
The Invasive Bank Vole (Myodes glareolus): A Model System for Studying Parasites and Ecoimmunology during a Biological Invasion


Abstract
:Simple Summary
Abstract
1. Emerging Infectious Diseases and the Need for Wildlife Models
2. Cycling Rodent Populations as Wildlife Disease Models
3. Factors Present in the Irish Invasion System Making It an Ideal Study System for Parasite Dynamics
4. Opportunities Presented by the Bank Vole Invasion Model
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Mackenzie, J.S.; Smith, D.W. COVID-19: A novel zoonotic disease caused by a coronavirus from China: What we know and what we don’t. Microbiol. Aust. 2020, 41, 45–50. [Google Scholar] [CrossRef]
- World Health Organization. WHO Coronavirus (COVID-19) Dashboard. Available online: https://covid19.who.int/ (accessed on 24 June 2021).
- Daszak, P.; Olival, K.J.; Li, H. A strategy to prevent future epidemics similar to the 2019-nCoV outbreak. Biosaf. Health 2020, 2, 6–8. [Google Scholar] [CrossRef] [PubMed]
- Le, T.T.; Andreadakis, Z.; Kumar, A.; Román, R.G.; Tollefsen, S.; Saville, M.; Mayhew, S. The COVID-19 vaccine development landscape. Nat. Rev. Drug Discov. 2020, 19, 305–306. [Google Scholar] [CrossRef] [PubMed]
- Zhu, N.; Zhang, D.; Wang, W.; Li, X.; Yang, B.; Song, J.; Zhao, X.; Huang, B.; Shi, W.; Lu, R. A novel coronavirus from patients with pneumonia in China, 2019. N. Engl. J. Med. 2020, 382(8), 727–733. [Google Scholar] [CrossRef]
- World Health Organization. WHO-Convened Global Study of Origins of SARS-CoV-2: China Part; WHO: Geneva, Switzerland, 2021. [Google Scholar]
- Jones, K.E.; Patel, N.G.; Levy, M.A.; Storeygard, A.; Balk, D.; Gittleman, J.L.; Daszak, P. Global trends in emerging infectious diseases. Nature 2008, 451, 990–993. [Google Scholar] [CrossRef]
- Wikelski, M.; Foufopoulos, J.; Vargas, H.; Snell, H. Galápagos birds and diseases: Invasive pathogens as threats for island species. Ecol. Soc. 2004, 9, 5. [Google Scholar] [CrossRef]
- Becker, D.J.; Albery, G.F.; Kessler, M.K.; Lunn, T.J.; Falvo, C.A.; Czirják, G.Á.; Martin, L.B.; Plowright, R.K. Macroimmunology: The drivers and consequences of spatial patterns in wildlife immune defence. J. Anim. Ecol. 2020, 89, 972–995. [Google Scholar] [CrossRef]
- Schreiner, C.L.; Nuismer, S.L.; Basinski, A.J. When to vaccinate a fluctuating wildlife population: Is timing everything? J. Appl. Ecol. 2020, 57, 307–319. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Daszak, P.; Cunningham, A.A.; Hyatt, A.D. Emerging infectious diseases of wildlife--threats to biodiversity and human health. Science 2000, 287, 443–449. [Google Scholar] [CrossRef]
- Wilcox, B.A.; Gubler, D.J. Disease ecology and the global emergence of zoonotic pathogens. Environ. Health Prev. Med. 2005, 10, 263–272. [Google Scholar] [CrossRef]
- Hatcher, M.J.; Dick, J.T.; Dunn, A.M. Disease emergence and invasions. Funct. Ecol. 2012, 26, 1275–1287. [Google Scholar] [CrossRef]
- Chalkowski, K.; Lepczyk, C.A.; Zohdy, S. Parasite ecology of invasive species: Conceptual framework and new hypotheses. Trends Parasitol. 2018, 34, 655–663. [Google Scholar] [CrossRef]
- Zhu, G.-L.; Tang, Y.-Y.; Limpanont, Y.; Wu, Z.-D.; Li, J.; Lv, Z.-Y. Zoonotic parasites carried by invasive alien species in China. Infect. Dis. Poverty 2019, 8, 1–17. [Google Scholar] [CrossRef] [PubMed]
- Sainsbury, A.W.; Nettleton, P.; Gilray, J.; Gurnell, J. Grey squirrels have high seroprevalence to a parapoxvirus associated with deaths in red squirrels. Anim. Conserv. 2000, 3, 229–233. [Google Scholar] [CrossRef]
- Tompkins, D.M.; Sainsbury, A.W.; Nettleton, P.; Buxton, D.; Gurnell, J. Parapoxvirus causes a deleterious disease in red squirrels associated with UK population declines. Proc. R. Soc. London Ser. B Biol. Sci. 2002, 269, 529–533. [Google Scholar] [CrossRef] [Green Version]
- Strauss, A.; White, A.; Boots, M. Invading with biological weapons: The importance of disease-mediated invasions. Funct. Ecol. 2012, 26, 1249–1261. [Google Scholar] [CrossRef]
- Dunn, A.M.; Hatcher, M.J. Parasites and biological invasions: Parallels, interactions, and control. Trends Parasitol. 2015, 31, 189–199. [Google Scholar] [CrossRef] [PubMed]
- Chinchio, E.; Crotta, M.; Romeo, C.; Drewe, J.A.; Guitian, J.; Ferrari, N. Invasive alien species and disease risk: An open challenge in public and animal health. PLoS Pathog. 2020, 16, e1008922. [Google Scholar] [CrossRef] [PubMed]
- Kelly, D.; Paterson, R.; Townsend, C.; Poulin, R.; Tompkins, D. Parasite spillback: A neglected concept in invasion ecology? Ecology 2009, 90, 2047–2056. [Google Scholar] [CrossRef]
- Stuart, P.; Paredis, L.; Henttonen, H.; Lawton, C.; Torres, C.A.O.; Holland, C.V. The hidden faces of a biological invasion: Parasite dynamics of invaders and natives. Int. J. Parasitol. 2020, 50, 111–123. [Google Scholar] [CrossRef]
- Cornet, S.; Brouat, C.; Diagne, C.; Charbonnel, N. Eco-immunology and bioinvasion: Revisiting the evolution of increased competitive ability hypotheses. Evol. Appl. 2016, 9, 952–962. [Google Scholar] [CrossRef] [Green Version]
- Lee, K.A.; Klasing, K.C. A role for immunology in invasion biology. Trends Ecol. Evol. 2004, 19, 523–529. [Google Scholar] [CrossRef]
- Diagne, C.; Galan, M.; Tamisier, L.; d’Ambrosio, J.; Dalecky, A.; Bâ, K.; Kane, M.; Niang, Y.; Diallo, M.; Sow, A. Ecological and sanitary impacts of bacterial communities associated to biological invasions in African commensal rodent communities. Sci. Rep. 2017, 7, 1–11. [Google Scholar] [CrossRef] [Green Version]
- Loxton, K. Parasites in a Host Species’ Invasion: A Unique Small Mammal Model System. Ph.D. Thesis, Trinity College Dublin, Dublin, Ireland, 2015; p. 21. [Google Scholar]
- Diagne, C.; Granjon, L.; Gueye, M.S.; Kane, M.; Niang, Y.; Tatard, C.; Brouat, C. Association between temporal patterns in helminth assemblages and successful range expansion of exotic Mus musculus domesticus in Senegal. Biol. Invasions 2020, 22, 3003–3016. [Google Scholar] [CrossRef]
- Hatcher, M.J.; Dunn, A.M. Parasites in Ecological Communities: From Interactions to Ecosystems; Cambridge University Press: New York, NY, USA, 2011. [Google Scholar]
- Stephens, C.R.; Heau, J.G.; González, C.; Ibarra-Cerdeña, C.N.; Sánchez-Cordero, V.; González-Salazar, C. Using biotic interaction networks for prediction in biodiversity and emerging diseases. PLoS ONE 2009, 4, e5725. [Google Scholar] [CrossRef]
- Roche, B.; Garchitorena, A.; Guégan, J.F.; Arnal, A.; Roiz, D.; Morand, S.; Zambrana-Torrelio, C.; Suzán, G.; Daszak, P. Was the COVID-19 pandemic avoidable? A call for a “solution-oriented” approach in pathogen evolutionary ecology to prevent future outbreaks. Ecol. Lett. 2020, 23, 1557–1560. [Google Scholar] [CrossRef]
- Morand, S. Emerging diseases, livestock expansion and biodiversity loss are positively related at global scale. Biol. Conserv. 2020, 248, 108707. [Google Scholar] [CrossRef]
- Norman, R.; Bowers, R.; Begon, M.; Hudson, P.J. Persistence of tick-borne virus in the presence of multiple host species: Tick reservoirs and parasite mediated competition. J. Theor. Biol. 1999, 200, 111–118. [Google Scholar] [CrossRef] [Green Version]
- Schmidt, K.A.; Ostfeld, R.S. Biodiversity and the dilution effect in disease ecology. Ecology 2001, 82, 609–619. [Google Scholar] [CrossRef]
- Tierney, P.A.; Caffrey, J.M.; Vogel, S.; Matthews, S.M.; Costantini, E.; Holland, C.V. Invasive freshwater fish (Leuciscus leuciscus) acts as a sink for a parasite of native brown trout Salmo trutta. Biol. Invasions 2020, 22, 2235–2250. [Google Scholar] [CrossRef]
- Han, B.A.; Schmidt, J.P.; Bowden, S.E.; Drake, J.M. Rodent reservoirs of future zoonotic diseases. Proc. Natl. Acad. Sci. USA 2015, 112, 7039–7044. [Google Scholar] [CrossRef] [Green Version]
- Grzybek, M.; Tołkacz, K.; Sironen, T.; Mäki, S.; Alsarraf, M.; Behnke-Borowczyk, J.; Biernat, B.; Nowicka, J.; Vaheri, A.; Henttonen, H.; et al. Zoonotic Viruses in Three Species of Voles from Poland. Animals 2020, 10, 1820. [Google Scholar] [CrossRef]
- Billings, A.N.; Rollin, P.E.; Milazzo, M.L.; Molina, C.P.; Eyzaguirre, E.J.; Livingstone, W.; Ksiazek, T.G.; Fulhorst, C.F. Pathology of Black Creek Canal virus infection in juvenile hispid cotton rats (Sigmodon hispidus). Vector-Borne Zoonotic Dis. 2010, 10, 621–628. [Google Scholar] [CrossRef] [Green Version]
- Pedersen, A.B.; Babayan, S.A. Wild immunology. Mol. Ecol. 2011, 20, 872–880. [Google Scholar] [CrossRef]
- Jackson, J. Immunology in wild nonmodel rodents: An ecological context for studies of health and disease. Parasite Immunol. 2015, 37, 220–232. [Google Scholar] [CrossRef]
- Forbes, K.M. Ecoimmunology at spatial scales. J. Anim. Ecol. 2020, 89, 2210–2213. [Google Scholar] [CrossRef]
- Voutilainen, L.; Sironen, T.; Tonteri, E.; Bäck, A.T.; Razzauti, M.; Karlsson, M.; Wahlström, M.; Niemimaa, J.; Henttonen, H.; Lundkvist, Å. Life-long shedding of Puumala hantavirus in wild bank voles (Myodes glareolus). J. Gen. Virol. 2015, 96, 1238–1247. [Google Scholar] [CrossRef] [PubMed]
- Turner, A.K.; Beldomenico, P.M.; Bown, K.; Burthe, S.; Jackson, J.; Lambin, X.; Begon, M. Host–parasite biology in the real world: The field voles of Kielder. Parasitology 2014, 141, 997–1017. [Google Scholar] [CrossRef]
- Hansson, L.; Henttonen, H. Gradients in density variations of small rodents: The importance of latitude and snow cover. Oecologia 1985, 67, 394–402. [Google Scholar] [CrossRef]
- Krebs, C.J. Population Fluctuations in Rodents; University of Chicago Press: Chicago, IL, USA, 2013; pp. 1–22, 162–174, 306. [Google Scholar]
- Andreassen, H.P.; Sundell, J.; Ecke, F.; Halle, S.; Haapakoski, M.; Henttonen, H.; Huitu, O.; Jacob, J.; Johnsen, K.; Koskela, E. Population cycles and outbreaks of small rodents: Ten essential questions we still need to solve. Oecologia 2021, 195, 601–622. [Google Scholar] [CrossRef]
- Hanski, I.; Hansson, L.; Henttonen, H. Specialist predators, generalist predators, and the microtine rodent cycle. J. Anim. Ecol. 1991, 60, 353–367. [Google Scholar] [CrossRef]
- Korpimäki, E.; Norrdahl, K.; Klemola, T.; Pettersen, T.; Stenseth, N.C. Dynamic effects of predators on cyclic voles: Field experimentation and model extrapolation. Proc. R. Soc. London Ser. B Biol. Sci. 2002, 269, 991–997. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lambin, X.; Bretagnolle, V.; Yoccoz, N.G. Vole population cycles in northern and southern Europe: Is there a need for different explanations for single pattern? J. Anim. Ecol. 2006, 75, 340–349. [Google Scholar] [CrossRef]
- Ims, R.A.; Henden, J.-A.; Killengreen, S.T. Collapsing population cycles. Trends Ecol. Evol. 2008, 23, 79–86. [Google Scholar] [CrossRef] [Green Version]
- Korpela, K.; Helle, P.; Henttonen, H.; Korpimäki, E.; Koskela, E.; Ovaskainen, O.; Pietiäinen, H.; Sundell, J.; Valkama, J.; Huitu, O. Predator–vole interactions in northern Europe: The role of small mustelids revised. Proc. R. Soc. B Biol. Sci. 2014, 281, 20142119. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Scott, M.; Lewis, J. Population dynamics of helminth parasites in wild and laboratory rodents. Mammal. Rev. 1987, 17, 95–103. [Google Scholar] [CrossRef]
- Tompkins, D.; Begon, M. Parasites can regulate wildlife populations. Parasitol. Today 1999, 15, 311–313. [Google Scholar] [CrossRef]
- Scott, M.E. Regulation of mouse colony abundance by Heligmosomoides polygyrus. Parasitology 1987, 95, 111–124. [Google Scholar] [CrossRef]
- Pedersen, A.B.; Greives, T.J. The interaction of parasites and resources cause crashes in a wild mouse population. J. Anim. Ecol. 2008, 77, 370–377. [Google Scholar] [CrossRef]
- Haukisalmi, V.; Henttonen, H.; Tenora, F. Population dynamics of common and rare helminths in cyclic vole populations. J. Anim. Ecol. 1988, 57, 807–825. [Google Scholar] [CrossRef]
- Haukisalmi, V.; Henttonen, H. Coexistence in helminths of the bank vole Clethrionomys glareolus. I. Patterns of co-occurrence. J. Anim. Ecol. 1993, 62, 221–229. [Google Scholar] [CrossRef]
- Haukisalmi, V.; Henttonen, H. Coexistence in helminths of the bank vole Clethrionomys glareolus. II. Intestinal distribution and interspecific interactions. J. Anim. Ecol. 1993, 62, 230–238. [Google Scholar] [CrossRef]
- Stien, A.; Voutilainen, L.; Haukisalmi, V.; Fuglei, E.; Mørk, T.; Yoccoz, N.; Ims, R.; Henttonen, H. Intestinal parasites of the Arctic fox in relation to the abundance and distribution of intermediate hosts. Parasitology 2010, 137, 149. [Google Scholar] [CrossRef] [PubMed]
- Mørk, T.; Ims, R.A.; Killengreen, S.T. Rodent population cycle as a determinant of gastrointestinal nematode abundance in a low-arctic population of the red fox. Int. J. Parasitol. Parasites Wildl. 2019, 9, 36–41. [Google Scholar] [CrossRef]
- Hazel, S.; Bennett, M.; Chantrey, J.; Bown, K.; Cavanagh, R.; Jones, T.; Baxby, D.; Begon, M. A longitudinal study of an endemic disease in its wildlife reservoir: Cowpox and wild rodents. Epidemiol. Infect. 2000, 124, 551–562. [Google Scholar] [CrossRef]
- Laakkonen, J.; Oksanen, A.; Soveri, T.; Henttonen, H. Dynamics of intestinal coccidia in peak density Microtus agrestis, Microtus oeconomus and Clethrionomus glareolus populations in Finland. Ecography 1998, 21, 135–139. [Google Scholar] [CrossRef]
- Voutilainen, L.; Kallio, E.R.; Niemimaa, J.; Vapalahti, O.; Henttonen, H. Temporal dynamics of Puumala hantavirus infection in cyclic populations of bank voles. Sci. Rep. 2016, 6, 1–15. [Google Scholar]
- Sironen, T.; Henttonen, H. Many faces of rodent-borne infections in Europe. In The Impact of A Decade (2004-2015) of Research on Vector-Borne Diseases in Europe; Lancelot, R., Laurens, S., Lewer, A., Eds.; CIRAD: Montpellier, France, 2015; pp. 10–16. ISBN 978-2-87614-707-2. [Google Scholar]
- Reil, D.; Imholt, C.; Eccard, J.A.; Jacob, J. Beech fructification and bank vole population dynamics-combined analyses of promoters of human Puumala virus infections in Germany. PLoS ONE 2015, 10, e0134124. [Google Scholar] [CrossRef]
- Ezenwa, V.O.; Jolles, A.E. From host immunity to pathogen invasion: The effects of helminth coinfection on the dynamics of microparasites. Integr. Comp. Biol. 2011, 51, 540–551. [Google Scholar] [CrossRef]
- Salvador, A.R.; Guivier, E.; Xuéreb, A.; Chaval, Y.; Cadet, P.; Poulle, M.-L.; Sironen, T.; Voutilainen, L.; Henttonen, H.; Cosson, J.-F. Concomitant influence of helminth infection and landscape on the distribution of Puumala hantavirus in its reservoir, Myodes glareolus. BMC Microbiol. 2011, 11, 30. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Guivier, E.; Galan, M.; Henttonen, H.; Cosson, J.-F.; Charbonnel, N. Landscape features and helminth co-infection shape bank vole immunoheterogeneity, with consequences for Puumala virus epidemiology. Heredity 2014, 112, 274–281. [Google Scholar] [CrossRef] [Green Version]
- Charbonnel, N.; Sironen, T.; Henttonen, H.; Vapalahti, O.; Mustonen, J.; Vaheri, A. Immunogenetic factors affecting susceptibility of humans and rodents to hantaviruses and the clinical course of hantaviral disease in humans. Viruses 2014, 6, 2214–2241. [Google Scholar] [CrossRef] [Green Version]
- Jackson, J.A.; Begon, M.; Birtles, R.; Paterson, S.; Friberg, I.M.; Hall, A.; Lowe, A.; Ralli, C.; Turner, A.; Zawadzka, M. The analysis of immunological profiles in wild animals: A case study on immunodynamics in the field vole, Microtus agrestis. Mol. Ecol. 2011, 20, 893–909. [Google Scholar] [CrossRef]
- Charbonnel, N.; Galan, M.; Tatard, C.; Loiseau, A.; Diagne, C.; Dalecky, A.; Parrinello, H.; Rialle, S.; Severac, D.; Brouat, C. Differential immune gene expression associated with contemporary range expansion in two invasive rodents in Senegal. Sci. Rep. 2020, 10, 1–13. [Google Scholar] [CrossRef]
- McDevitt, A.; Vega, R.; Rambau, R.V.; Yannic, G.; Herman, J.S.; Hayden, T.; Searle, J.B. Colonization of Ireland: Revisiting ‘the pygmy shrew syndrome’using mitochondrial, Y chromosomal and microsatellite markers. Heredity 2011, 107, 548–557. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Marnell, F.; Kingston, N.; Looney, D. Ireland Red List No. 3: Terrestrial Mammals; National Parks and Wildlife Service, Department of the Environment, Heritage and Local Government: Dublin, Ireland, 2009.
- National Biodiversity Data Centre, Ireland. Bank Vole (Myodes glareolus). Available online: https://maps.biodiversityireland.ie/Species/119785 (accessed on 16 April 2021).
- Stuart, P.; Mirimin, L.; Cross, T.; Sleeman, D.; Buckley, N.J.; Telfer, S.; Birtles, R.; Kotlik, P.; Searle, J. The origin of Irish bank voles Clethrionomys glareolus assessed by mitochondrial DNA analysis. Ir. Nat. J. 2007, 28, 440–446. [Google Scholar]
- White, T.A.; Perkins, S.E.; Heckel, G.; Searle, J.B. Adaptive evolution during an ongoing range expansion: The invasive bank vole (M yodes glareolus) in Ireland. Mol. Ecol. 2013, 22, 2971–2985. [Google Scholar] [CrossRef]
- White, T.A.; Lundy, M.G.; Montgomery, W.I.; Montgomery, S.; Perkins, S.E.; Lawton, C.; Meehan, J.M.; Hayden, T.J.; Heckel, G.; Reid, N. Range expansion in an invasive small mammal: Influence of life-history and habitat quality. Biol. Invasions 2012, 14, 2203–2215. [Google Scholar] [CrossRef]
- Perkins, S.E.; White, T.A.; Pascoe, E.L.; Gillingham, E.L. Parasite community dynamics in an invasive vole–From focal introduction to wave front. Int. J. Parasitol. Parasites Wildl. 2017, 6, 412–419. [Google Scholar] [CrossRef]
- O’Sullivan, H.M.; Smal, C.M.; Fairley, J.S. A Study of Parasite Infestations in Populations of Small Rodents (Apodemus Sylvaticus and Clethrionomys Glareolus) on Ross Island, Killarney. J. Life Sci R. Dubl. Soc. 1984, 5, 29–42. [Google Scholar]
- Montgomery, S.; Montgomery, W. Cyclic and non-cyclic dynamics in populations of the helminth parasites of wood mice, Apodemus sylvaticus. J. Helminthol. 1988, 62, 78–90. [Google Scholar] [CrossRef] [PubMed]
- Montgomery, S.; Montgomery, W. Spatial and temporal variation in the infracommunity structure of helminths of Apodemus sylvaticus (Rodentia: Muridae). Parasitology 1989, 98, 145–150. [Google Scholar] [CrossRef]
- Montgomery, S.; Montgomery, W. Structure, stability and species interactions in helminth communities of wood mice, Apodemus sylvaticus. Int. J. Parasitol. 1990, 20, 225–242. [Google Scholar] [CrossRef]
- Loxton, K.C.; Lawton, C.; Stafford, P.; Holland, C.V. Reduced helminth parasitism in the introduced bank vole (Myodes glareolus): More parasites lost than gained. Int. J. Parasitol. Parasites Wildl. 2016, 5, 175–183. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Loxton, K.C.; Lawton, C.; Stafford, P.; Holland, C.V. Parasite dynamics in an invaded ecosystem: Helminth communities of native wood mice are impacted by the invasive bank vole. Parasitology 2017, 144, 1476–1489. [Google Scholar] [CrossRef]
- Haukisalmi, V.; Henttonen, H. Variability of helminth assemblages and populations in the bank vole Clethrionomys glareolus. Pol. J. Ecol. 2000, 48, 219–231. [Google Scholar]
- Telfer, S.; Bown, K.J.; Sekules, R.; Begon, M.; Hayden, T.; Birtles, R. Disruption of a host-parasite system following the introduction of an exotic host species. Parasitology 2005, 130, 661–668. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kronfeld-Schor, N.; Stevenson, T.; Nickbakhsh, S.; Schernhammer, E.; Dopico, X.; Dayan, T.; Martinez, M.; Helm, B. Drivers of infectious disease seasonality: Potential implications for COVID-19. J. Biol. Rhythm. 2021, 36, 35–54. [Google Scholar] [CrossRef]
- Montgomery, W.I.; Lundy, M.G.; Reid, N. ‘Invasional meltdown’: Evidence for unexpected consequences and cumulative impacts of multispecies invasions. Biol. Invasions 2012, 14, 1111–1125. [Google Scholar] [CrossRef]
- Tosh, D.G.; Lusby, J.; Montgomery, W.I.; O’halloran, J. First record of greater white-toothed shrew Crocidura russula in Ireland. Mammal. Rev. 2008, 38, 321–326. [Google Scholar] [CrossRef]
- Cornette, R.; Tresset, A.; Houssin, C.; Pascal, M.; Herrel, A. Does bite force provide a competitive advantage in shrews? The case of the greater white-toothed shrew. Biol. J. Linn. Soc. 2015, 114, 795–807. [Google Scholar] [CrossRef] [Green Version]
- White, T.A.; Perkins, S.E. The ecoimmunology of invasive species. Funct. Ecol. 2012, 26, 1313–1323. [Google Scholar] [CrossRef]
- Montgomery, W.; Dowie, M. The distribution of the wood mouse Apodemus sylvaticus and the house mouse Mus domesticus on farmland in north-east Ireland. Ir. Nat. J. 1993, 24, 199–203. [Google Scholar]
- National Biodiversity Data Centre, Ireland. Bank Vole (Myodes glareolus), Image. Available online: https://maps.biodiversityireland.ie/Species/TerrestrialDistributionMapPrintSize/119785 (accessed on 16 March 2021).
- Haukisalmi, V.; Henttonen, H.; Batzli, G.O. Helminth parasitism in the voles Microtus oeconomus and M. miurus on the North Slope of Alaska: Host specificity and the effects of host sex, age and breeding status. Ann. Zool. Fenn. JSTOR 1995, 32, 193–201. [Google Scholar]
- Fasola, M.; Canova, L. Asymmetrical competition between the bank vole and the wood mouse, a removal experiment. Acta Theriol. 2000, 45, 353–366. [Google Scholar] [CrossRef]
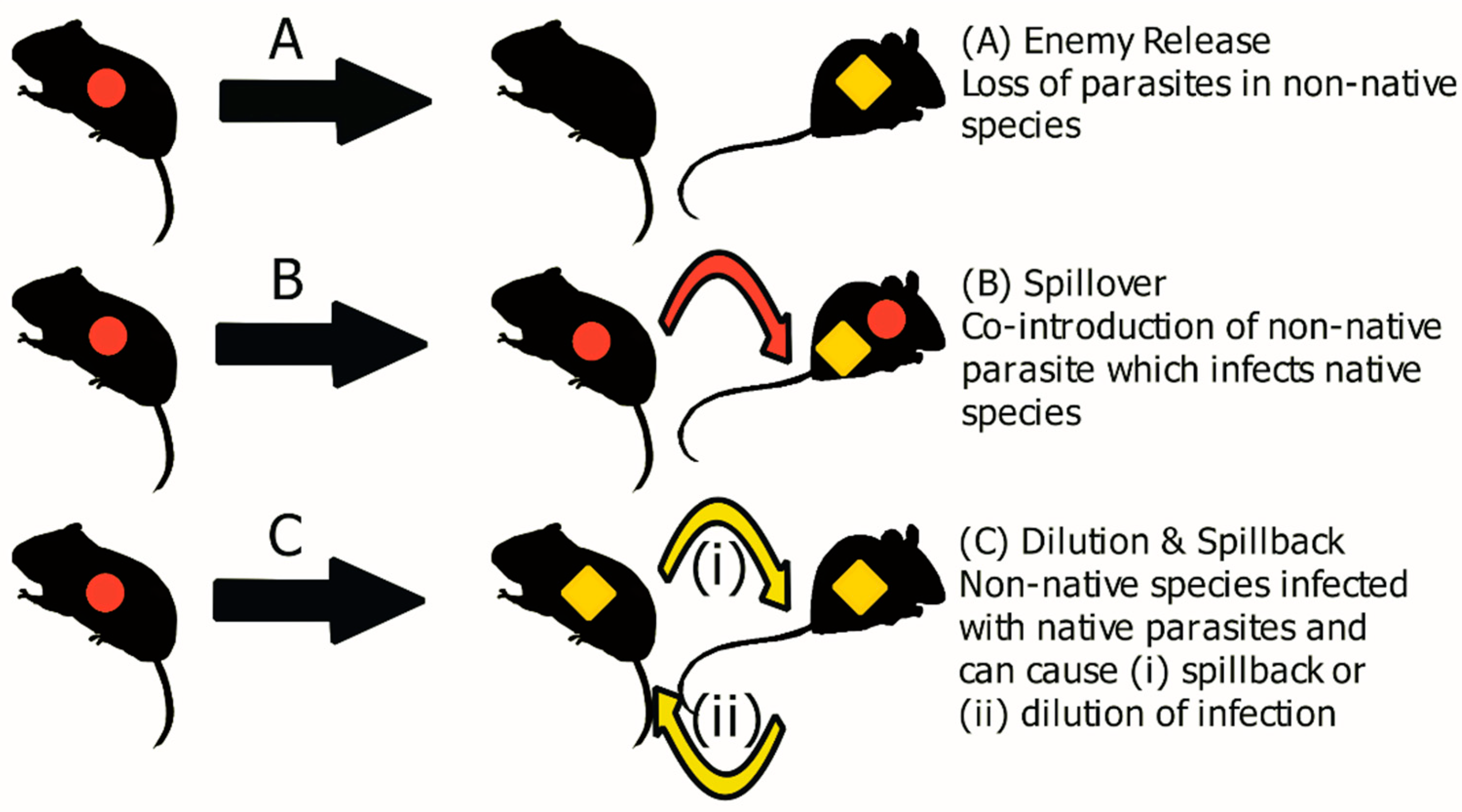


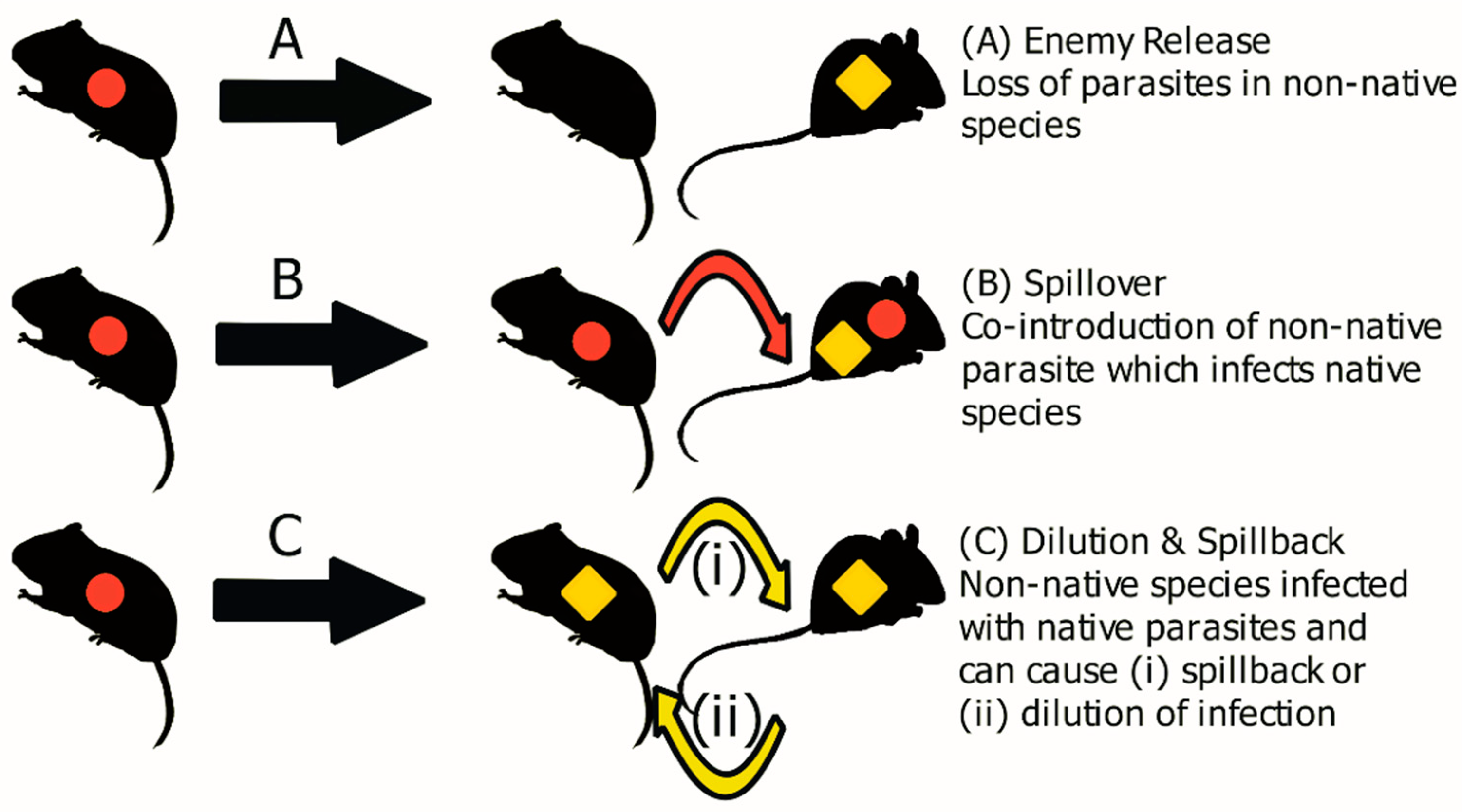{kind=link}
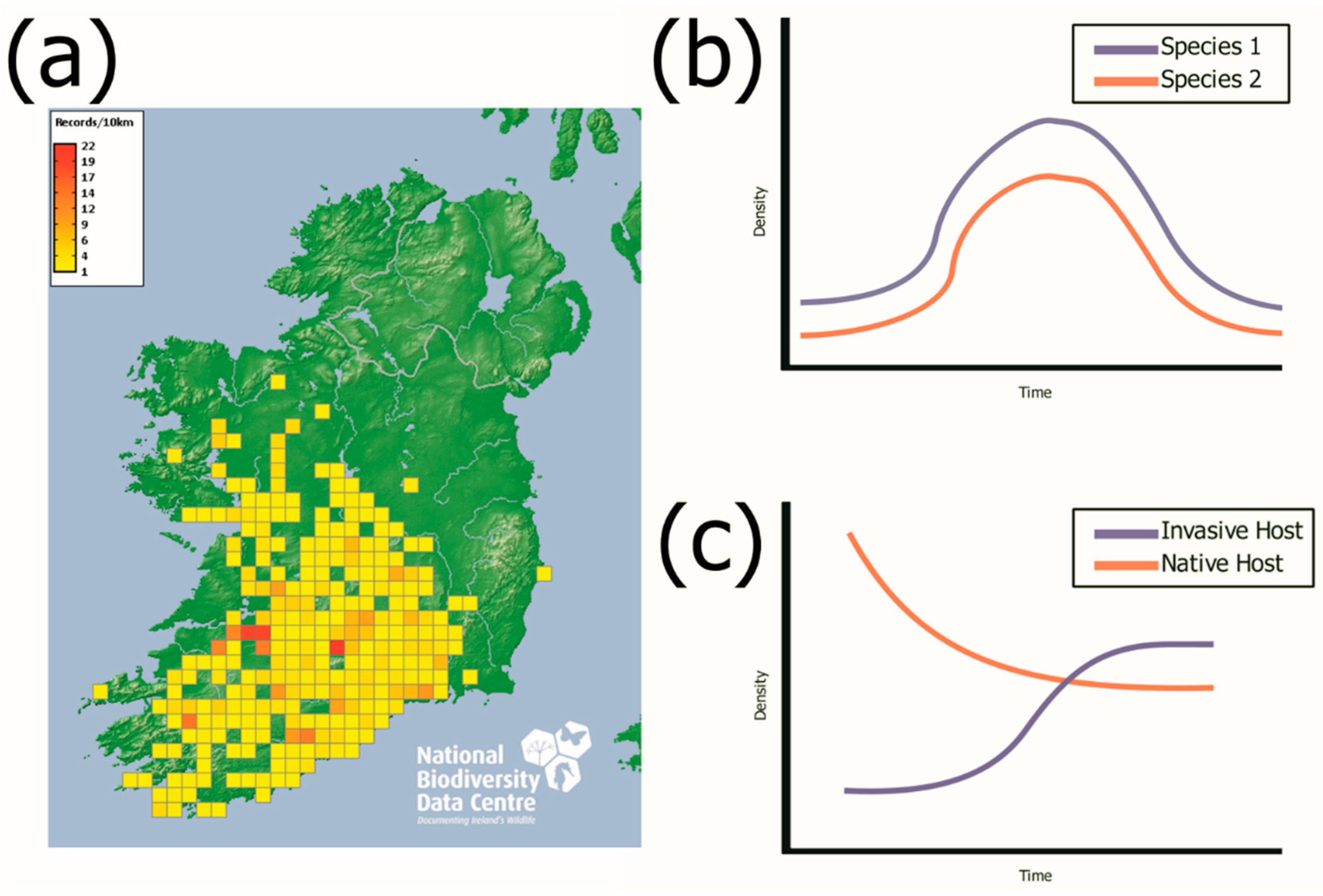{kind=link}
| A. sylvaticus | ||||
|---|---|---|---|---|
| Taxon | O’Sullivan, Smal and Fairley (1982) | Montgomery and Montgomery (1988; 1990) | Loxton et al. (2016; 2017) | Stuart et al. (2020) |
| Nematoda | Syphacia stroma Capillaria muris-sylvatici Trichuris muris Heligmosomoides polygyrus (Nematospiroides dubius) | Syphacia stroma Capillaria murissylvatici Trichuris muris Heligmosomoides polygyrus (Nematospiroides dubius) | Syphacia stroma Aonchotheca murissylvatici Trichuris muris Heligmosomoides polygyrus | Syphacia stroma Aonchotheca murissylvatici Trichuris muris Heligmosomoides polygyrus |
| Cestoda | Hymenolepis straminea Skrjabinotaenia lobata (Catenotaenia lobata) Hydatigera taeniaeformis (Taenia taeniaeformis) | Hymenolepis hibernia Skrjabinotaenia lobata (Catenotaenia lobata) Hydatigera taeniaeformis (Taenia taeniaeformis) | Hymenolepis hibernia Skrjabinotaenia lobata Taenia martis Hydatigera taeniaeformis(Taenia taeniaeformis) | Hymenolepis hibernia Hymenolepis sp. Skrjabinotaenia lobata Taenia martis Taenia polyacantha Hydatigera taeniaeformis |
| Trematoda | Brachylaimus recurvum Corrigia vitta Plagiorchis muris | Brachylaima recurva Corrigia vitta | Brachylaemus recurvum Corrigia vitta | Brachylaemus recurvum Corrigia vitta |
| Total Sp. | 10 | 9 | 10 | 12 |
| M. glareolus | ||||
| Taxon | O’Sullivan, Smal and Fairley (1982) | Loxton et al. (2016; 2017) | Stuart et al. (2020) | |
| Nematoda | Capillaria muris-sylvatici Trichuris muris | Aonchotheca murissylvatici Aspiculuris tianjinensis | Aonchotheca murissylvatici Aspiculuris tianjinensis | |
| Cestoda | Taenia martis | Taenia martis Taenia polyacantha | ||
| Trematoda | Corrigia vitta | |||
| Total Sp. | 3 | 3 | 4 | |
| Current Knowledge Base: |
| Long history of using rodent models in laboratory and wild situations to study disease dynamics worldwide. |
| Single introduction point and no current eradication attempts. |
| Presence of baseline parasitological data on the Irish M. glareolus biological invasion; |
| including long established invaded sites, recently invaded sites and uninvaded sites. |
| Main Research Questions: |
| How parasite dynamics and immune gene expression change during a biological invasion. |
| How This Can Be Addressed: |
| Data collected during previous studies can be used as a baseline for future studies. |
| The addition of multiple invaders can further investigate how parasite dynamics change during a biological invasion. |
| Application of knowledge gained to similar rodent systems elsewhere. |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
McManus, A.; Holland, C.V.; Henttonen, H.; Stuart, P. The Invasive Bank Vole (Myodes glareolus): A Model System for Studying Parasites and Ecoimmunology during a Biological Invasion. Animals 2021, 11, 2529. https://doi.org/10.3390/ani11092529
McManus A, Holland CV, Henttonen H, Stuart P. The Invasive Bank Vole (Myodes glareolus): A Model System for Studying Parasites and Ecoimmunology during a Biological Invasion. Animals. 2021; 11(9):2529. https://doi.org/10.3390/ani11092529
Chicago/Turabian StyleMcManus, Andrew, Celia V. Holland, Heikki Henttonen, and Peter Stuart. 2021. "The Invasive Bank Vole (Myodes glareolus): A Model System for Studying Parasites and Ecoimmunology during a Biological Invasion" Animals 11, no. 9: 2529. https://doi.org/10.3390/ani11092529
APA StyleMcManus, A., Holland, C. V., Henttonen, H., & Stuart, P. (2021). The Invasive Bank Vole (Myodes glareolus): A Model System for Studying Parasites and Ecoimmunology during a Biological Invasion. Animals, 11(9), 2529. https://doi.org/10.3390/ani11092529