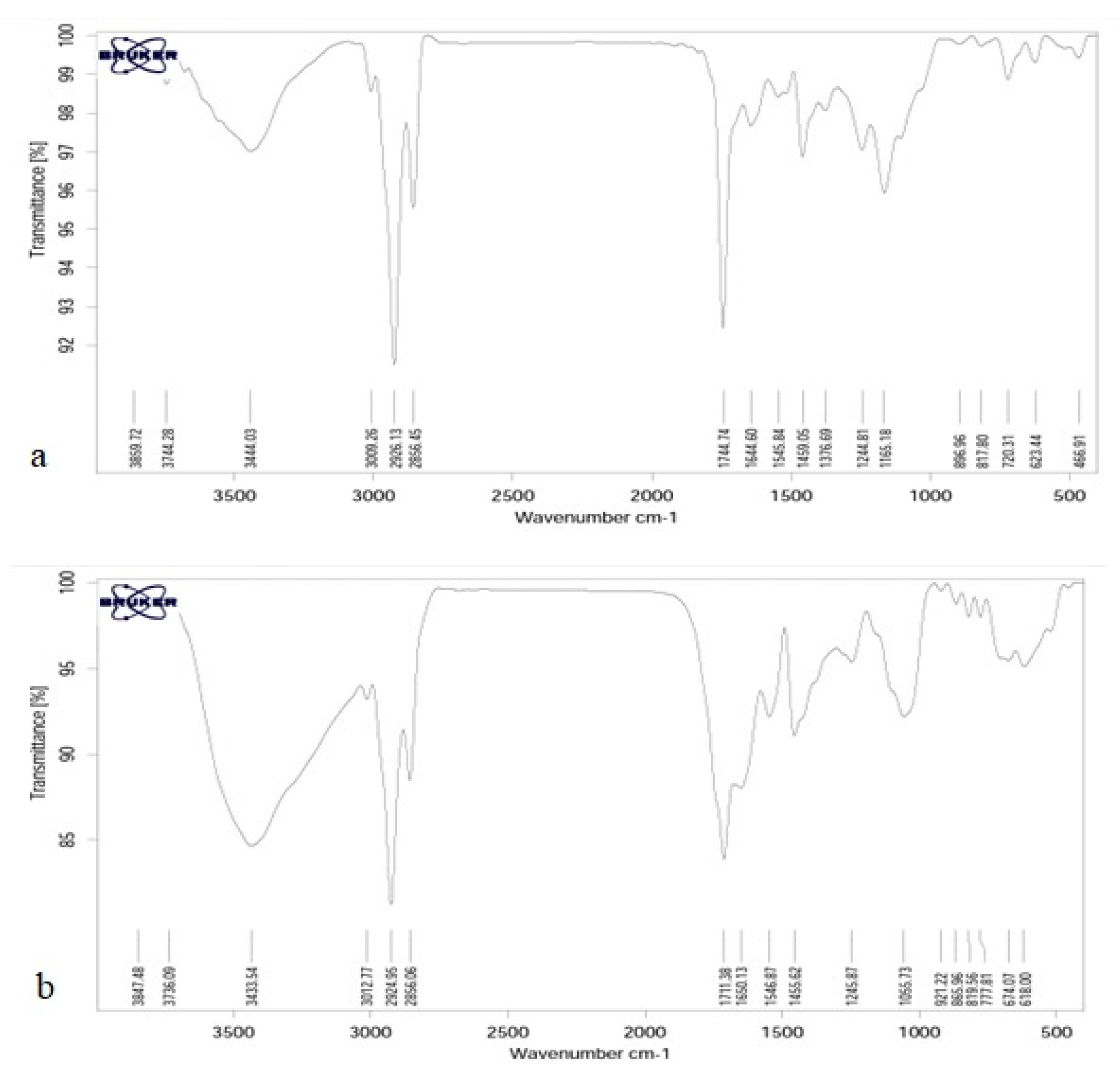
3.4. Fourier Transform Infrared Spectroscopy Test (FTIR)
3.5. Effect of Encapsulation on In Vitro Gas Production
3.6. The Effect of Flaxseed oil Encapsulation on the In Vitro Digestibility of Dairy Cattle Diets
3.7. The Effect of Flaxseed Oil Encapsulation on Biohydrogenation of Fatty Acids in Experimental Treatments
4. Discussion
4.1. Capsulation Efficiency
4.2. Zeta Potential
4.3. Particle Size Distribution
4.4. Effect of Encapsulation on In Vitro Gas Production
4.5. The Effect of Flaxseed Oil Encapsulation on the In Vitro Digestibility of Dairy Cattle Diets
4.6. The Effect of Flaxseed Oil Encapsulation on Biohydrogenation of Fatty Acids in Experimental Treatments
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Kalyesubula, M.; Rosov, A.; Alon, T.; Moallem, U.; Dvir, H. Intravenous Infusions of Glycerol Versus Propylene Glycol for the Regulation of Negative Energy Balance in Sheep: A Randomized Trial. Animals 2019, 9, 731. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sharma, A.; Baddela, V.S.; Becker, F.; Dannenberger, D.; Viergutz, T.; Vanselow, J. Elevated free fatty acids affect bovine granulosa cell function: A molecular cue for compromised reproduction during negative energy balance. J. Endocr. Connect. 2019, 8, 493–505. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cabezas-Garcia, E.H.; Gordon, A.W.; Mulligan, F.J.; Ferris, C.P. Revisiting the Relationships between Fat-to-Protein Ratio in Milk and Energy Balance in Dairy Cows of Different Parities, and at Different Stages of Lactation. Animals 2021, 11, 3256. [Google Scholar] [CrossRef]
- Moallem, U. Invited review: Roles of dietary n-3 fatty acids in performance, milk fat composition, and reproductive and immune systems in dairy cattle. J. Dairy Sci. 2018, 101, 8641–8661. [Google Scholar] [CrossRef] [Green Version]
- Oliveira, R.C.; Pralle, R.S.; de Resende, L.C.; Nova, C.H.P.; Caprarulo, V.; Jendza, J.A.; Troescher, A.; White, H.M. Prepartum supplementation of conjugated linoleic acids (CLA) increased milk energy output and decreased serum fatty acids and β-hydroxybutyrate in early lactation dairy cows. PLoS ONE 2018, 13, e0197733. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bradford, B. Transition cows-How fatty acids affect immunity, production and health. J. Dairy Res. 2020, 87, 4–13. [Google Scholar]
- Williams, S.R.O.; Milner, T.C.; Garner, J.B.; Moate, P.J.; Jacobs, J.L.; Hannah, M.C.; Wales, W.J.; Marett, L.C. Dietary Fat and Betaine Supplements Offered to Lactating Cows Affect Dry Matter Intake, Milk Production and Body Temperature Responses to an Acute Heat Challenge. Animals 2021, 11, 3110. [Google Scholar] [CrossRef] [PubMed]
- Zhao, L.; Huang, Y.; Du, M. Farm animals for studying muscle development and metabolism: Dual purposes for animal production and human health. Anim. Front. 2019, 9, 21–27. [Google Scholar] [CrossRef] [Green Version]
- Jia, S.; Li, X.; He, W.; Wu, G. Protein-Sourced Feedstuffs for Aquatic Animals in Nutrition Researchand Aquaculture. In Recent Advances in Animal Nutrition and Metabolism; Wu, G., Ed.; Springer International Publishing: Cham, Switzerland, 2022; pp. 237–261. [Google Scholar] [CrossRef]
- Ahmad, N.; Khan, M.K.; Imran, M.; Suleman, M.N.; Afzal, S. Bovine Feed Manipulation, Enhancement of Conjugated Linoleic Acid and Its Bioavailability. In Bovine Science-A Key to Sustainable Development; IntechOpen: London, UK, 2019. [Google Scholar]
- Haubold, S.; Kröger-Koch, C.; Tuchscherer, A.; Kanitz, E.; Weitzel, J.M.; Hoeflich, A.; Starke, A.; Tröscher, A.; Sauerwein, H.; Hammon, H.M. Effects of a combined essential fatty acid and conjugated linoleic acid abomasal infusion on metabolic and endocrine traits, including the somatotropic axis, in dairy cows. J. Dairy Sci. 2020, 103, 12069–12082. [Google Scholar] [CrossRef]
- Vogel, L.; Gnott, M.; Kröger-Koch, C.; Dannenberger, D.; Tuchscherer, A.; Tröscher, A.; Kienberger, H.; Rychlik, M.; Starke, A.; Bachmann, L.; et al. Effects of abomasal infusion of essential fatty acids together with conjugated linoleic acid in late and early lactation on performance, milk and body composition, and plasma metabolites in dairy cows. J. Dairy Sci. 2020, 103, 7431–7450. [Google Scholar] [CrossRef]
- Palmquist, D.L. Biohydrogenation then and now. Eur. J. Lipid Sci. Technol. 2007, 109, 737–739. [Google Scholar] [CrossRef]
- Bauman, D.E.; Corl, B.A.; Peterson, D.G. The biology of conjugated linoleic acids in ruminants. Adv. Conjug. Linoleic Acid Res. 2020, 2, 146–173. [Google Scholar]
- Alves, S.P.; Mendonça, S.H.; Silva, J.L.; Bessa, R.J.B. Nannochloropsis oceanica, a novel natural source of rumen-protected eicosapentaenoic acid (EPA) for ruminants. Sci. Rep. 2018, 8, 10269. [Google Scholar] [CrossRef] [PubMed]
- Honkanen, A. Biohydrogenation of Linoleic and Alpha-Linolenic Acid in Rumen Fluid In Vitro. Ph.D. Thesis, University of Helsinki, Helsinki, Finland, 2020. [Google Scholar]
- Dewanckele, L.; Vlaeminck, B.; Hernandez-Sanabria, E.; Ruiz-González, A.; Debruyne, S.; Jeyanathan, J.; Fievez, V. Rumen Biohydrogenation and Microbial Community Changes Upon Early Life Supplementation of 22:6n-3 Enriched Microalgae to Goats. Front. Microbiol. 2018, 9, 573. [Google Scholar] [CrossRef] [Green Version]
- Vasta, V.; Daghio, M.; Cappucci, A.; Buccioni, A.; Serra, A.; Viti, C.; Mele, M. Invited review: Plant polyphenols and rumen microbiota responsible for fatty acid biohydrogenation, fiber digestion, and methane emission: Experimental evidence and methodological approaches. J. Dairy Sci. 2019, 102, 3781–3804. [Google Scholar] [CrossRef]
- Jenkins, T.C.; Wallace, R.J.; Moate, P.J.; Mosley, E.E. Board-Invited Review: Recent advances in biohydrogenation of unsaturated fatty acids within the rumen microbial ecosystem1. J. Anim. Sci. 2008, 86, 397–412. [Google Scholar] [CrossRef]
- Francisco, A.E.; Santos-Silva, J.M.; Portugal, A.P.V.; Alves, S.P.; B. Bessa, R.J. Relationship between rumen ciliate protozoa and biohydrogenation fatty acid profile in rumen and meat of lambs. PLoS ONE 2019, 14, e0221996. [Google Scholar] [CrossRef] [Green Version]
- Devillard, E.; McIntosh, F.M.; Newbold, C.J.; Wallace, R.J. Rumen ciliate protozoa contain high concentrations of conjugated linoleic acids and vaccenic acid, yet do not hydrogenate linoleic acid or desaturate stearic acid. Br. J. Nutr. 2006, 96, 697–704. [Google Scholar] [CrossRef]
- Yáñez-Ruiz, D.R.; Williams, S.; Newbold, C.J. The effect of absence of protozoa on rumen biohydrogenation and the fatty acid composition of lamb muscle. Br. J. Nutr. 2007, 97, 938–948. [Google Scholar] [CrossRef] [Green Version]
- Huyen, N.T.; Verstegen, M.W.A.; Hendriks, W.H.; Pellikaan, W.F. Sainfoin (Onobrychis viciifolia) silage in dairy cow rations reduces ruminal biohydrogenation and increases transfer efficiencies of unsaturated fatty acids from feed to milk. Anim. Nutr. 2020, 6, 333–341. [Google Scholar] [CrossRef]
- Vítor, A.C.M.; Francisco, A.E.; Silva, J.; Pinho, M.; Huws, S.A.; Santos-Silva, J.; Bessa, R.J.B.; Alves, S.P. Freeze-dried Nannochloropsis oceanica biomass protects eicosapentaenoic acid (EPA) from metabolization in the rumen of lambs. Sci. Rep. 2021, 11, 21878. [Google Scholar] [CrossRef] [PubMed]
- Jenkins, T.C.; Bridges, W.C., Jr. Protection of fatty acids against ruminal biohydrogenation in cattle. Eur. J. Lipid Sci. Technol. 2007, 109, 778–789. [Google Scholar] [CrossRef]
- Yang, M.; Liang, Z.; Wang, L.; Qi, M.; Luo, Z.; Li, L. Microencapsulation Delivery System in Food Industry—Challenge and the Way Forward. Adv. Polym. Technol. 2020, 2020, 7531810. [Google Scholar] [CrossRef]
- Hafeez, A.; Männer, K.; Schieder, C.; Zentek, J. Effect of supplementation of phytogenic feed additives (powdered vs. encapsulated) on performance and nutrient digestibility in broiler chickens. Poult. Sci. 2016, 95, 622–629. [Google Scholar] [CrossRef]
- Pliego, A.B.; Tavakoli, M.; Khusro, A.; Seidavi, A.; Elghandour, M.M.M.Y.; Salem, A.Z.M.; Márquez-Molina, O.; Rene Rivas-Caceres, R. Beneficial and adverse effects of medicinal plants as feed supplements in poultry nutrition: A review. Anim. Biotechnol. 2022, 33, 369–391. [Google Scholar] [CrossRef]
- Mehta, N.; Kumar, P.; Verma, A.K.; Umaraw, P.; Kumar, Y.; Malav, O.P.; Sazili, A.Q.; Domínguez, R.; Lorenzo, J.M. Microencapsulation as a Noble Technique for the Application of Bioactive Compounds in the Food Industry: A Comprehensive Review. Appl. Sci. 2022, 12, 1424. [Google Scholar] [CrossRef]
- Jain, D.; Atochina-Vasserman, E.N.; Tomer, Y.; Kadire, H.; Beers, M.F. Surfactant Protein D Protects against Acute Hyperoxic Lung Injury. Am. J. Respir. Crit. Care Med. 2008, 178, 805–813. [Google Scholar] [CrossRef] [Green Version]
- Safamehr, A.; Fallah, F.; Nobakht, A. Growth Performance and Biochemical Parameters of Broiler Chickens on Diets Consist of Chichory (Cichorium Intubus) and Nettle (Urtica Dioica) with ot without Multi-Enzyme. Iran. J. Appl. Anim. Sci. 2013, 3, 131–137. [Google Scholar]
- Mousapour, A.; Salarmoini, M.; Afsharmanesh, M.; Ebrahimnejad, H.; Meimandipour, A. Efficacy of Savory Essential Oil Utilization in Conventional and Encapsulated Forms on Performance of Broiler Chickens. Braz. J. Poult. Sci. 2020, 22. [Google Scholar] [CrossRef]
- Muzzarelli, R.A.A. Biochemical significance of exogenous chitins and chitosans in animals and patients. Carbohydr. Polym. 1993, 20, 7–16. [Google Scholar] [CrossRef]
- Kong, M.; Chen, X.G.; Xing, K.; Park, H.J. Antimicrobial properties of chitosan and mode of action: A state of the art review. Int. J. Food Microbiol. 2010, 144, 51–63. [Google Scholar] [CrossRef] [PubMed]
- Matsuhashi, S.; Kume, T. Enhancement of Antimicrobial Activity of Chitosan by Irradiation. Food Agric. 1997, 73, 237–241. [Google Scholar] [CrossRef]
- Goiri, I.; Oregui, L.M.; Garcia-Rodriguez, A. Use of chitosans to modulate ruminal fermentation of a 50:50 forage-to-concentrate diet in sheep1. J. Anim. Sci. 2010, 88, 749–755. [Google Scholar] [CrossRef] [PubMed]
- Wu, Y.; Zheng, Y.; Yang, W.; Wang, C.; Hu, J.; Fu, S. Synthesis and characterization of a novel amphiphilic chitosan–polylactide graft copolymer. Carbohydr. Polym. 2005, 59, 165–171. [Google Scholar] [CrossRef]
- Sosnik, A. Alginate Particles as Platform for Drug Delivery by the Oral Route: State-of-the-Art. ISRN Pharm 2014, 2014, 926157. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hosseini, S.F.; Zandi, M.; Rezaei, M.; Farahmandghavi, F. Two-step method for encapsulation of oregano essential oil in chitosan nanoparticles: Preparation, characterization and in vitro release study. Carbohydr. Polym. 2013, 95, 50–56. [Google Scholar] [CrossRef]
- Keawchaoon, L.; Yoksan, R. Preparation, characterization and in vitro release study of carvacrol-loaded chitosan nanoparticles. Colloids Surf. B Biointerfaces 2011, 84, 163–171. [Google Scholar] [CrossRef]
- Dilbaghi, N.; Kaur, H.; Ahuja, M.; Arora, P.; Kumar, S. Synthesis and evaluation of ciprofloxacin-loaded carboxymethyl tamarind kernel polysaccharide nanoparticles. J. Exp. Nanosci. 2014, 9, 1015–1025. [Google Scholar] [CrossRef]
- Chopra, M.; Kaur, P.; Bernela, M.; Thakur, R. Synthesis and optimization of streptomycin loaded chitosan-alginate nanoparticles. Int. J. Sci. Technol. Res. 2012, 1, 31–34. [Google Scholar]
- Fedorah, P.M.; Hrudey, S.E. A simple apparatus for measuring gas production by methanogenic cultures in serum bottles. Environ. Technol. Lett. 1983, 4, 425–432. [Google Scholar] [CrossRef]
- Maggiolino, A.; Lorenzo, J.M.; Quiñones, J.; Latorre, M.A.; Blando, F.; Centoducati, G.; Dahl, G.E.; De Palo, P. Effects of dietary supplementation with Pinus taeda hydrolyzed lignin on in vivo performances, in vitro nutrient apparent digestibility, and gas emission in beef steers. Anim. Feed. Sci. Technol. 2019, 255, 114217. [Google Scholar] [CrossRef]
- McDougall, E.I. The composition and output of sheep’s saliva. Biochem. J. 1948, 43, 99–109. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- De Bellis, P.; Maggiolino, A.; Albano, C.; De Palo, P.; Blando, F. Ensiling Grape Pomace With and Without Addition of a Lactiplantibacillus plantarum Strain: Effect on Polyphenols and Microbiological Characteristics, in vitro Nutrient Apparent Digestibility, and Gas Emission. Front. Vet. Sci. 2022, 9, 808293. [Google Scholar] [CrossRef] [PubMed]
- AOAC. Official methods of analysis, 18th ed.; Chem, A.O.A., Ed.; Association of Officiating Analytical Chemists: Washington, DC, USA, 2005. [Google Scholar]
- Van Soest, P.J.; Robertson, J.B.; Lewis, B.A. Methods for Dietary Fiber, Neutral Detergent Fiber, and Nonstarch Polysaccharides in Relation to Animal Nutrition. J. Dairy Sci. 1991, 74, 3583–3597. [Google Scholar] [CrossRef]
- Enjalbert, F.; Eynard, P.; Nicot, M.C.; Troegeler-Meynadier, A.; Bayourthe, C.; Moncoulon, R. In Vitro Versus in Situ Ruminal Biohydrogenation of Unsaturated Fatty Acids from a Raw or Extruded Mixture of Ground Canola Seed/Canola Meal. J. Dairy Sci. 2003, 86, 351–359. [Google Scholar] [CrossRef] [Green Version]
- Azadmard-Damirchi, S.; Dutta, P.C. Novel solid-phase extraction method to separate 4-desmethyl-, 4-monomethyl-, and 4,4′-dimethylsterols in vegetable oils. J. Chromatogr. A 2006, 1108, 183–187. [Google Scholar] [CrossRef]
- Soottitantawat, A.; Bigeard, F.; Yoshii, H.; Furuta, T.; Ohkawara, M.; Linko, P. Influence of emulsion and powder size on the stability of encapsulated d-limonene by spray drying. Innov. Food Sci. Emerg. Technol. 2005, 6, 107–114. [Google Scholar] [CrossRef]
- Klinkesorn, U.; Sophanodora, P.; Chinachoti, P.; Decker, E.A.; McClements, D.J. Characterization of spray-dried tuna oil emulsified in two-layered interfacial membranes prepared using electrostatic layer-by-layer deposition. Food Res. Int. 2006, 39, 449–457. [Google Scholar] [CrossRef]
- Drusch, S.; Berg, S. Extractable oil in microcapsules prepared by spray-drying: Localisation, determination and impact on oxidative stability. Food Chem. 2008, 109, 17–24. [Google Scholar] [CrossRef]
- Drusch, S.; Serfert, Y.; Berger, A.; Shaikh, M.Q.; Rätzke, K.; Zaporojtchenko, V.; Schwarz, K. New insights into the microencapsulation properties of sodium caseinate and hydrolyzed casein. Food Hydrocoll. 2012, 27, 332–338. [Google Scholar] [CrossRef]
- Hogan, S.A.; O’Riordan, E.D.; O’Sullivan, M. Microencapsulation and oxidative stability of spray-dried fish oil emulsions. J. Microencapsul. 2003, 20, 675–688. [Google Scholar] [CrossRef]
- Heinzelmann, K.; Franke, K. Using freezing and drying techniques of emulsions for the microencapsulation of fish oil to improve oxidation stability. Colloids Surf. B Biointerfaces 1999, 12, 223–229. [Google Scholar] [CrossRef]
- Lin, C.-C.; Lin, S.-Y.; Hwang, L.S. Microencapsulation of Squid Oil with Hydrophilic Macromolecules for Oxidative and Thermal Stabilization. J. Food Sci. 1995, 60, 36–39. [Google Scholar] [CrossRef]
- Velasco, J.; Dobarganes, M.C.; Márquez-Ruiz, G. Oxidation of free and encapsulated oil fractions in dried microencapsulated fish oils. Grasas Aceites 2000, 51, 439–446. [Google Scholar] [CrossRef]
- Swetank, Y.; Karthik, P.; Anandharamakrishnan, C. Effect of whey protein isolate and b-cyclodextrin wall systems on stability of microencapsulated vanillin by spray–freeze drying method. J. Food Chemisry Technol. 2015, 174, 16–24. [Google Scholar]
- Liu, S.; Low, N.H.; Nickerson, M.T. Entrapment of Flaxseed Oil Within Gelatin-Gum Arabic Capsules. J. Am. Oil Chem. Soc. 2010, 87, 809–815. [Google Scholar] [CrossRef]
- Ajun, W.; Yan, S.; Li, G.; Huili, L. Preparation of aspirin and probucol in combination loaded chitosan nanoparticles and in vitro release study. Carbohydr. Polym. 2009, 75, 566–574. [Google Scholar] [CrossRef]
- Roy, B.; Guha, P.; Nahak, P.; Karmakar, G.; Maiti, S.; Mandal, A.K.; Bykov, A.G.; Akentiev, A.V.; Noskov, B.A.; Tsuchiya, K.; et al. Biophysical Correlates on the Composition, Functionality, and Structure of Dendrimer–Liposome Aggregates. ACS Omega 2018, 3, 12235–12245. [Google Scholar] [CrossRef]
- García-Moreno, P.J.; Guadix, A.; Guadix, E.M.; Jacobsen, C. Physical and oxidative stability of fish oil-in-water emulsions stabilized with fish protein hydrolysates. Food Chem. 2016, 203, 124–135. [Google Scholar] [CrossRef] [Green Version]
- Greenwood, R.; Kendall, K. Selection of Suitable Dispersants for Aqueous Suspensions of Zirconia and Titania Powders using Acoustophoresis. J. Eur. Ceram. Soc. 1999, 19, 479–488. [Google Scholar] [CrossRef]
- Calvo, P.; Remuñán-López, C.; Vila-Jato, J.L.; Alonso, M.J. Novel hydrophilic chitosan-polyethylene oxide nanoparticles as protein carriers. J. Appl. Polym. Sci. 1997, 63, 125–132. [Google Scholar] [CrossRef]
- Bahramizadeh, I.; Rahmanifarah, K. Effect of different concentrations of xanthan and alginate on the quality of encapsulated fish oil. Iran. Sci. Fish. J. 2017, 26, 139–150. [Google Scholar]
- Mansuri, H.; Nikkhah, A.; Rezaeian, M.; Moradi Shahrbaback, M.; Mirhadi, M. Determination of Roughages degradability through In vitro gas production and nylon bag techniques. Iranain J. Agric. Sci. 2003, 34, 495–507. [Google Scholar]
- Sinclair, L.A.; Cooper, S.L.; Huntington, J.A.; Wilkinson, R.G.; Hallett, K.G.; Enser, M.; Wood, J.D. In vitro biohydrogenation of n−3 polyunsaturated fatty acids protected against ruminal microbial metabolism. Anim. Feed. Sci. Technol. 2005, 123–124, 579–596. [Google Scholar] [CrossRef]
- Safari, R.; Valizadeh, R.; Kadkhodayi, R.; Alamolhada, H.; Tahmasbi, A.; Naserian, A. Investigation of the resistance of fish oil microcapsules in ruminal conditions and their effect on gas production and digestibility in invitro. Iran. J. Anim. Sci. Res. 2011, 4, 265–273. [Google Scholar]
- Da Silva, M.S.; Julien, P.; Pérusse, L.; Vohl, M.-C.; Rudkowska, I. Natural Rumen-Derived trans Fatty Acids Are Associated with Metabolic Markers of Cardiac Health. Lipids 2015, 50, 873–882. [Google Scholar] [CrossRef] [PubMed]
- Mapato, C.; Wanapat, M.; Cherdthong, A. Effects of urea treatment of straw and dietary level of vegetable oil on lactating dairy cows. Trop. Anim. Health Prod. 2010, 42, 1635–1642. [Google Scholar] [CrossRef]
- Wanapat, M.; Kongmun, P.; Poungchompu, O.; Cherdthong, A.; Khejornsart, P.; Pilajun, R.; Kaenpakdee, S. Effects of plants containing secondary compounds and plant oils on rumen fermentation and ecology. Trop. Anim. Health Prod. 2012, 44, 399–405. [Google Scholar] [CrossRef]
- Jenkins, T.C. Lipid Metabolism in the Rumen. J. Dairy Sci. 1993, 76, 3851–3863. [Google Scholar] [CrossRef]
- Zinn, R.A.; Gulati, S.K.; Plascencia, A.; Salinas, J. Influence of ruminal biohydrogenation on the feeding value of fat in finishing diets for feedlot cattle. J. Anim. Sci. 2000, 78, 1738–1746. [Google Scholar] [CrossRef] [Green Version]
- Bhatt, R.S.; Karim, S.A.; Sahoo, A.; Shinde, A.K. Growth performance of lambs fed diet supplemented with rice bran oil as such or as calcium soap. Asian-Australas J. Anim. Sci. 2013, 26, 812–819. [Google Scholar] [CrossRef] [PubMed]
- Jordan, E.; Kenny, D.; Hawkins, M.; Malone, R.; Lovett, D.K.; O’Mara, F.P. Effect of refined soy oil or whole soybeans on intake, methane output, and performance of young bulls1. J. Anim. Sci. 2006, 84, 2418–2425. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Machmüller, A.; Ossowski, D.A.; Kreuzer, M. Comparative evaluation of the effects of coconut oil, oilseeds and crystalline fat on methane release, digestion and energy balance in lambs. Anim. Feed. Sci. Technol. 2000, 85, 41–60. [Google Scholar] [CrossRef]
- Machmüller, A.; Soliva, C.R.; Kreuzer, M. Effect of coconut oil and defaunation treatment on methanogenesis in sheep. Reprod. Nutr. Dev. 2003, 43, 41–55. [Google Scholar] [CrossRef] [Green Version]
- Beauchemin, K.A.; Kreuzer, M.; O’Mara, F.; McAllister, T.A. Nutritional management for enteric methane abatement: A review. Aust. J. Exp. Agric. 2008, 48, 21–27. [Google Scholar] [CrossRef]
- Naik, P.K.; Saijpaul, S.; Rani, N. Effect of ruminally protected fat on in vitro fermentation and apparent nutrient digestibility in buffaloes (Bubalus bubalis). Anim. Feed. Sci. Technol. 2009, 153, 68–76. [Google Scholar] [CrossRef]
- Bhatt, R.S.; Sahoo, A. Effect of feeding complete feed block containing rumen protected protein, non-protein nitrogen and rumen protected fat on improving body condition and carcass traits of cull ewes. J. Anim. Physiol. Anim. Nutr. 2017, 101, 1147–1158. [Google Scholar] [CrossRef]
- Belanche, A.; Pinloche, E.; Preskett, D.; Newbold, C.J. Effects and mode of action of chitosan and ivy fruit saponins on the microbiome, fermentation and methanogenesis in the rumen simulation technique. FEMS Microbiol. Ecol. 2015, 92. [Google Scholar] [CrossRef] [Green Version]
- Bach, A.; Calsamiglia, S.; Stern, M.D. Nitrogen Metabolism in the Rumen. J. Dairy Sci. 2005, 88, E9–E21. [Google Scholar] [CrossRef] [Green Version]
- Chouinard, P.Y.; Corneau, L.; Bauman, D.E.; Butler, W.R.; Chilliard, Y.; Drackley, K. Conjugated linoleic acid content of milk from cows fed different sources of dietary fat. In Proceedings of the American Society of Animal Science and American Dairy Science Association meeting, Denver, CO, USA, 28–31 July 1998. [Google Scholar]
- Jenkins, T.C.; Fotouhi, N. Effects of lecithin and corn oil on site of digestion, ruminal fermentation and microbial protein synthesis in sheep. J. Anim. Sci. 1990, 68, 460–466. [Google Scholar] [CrossRef]
- Maia, M.R.G.; Chaudhary, L.C.; Bestwick, C.S.; Richardson, A.J.; McKain, N.; Larson, T.R.; Graham, I.A.; Wallace, R.J. Toxicity of unsaturated fatty acids to the biohydrogenating ruminal bacterium, Butyrivibrio fibrisolvens. BMC Microbiol. 2010, 10, 52. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schauff, D.J.; Clark, J.H. Effects of Feeding Diets Containing Calcium Salts of Long-Chain Fatty Acids to Lactating Dairy Cows1. J. Dairy Sci. 1992, 75, 2990–3002. [Google Scholar] [CrossRef]
- Geraeily, M. The Effect of Flaxseed Oil on Performance of Dairy Calves under Heat Stress Conditions. Master’s Thesis, Gorgan University of Agricultural Sciences and Natural Resources, Gorgan, Iran, 2017. (In Persian). [Google Scholar]
- Schroeder, J.W.; Bauer, M.L.; Bork, N.R. Effect of flaxseed physical form on digestibility of lactation diets fed to Holstein steers. J. Dairy Sci. 2014, 97, 5718–5728. [Google Scholar] [CrossRef] [PubMed]
- Harfoot, C.G.; HAzlewood, G.P. Lipid metabolism in the rumen. In The Rumen Microbial Ecosytem; Hobson, P., Stewart, C.S., Eds.; Springer: Berlin/Heidelberg, Germany, 1997; pp. 382–419. [Google Scholar]
- Hosoda, K.; Kuramoto, K.; Eruden, B.; Nishida, T.; Shioya, S. The effects of three herbs as feed supplements on blood metabolites, hormones, antioxidant activity, IgG concentration, and ruminal fermentation in Holstein steers. Asian-Australas J. Anim. Sci. 2006, 19, 35–41. [Google Scholar] [CrossRef]
- Yuan, G.; Chen, X.; Li, D. Chitosan films and coatings containing essential oils: The antioxidant and antimicrobial activity, and application in food systems. Food Res. Int. 2016, 89, 117–128. [Google Scholar] [CrossRef] [PubMed]
- Glasser, F.; Ferlay, A.; Chilliard, Y. Oilseed Lipid Supplements and Fatty Acid Composition of Cow Milk: A Meta-Analysis. J. Dairy Sci. 2008, 91, 4687–4703. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wood, J.D.; Enser, M.; Fisher, A.V.; Nute, G.R.; Sheard, P.R.; Richardson, R.I.; Hughes, S.I.; Whittington, F.M. Fat deposition, fatty acid composition and meat quality: A review. Meat Sci. 2008, 78, 343–358. [Google Scholar] [CrossRef]
- Dohme, F.; Fievez, V.; Raes, K.; Demeyer, D.I. Increasing levels of two different fish oils lower ruminal biohydrogenation of eicosapentaenoic and docosahexaenoic acid in vitro. Anim. Res. 2003, 52, 309–320. [Google Scholar] [CrossRef]
- Chilliard, Y.; Glasser, F.; Ferlay, A.; Bernard, L.; Rouel, J.; Doreau, M. Diet, rumen biohydrogenation and nutritional quality of cow and goat milk fat. Eur. J. Lipid Sci. Technol. 2007, 109, 828–855. [Google Scholar] [CrossRef]
- Liu, Z.; Yang, D.; Chen, P.; Lin, S.; Jiang, X.; Zhao, W.; Dong, W. Effect of dietary sources of roasted oilseeds on blood parameters and milk fatty acid composition. J. Anim. Sci. 2008, 5, 219–226. [Google Scholar] [CrossRef] [Green Version]
- Khalili, S.T.; Mohsenifar, A.; Beyki, M.; Zhaveh, S.; Rahmani-Cherati, T.; Abdollahi, A.; Bayat, M.; Tabatabaei, M. Encapsulation of Thyme essential oils in chitosan-benzoic acid nanogel with enhanced antimicrobial activity against Aspergillus flavus. LWT Food Sci. Technol. 2015, 60, 502–508. [Google Scholar] [CrossRef]
- Szumacher-Strabel, M.; Cieślak, A.; Nowakowska, A. Effect of oils rich in linoleic acid on in vitro rumen fermentation parameters of sheep, goats and dairy cows. J. Anim. Feed Sci. 2009, 18, 440–452. [Google Scholar] [CrossRef]
- Elnashar, M.; Gawad, R.; Strabel, M.; Abo El-Nor, S.; Kattab, H.; Cieslak, A. Encapsulation method to protect unsaturated fatty acids from rumen biohydrogenation in vitro. J. Innov. Pharm. Biol. Sci. 2015, 2, 240–251. [Google Scholar]
- Kim, T.-B.; Lee, J.-S.; Cho, S.-Y.; Lee, H.-G. In Vitro and In Vivo Studies of Rumen-Protected Microencapsulated Supplement Comprising Linseed Oil, Vitamin E, Rosemary Extract, and Hydrogenated Palm Oil on Rumen Fermentation, Physiological Profile, Milk Yield, and Milk Composition in Dairy Cows. Animals 2020, 10, 1631. [Google Scholar] [CrossRef] [PubMed]
- Khalilvandi-Behroozyar, H.; Dehghan-Banadaky, M.; Pirmohammadi, R.; AsadNejad, B. Evaluation of Nutritional efficiency of encapsulated fish oil ca-salts in vitro and in vivo. Iran. J. Anim. Sci. 2018, 48, 505–521. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
| Diet Ingredients | Content (%) |
|---|---|
| Alfalfa hay | 21 |
| Corn silage | 19 |
| Beet pulp | 7 |
| Wheat bran | 2 |
| Concentrate | 51 |
| Chemical Composition | |
| Dry matter | 59 |
| Organic matter | 92.86 |
| Crude protein | 20.3 |
| Neutral detergent fiber | 38.57 |
| Acid detergent fiber | 20.00 |
| Efficiency (%) | Zeta Potential (mv) | Particle Size (nm) | |
|---|---|---|---|
| chitosan 500 ppm | 87.47 | +56.2 | 190.2 |
| chitosan 1000 ppm | 67.45 | +45.5 | - |
| calcium alginate 500 ppm | 74.50 | +0.9 | 334 |
| calcium alginate 1000 ppm | 53.28 | +0.6 | - |
| Treatments | Incubation Times (h) | ||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| 2 | 4 | 6 | 8 | 12 | 16 | 24 | 36 | 48 | 72 | 96 | |
| Control | 5.52 a | 15.04 b,c | 26.79 b,c | 38.29 a,b,c | 56.77 b | 74.45 c,d | 93.69 b,c | 109.42 d | 120.31 c | 126.30 c | 130.41 c |
| flaxseed oil (7%) | 5.39 a | 17.70 a,b | 30.19 a,b | 42.08 a,b | 69.56 a | 98.83 a | 116.18 a | 138.90 a | 147.93 a | 153.66 a | 156.44 a |
| flaxseed oil (14%) | 2.79 b | 11.89 c | 20.33 c | 31.29 c | 46.12 c | 63.80 d | 75.84 d | 88.31 e | 94.81 d | 98.34 d | 99.72 d |
| Chitosan (7%) | 6.19 a | 15.11 b,c | 26.13 b,c | 37.22 b,c | 54.24 b,c | 70.92 c,d | 88.61 c,d | 115.83 c,d | 127.55 b,c | 132.62 bc | 135.79 b,c |
| Chitosan (14%) | 3.66 a,b | 17.64 a,b | 28.59 a,b | 40.09 a,b,c | 64.72 a,b | 87.82 a,b | 103.93 a,b | 129.98 a,b,c | 138.28 a,b | 146.93 a,b | 150.78 a,b |
| Calcium alginate (7%) | 5.52 a | 16.79 b,c | 28.94 a,b | 42.90 a,b | 61.19 a,b | 81.53 b,c | 98.17 b,c | 122.22 b,c,d | 131.85 a,b,c | 139.84 a,b,c | 144.02 a,b,c |
| Calcium alginate (14%) | 5.46 a | 22.38 a | 35.27 a | 47.29 a | 70.38 a | 92.79 a,b | 107.22 a,b | 133.08 a,b | 140.24 a,b | 148.50 a,b | 153.21 a,b |
| SEM | 0.788 | 1.711 | 2.391 | 2.927 | 3.371 | 4.204 | 4.426 | 5.098 | 5.420 | 5.924 | 6.040 |
| p-value | 0.053 | 0.0096 | 0.0094 | 0.022 | 0.0002 | <0.0001 | <0.0001 | <0.0001 | <0.0001 | <0.0001 | <0.0001 |
| Treatments | Incubation Times (h) | ||||
|---|---|---|---|---|---|
| 2 | 4 | 8 | 12 | 24 | |
| Dry matter digestibility | |||||
| Control | 32.42 a | 34.85 b | 49.81 a | 51.18 b | 61.10 b |
| flaxseed oil (7%) | 25.99 b | 32.44 b | 37.54 d | 45.76 c | 49.23 d |
| flaxseed oil (14%) | 32.40 a | 33.87 b | 39.54 cd | 44.21 c,d | 53.73 c |
| Chitosan (7%) | 21.98 c | 23.74 c | 36.60 d | 42.56 d | 46.35 d,e |
| Chitosan (14%) | 25.61 b | 33.73 b | 37.25 d | 42.07 d | 43.58 e |
| Calcium alginate (7%) | 23.11 b,c | 24.66 c | 42.60 b,c | 45.76 c | 74.40 a |
| Calcium alginate (14%) | 33.55 a | 38.93 a | 45.75 b | 54.32 a | 75.06 a |
| SEM | 1.007 | 0.966 | 1.162 | 0.729 | 1.233 |
| p-value | <0.0001 | <0.0001 | <0.0001 | <0.0001 | <0.0001 |
| Organic matter digestibility | |||||
| Control | 39.71 a | 44.36 b | 50.50 b,c | 55.49 a,b,c | 61.84 a,b,c |
| flaxseed oil (7%) | 31.19 c | 42.50 b | 47.46 c,d | 54.28 b,c | 58.03 c,d |
| flaxseed oil (14%) | 28.01 d | 43.07 b | 45.88 d | 58.10 a,b | 60.33 b,c |
| Chitosan (7%) | 20.95 e | 29.22 c | 47.65 b,c,d | 57.83 a,b | 61.80 a,b,c |
| Chitosan (14%) | 36.14 b | 49.87 a | 51.13 b | 52.42 c | 54.71 d |
| Calcium alginate (7%) | 22.22 e | 26.20 c | 47.01 c,d | 58.57 a,b | 63.57 a,b |
| Calcium alginate (14%) | 31.59 c | 51.18 a | 55.61 a | 60.30 a | 66.62 a |
| SEM | 0.743 | 1.678 | 1.107 | 1.622 | 1.610 |
| p-value | <0.0001 | <0.0001 | 0.0003 | 0.048 | 0.0035 |
| Crude protein digestibility | |||||
| Control | 27.72 c | 48.88 c | 53.99 b | 59.30 b | 67.71 a |
| flaxseed oil (7%) | 40.59 b | 45.39 c,d | 54.47 b | 67.55 a | 68.58 a |
| flaxseed oil (14%) | 37.20 b | 53.70 b | 63.40 a | 66.81 a | 69.85 a |
| Chitosan (7%) | 19.37 d | 43.01 d | 49.29 c | 50.73 d | 63.60 b |
| Chitosan (14%) | 38.86 b | 52.71 b | 55.42 b | 61.06 b | 69.70 a |
| Calcium alginate (7%) | 31.45 c | 36.77 e | 49.29 c | 55.08 c | 57.36 c |
| Calcium alginate (14%) | 50.97 a | 63.35 a | 66.38 a | 67.79 a | 69.59 a |
| SEM | 1.623 | 1.148 | 1.048 | 0.819 | 0.681 |
| p-value | <0.0001 | <0.0001 | <0.0001 | <0.0001 | <0.0001 |
| NDF digestibility | |||||
| Control | 37.30 b | 46.78 a,b | 47.42 c | 54.88 b | 64.57 a |
| flaxseed oil (7%) | 28.25 c | 43.51 a,b,c | 50.35 a,b | 54.74 b | 57.67 b |
| flaxseed oil (14%) | 27.09 c | 46.54 a,b | 47.14 b,c | 53.10 b | 55.52 b,c |
| Chitosan (7%) | 38.16 b | 38.51 c | 52.39 a | 53.64 b | 54.71 c |
| Chitosan (14%) | 45.95 a | 46.95 a,b | 52.12 a,b | 54.20 b | 55.08 c |
| Calcium alginate (7%) | 38.98 b | 42.32 b,c | 49.51 a,b | 57.11 a,b | 66.85 a |
| Calcium alginate (14%) | 35.76 b | 47.85 a | 51.85 a,b | 62.81 a | 65.97 a |
| SEM | 1.452 | 1.616 | 2.430 | 1.885 | 0.799 |
| p-value | <0.0001 | 0.013 | 0.035 | 0.035 | <0.0001 |
| ADF digestibility | |||||
| Control | 23.96 c | 31.65 b | 42.83 a,b,c | 52.83 a,b | 57.30 b |
| flaxseed oil (7%) | 28.20 b | 33.05 b | 39.94 b,c | 52.64 a,b | 57.11 b |
| flaxseed oil (14%) | 10.49 d | 34.87 b | 45.06 a,b | 56.27 a | 58.05 b |
| Chitosan (7%) | 23.34 c | 25.33 c | 36.94 c | 45.85 c | 51.24 c |
| Chitosan (14%) | 31.22 b | 33.94 b | 36.69 c | 46.08 c | 51.39 c |
| Calcium alginate (7%) | 30.41 b | 33.46 b | 48.31 a | 52.62 a,b | 55.60 b |
| Calcium alginate (14%) | 37.36 a | 39.58 a | 44.60 a,b,c | 49.43 b,c | 63.20 a |
| SEM | 1.032 | 1.215 | 2.504 | 1.824 | 1.193 |
| p-value | <0.0001 | <0.0001 | 0.035 | 0.0098 | <0.0001 |
| Treatments | ||||||||||
|---|---|---|---|---|---|---|---|---|---|---|
| Items | Time (h) | Control | Flaxseed Oil (7%) | Flaxseed Oil (14%) | Chitosan (7%) | Chitosan (14%) | Calcium Alginate (7%) | Calcium Alginate (14%) | SEM | p-Value |
| SFA | 2 | 79.52 c | 80.31 c | 61.14 d | 56.25 e | 52.37 f | 98.60 a | 91.40 b | 0.406 | <0.0001 |
| 4 | 82.62 d | 82.86 d | 96.17 c | 59.31 e | 53.82 f | 99.11 a | 92.15 b | 0.692 | <0.0001 | |
| 24 | 84.16 d | 92.98 a | 88.46 b | 61.05 d | 68.69 c | 96.00 a | 92.79 a | 0.958 | <0.0001 | |
| USFA | 2 | 20.45 c | 18.46 c | 38.63 b | 39.56 b | 42.42 a | 1.22 e | 4.92 d | 0.380 | <0.0001 |
| 4 | 17.03 c | 16.42 c | 12.19 d | 35.29 b | 40.22 a | 0.48 f | 3.91 e | 0.234 | <0.0001 | |
| 24 | 15.08 b | 4.61 c | 10.74 b,c | 37.88 a | 13.93 b | 1.98 c | 3.12 c | 2.534 | <0.0001 | |
| SCFA | 2 | 31.85 d | 36.07 c | 15.14 d | 3.26 f | 5.39 e | 77.11 a | 40.54 b | 0.224 | <0.0001 |
| 4 | 29.75 e | 45.45 b | 39.00 c | 8.87 e | 33.95 d | 64.55 a | 34.56 d | 0.954 | <0.0001 | |
| 24 | 39.66 d | 47.65 c | 30.65 e | 13.30 f | 53.82 b | 70.77 a | 36.45 d | 0.741 | <0.0001 | |
| MCFA | 2 | 7.42 f | 17.55 c | 16.91 c | 28.56 b | 45.61 a | 14.06 d | 12.32 e | 0.308 | <0.0001 |
| 4 | 6.60 e | 9.70 d | 25.80 a | 25.53 a | 22.87 b | 25.83 a | 16.49 c | 0.491 | <0.0001 | |
| 24 | 5.92 f | 12.83 e | 19.65 b | 26.99 a | 17.22 c | 12.53 e | 14.54 d | 0.368 | <0.0001 | |
| LCFA | 2 | 35.45 d | 16.86 d | 48.02 b | 53.92 a | 46.56 b | 8.67 e | 41.47 c | 0.791 | <0.0001 |
| 4 | 38.24 c | 22.64 d | 23.48 d | 50.34 a | 34.91 c | 7.82 e | 45.32 b | 0.448 | <0.0001 | |
| 24 | 30.79 c | 17.83 e | 30.93 c | 47.85 a | 28.88 d | 15.45 f | 40.36 b | 0.336 | <0.0001 | |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Besharati, M.; Giannenas, I.; Palangi, V.; Ayasan, T.; Noorian, F.; Maggiolino, A.; Lorenzo, J.M. Chitosan/Calcium–Alginate Encapsulated Flaxseed Oil on Dairy Cattle Diet: In Vitro Fermentation and Fatty Acid Biohydrogenation. Animals 2022, 12, 1400. https://doi.org/10.3390/ani12111400
Besharati M, Giannenas I, Palangi V, Ayasan T, Noorian F, Maggiolino A, Lorenzo JM. Chitosan/Calcium–Alginate Encapsulated Flaxseed Oil on Dairy Cattle Diet: In Vitro Fermentation and Fatty Acid Biohydrogenation. Animals. 2022; 12(11):1400. https://doi.org/10.3390/ani12111400
Chicago/Turabian StyleBesharati, Maghsoud, Ilias Giannenas, Valiollah Palangi, Tugay Ayasan, Fatemeh Noorian, Aristide Maggiolino, and Jose Manuel Lorenzo. 2022. "Chitosan/Calcium–Alginate Encapsulated Flaxseed Oil on Dairy Cattle Diet: In Vitro Fermentation and Fatty Acid Biohydrogenation" Animals 12, no. 11: 1400. https://doi.org/10.3390/ani12111400
APA StyleBesharati, M., Giannenas, I., Palangi, V., Ayasan, T., Noorian, F., Maggiolino, A., & Lorenzo, J. M. (2022). Chitosan/Calcium–Alginate Encapsulated Flaxseed Oil on Dairy Cattle Diet: In Vitro Fermentation and Fatty Acid Biohydrogenation. Animals, 12(11), 1400. https://doi.org/10.3390/ani12111400