
Chronological Trends and Mercury Bioaccumulation in an Aquatic Semiarid Ecosystem under a Global Climate Change Scenario in the Northeastern Coast of Brazil

 ,
,  and
and
Simple Summary
Abstract
1. Introduction
2. Materials and Methods
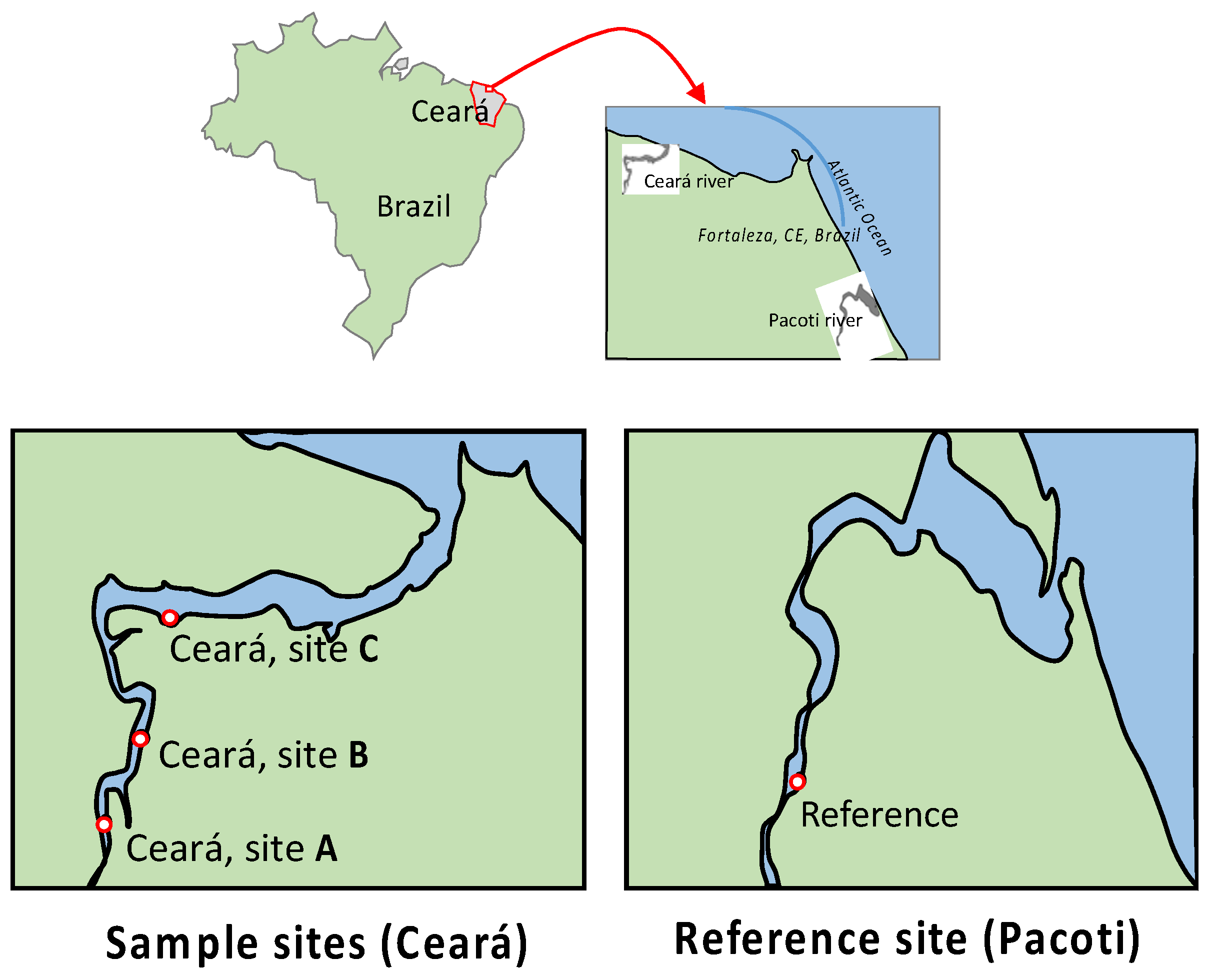
2.1. Study Area
2.2. Study Sampling Design
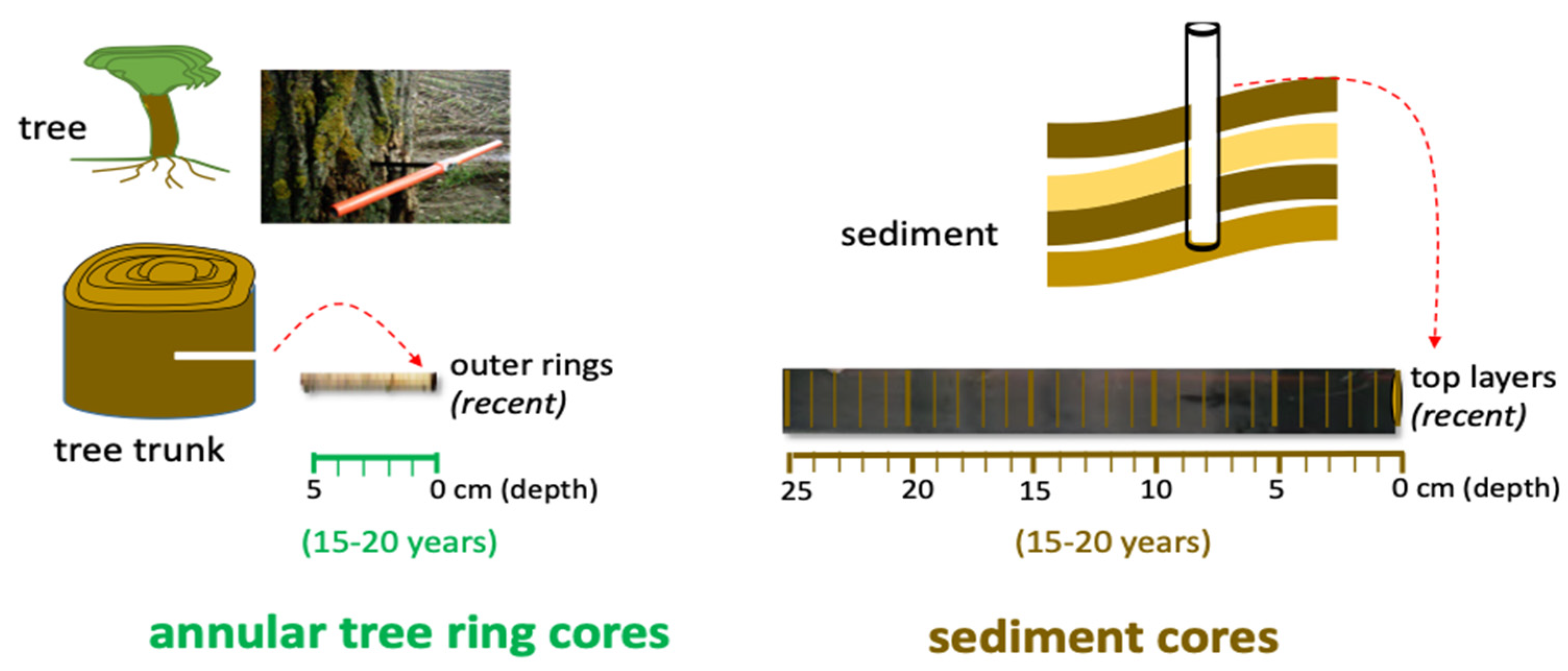
2.3. Rhizophora mangle L. (Red Mangrove) Sampling and Treatment
2.4. Sphoeroides Testudineus Sampling and Treatment
2.5. Surface Sediment and Water Sampling and Treatment
2.6. Mercury Analysis and Data Quality Control
2.7. Biota–Sediment Accumulation Factor Determination
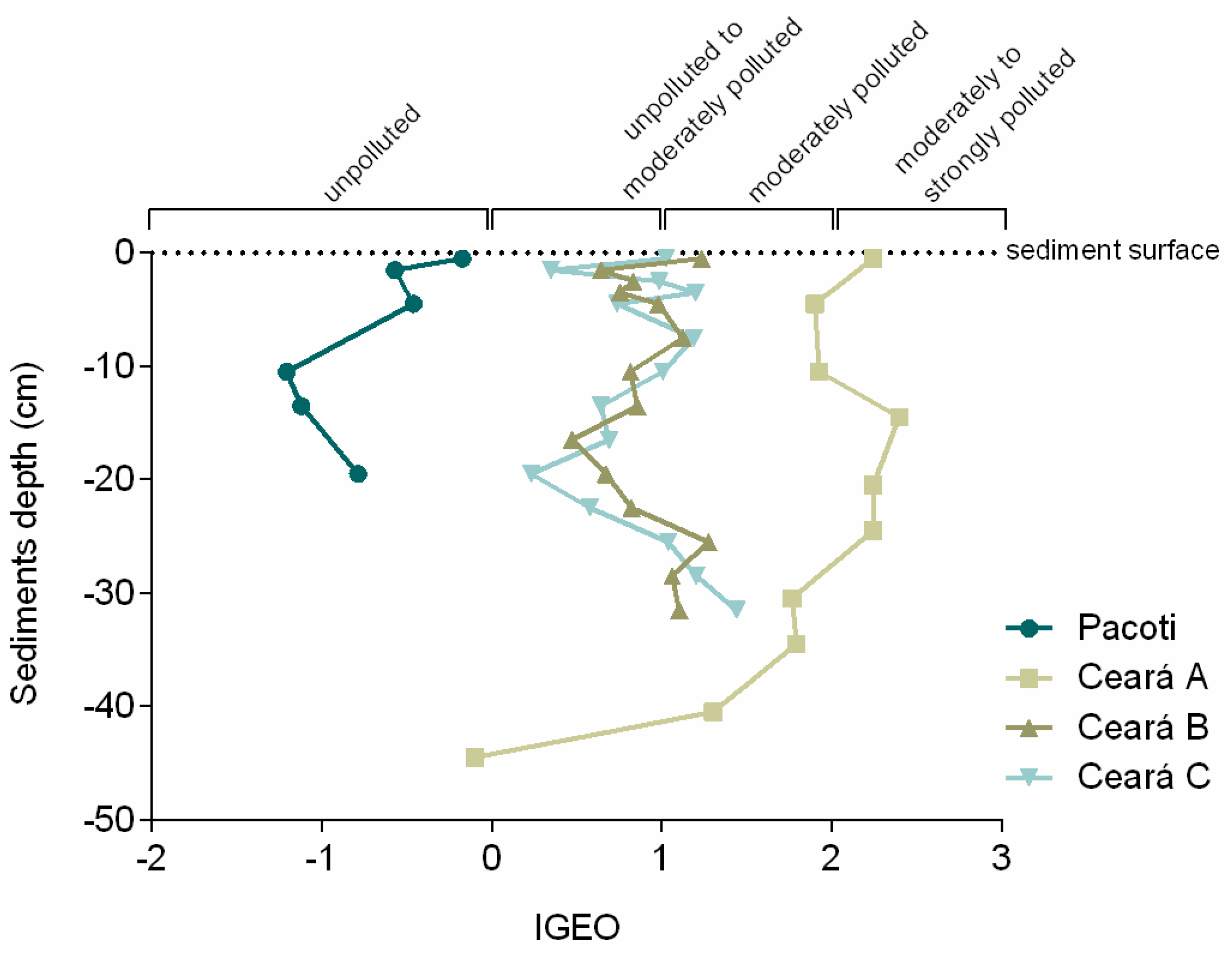
2.8. Geoaccumulation Index (Igeo)
2.9. Statistical Analyses
3. Results
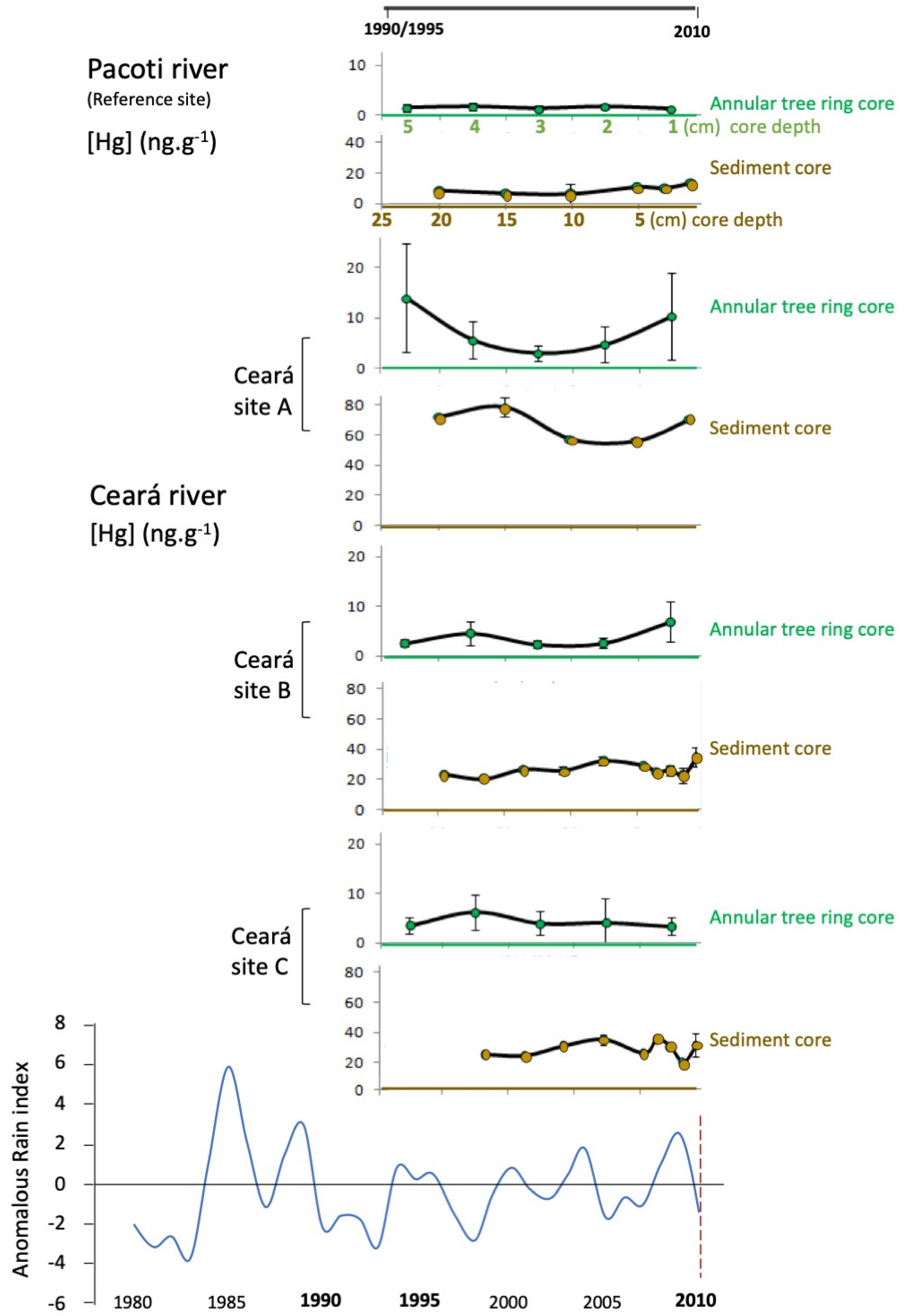
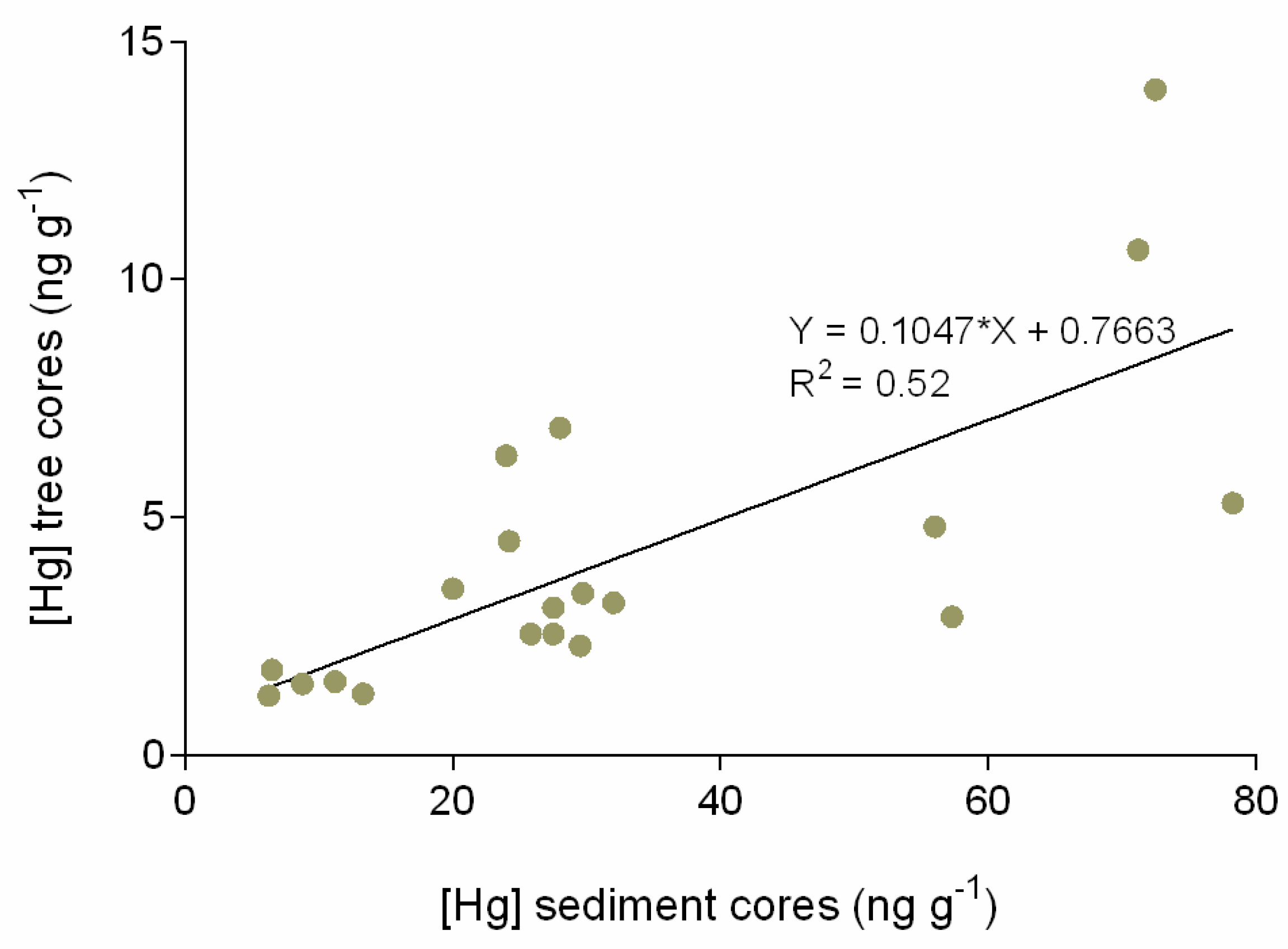
3.1. Mercury in Tree and Sediment Cores
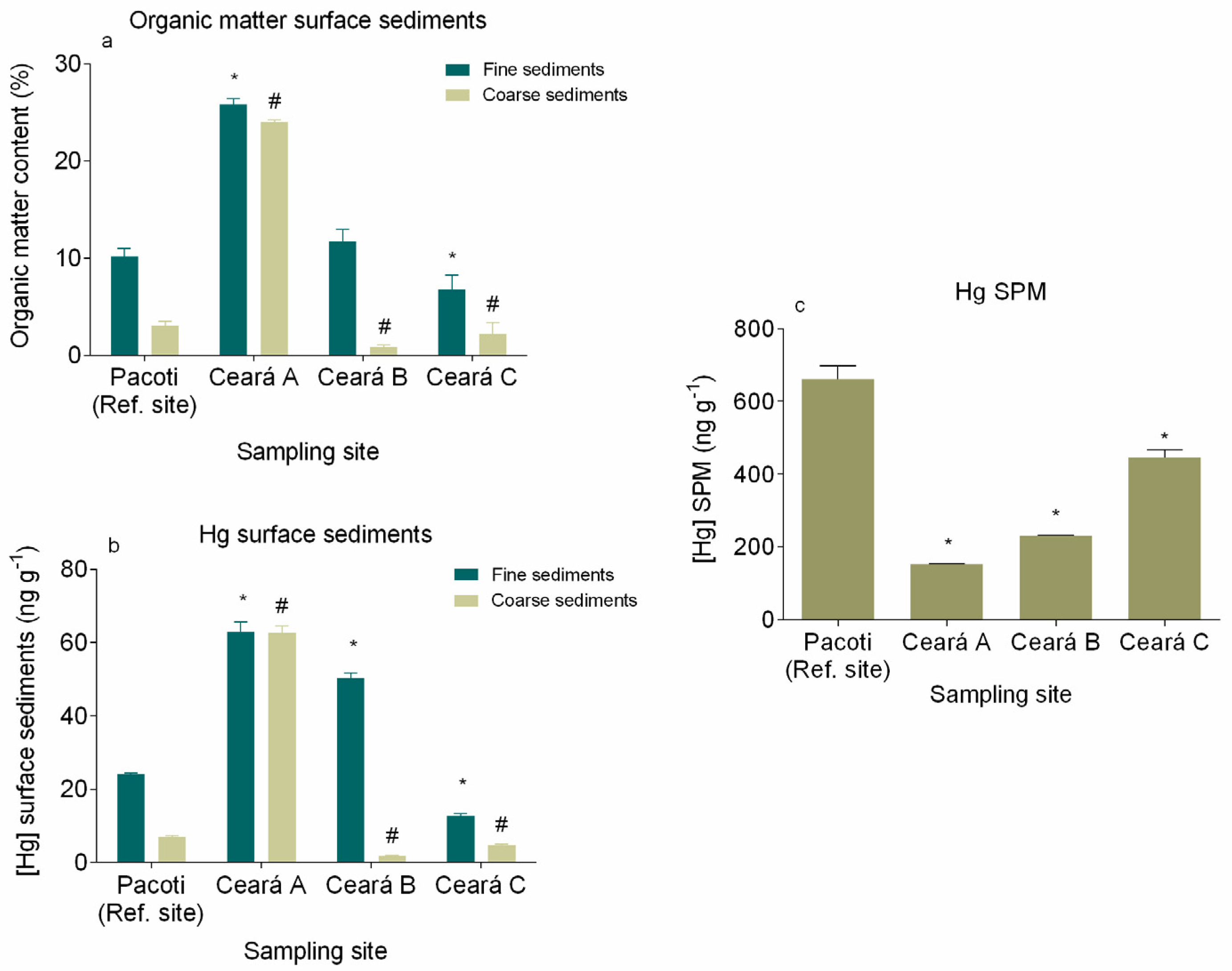
3.2. Mercury in Surface Sediments and SPM
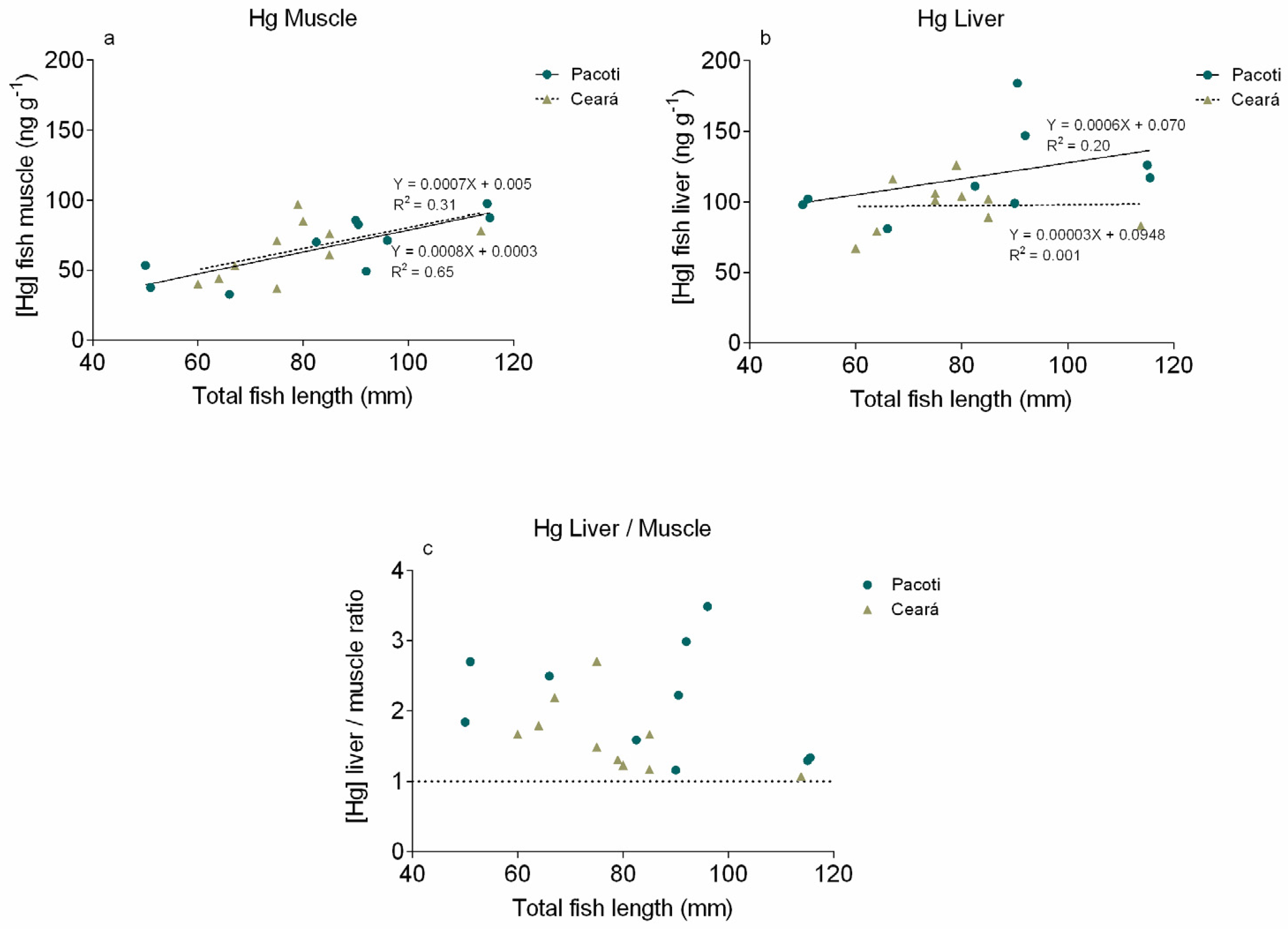
3.3. Mercury in Sphoeroides testudineus Tissues
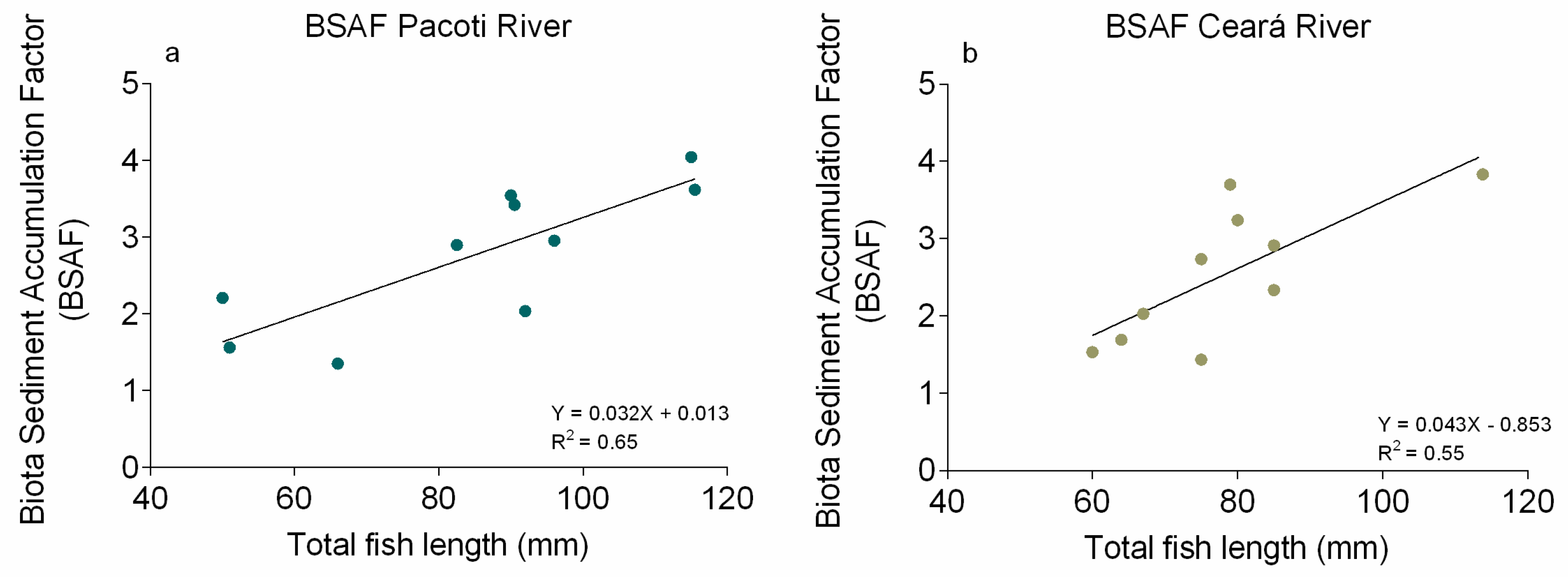
3.4. Biota–Sediment Accumulation Factor Determination
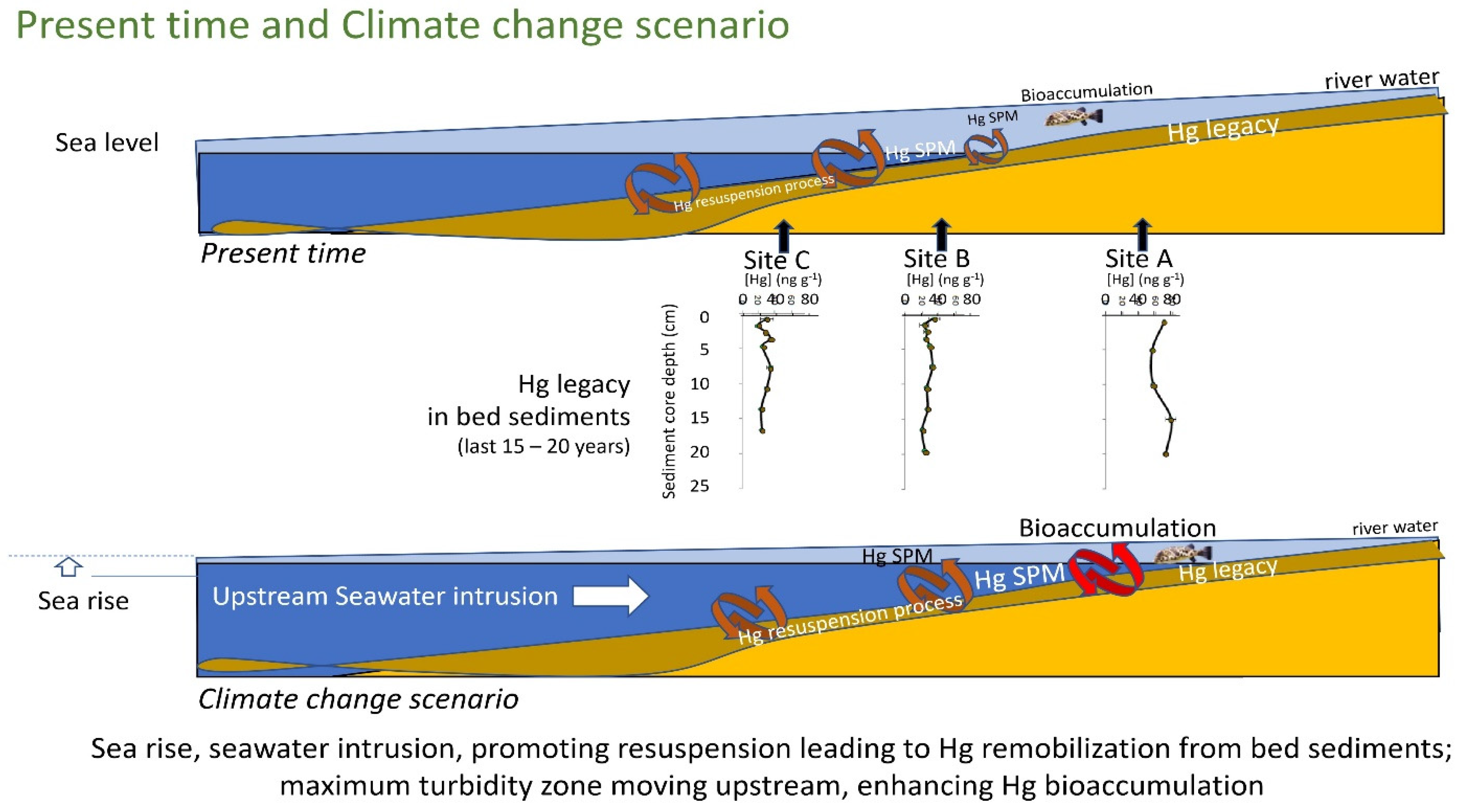
3.5. Conceptual Model of Present Time and Climate Change Scenario
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Obrist, D.; Kirk, J.L.; Zhang, L.; Sunderland, E.M.; Jiskra, M.; Selin, N.E. A review of global environmental mercury processes in response to human and natural perturbations: Changes of emissions, climate, and land use. Ambio 2018, 47, 116–140. [Google Scholar] [CrossRef]
- Driscoll, C.T.; Mason, R.P.; Chan, H.M.; Jacob, D.J.; Pirrone, N. Mercury as a Global Pollutant: Sources, Pathways, and Effects. Environ. Sci. Technol. 2013, 47, 4967–4983. [Google Scholar] [CrossRef] [PubMed]
- Amos, H.M.; Sonke, J.E.; Obrist, D.; Robins, N.; Hagan, N.; Horowitz, H.M.; Mason, R.P.; Witt, M.; Hedgecock, I.M.; Corbitt, E.S. Observational and modeling constraints on global anthropogenic enrichment of mercury. Environ. Sci. Technol. 2015, 49, 4036–4047. [Google Scholar] [CrossRef]
- Streets, D.G.; Horowitz, H.M.; Jacob, D.J.; Lu, Z.; Levin, L.; Ter Schure, A.F.; Sunderland, E.M. Total mercury released to the environment by human activities. Environ. Sci. Technol. 2017, 51, 5969–5977. [Google Scholar] [CrossRef]
- Hsu-Kim, H.; Eckley, C.S.; Achá, D.; Feng, X.; Gilmour, C.C.; Jonsson, S.; Mitchell, C.P. Challenges and opportunities for managing aquatic mercury pollution in altered landscapes. Ambio 2018, 47, 141–169. [Google Scholar] [CrossRef] [PubMed]
- Evers, D. The effects of methylmercury on wildlife: A comprehensive review and approach for interpretation. Encycl. Anthr. 2018, 5, 181–194. [Google Scholar]
- Eagles-Smith, C.A.; Silbergeld, E.K.; Basu, N.; Bustamante, P.; Diaz-Barriga, F.; Hopkins, W.A.; Kidd, K.A.; Nyland, J.F. Modulators of mercury risk to wildlife and humans in the context of rapid global change. Ambio 2018, 47, 170–197. [Google Scholar] [CrossRef]
- Morcillo, P.; Esteban, M.A.; Cuesta, A. Mercury and its toxic effects on fish. AIMS Environ. Sci. 2017, 4, 386–402. [Google Scholar] [CrossRef]
- Scheuhammer, A.M.; Meyer, M.W.; Sandheinrich, M.B.; Murray, M.W. Effects of environmental methylmercury on the health of wild birds, mammals, and fish. Ambio 2007, 36, 12–19. [Google Scholar] [CrossRef]
- Dietz, R.; Sonne, C.; Basu, N.; Braune, B.; O’Hara, T.; Letcher, R.J.; Scheuhammer, T.; Andersen, M.; Andreasen, C.; Andriashek, D. What are the toxicological effects of mercury in Arctic biota? Sci. Total Environ. 2013, 443, 775–790. [Google Scholar] [CrossRef]
- Braune, B.; Chételat, J.; Amyot, M.; Brown, T.; Clayden, M.; Evans, M.; Fisk, A.; Gaden, A.; Girard, C.; Hare, A. Mercury in the marine environment of the Canadian Arctic: Review of recent findings. Sci. Total Environ. 2015, 509, 67–90. [Google Scholar] [CrossRef]
- Jackson, A.K.; Evers, D.C.; Adams, E.M.; Cristol, D.A.; Eagles-Smith, C.; Edmonds, S.T.; Gray, C.E.; Hoskins, B.; Lane, O.P.; Sauer, A. Songbirds as sentinels of mercury in terrestrial habitats of eastern North America. Ecotoxicology 2015, 24, 453–467. [Google Scholar] [CrossRef] [PubMed]
- Ackerman, J.T.; Eagles-Smith, C.A.; Herzog, M.P.; Hartman, C.A.; Peterson, S.H.; Evers, D.C.; Jackson, A.K.; Elliott, J.E.; Vander Pol, S.S.; Bryan, C.E. Avian mercury exposure and toxicological risk across western North America: A synthesis. Sci. Total Environ. 2016, 568, 749–769. [Google Scholar] [CrossRef]
- Crowe, W.; Allsopp, P.J.; Watson, G.E.; Magee, P.J.; Strain, J.J.; Armstrong, D.J.; Ball, E.; McSorley, E.M. Mercury as an environmental stimulus in the development of autoimmunity—A systematic review. Autoimmun. Rev. 2017, 16, 72–80. [Google Scholar] [CrossRef] [PubMed]
- Hawley, D.M.; Hallinger, K.K.; Cristol, D.A. Compromised immune competence in free-living tree swallows exposed to mercury. Ecotoxicology 2009, 18, 499–503. [Google Scholar] [CrossRef]
- Lewis, C.A.; Cristol, D.A.; Swaddle, J.P.; Varian-Ramos, C.W.; Zwollo, P. Decreased immune response in zebra finches exposed to sublethal doses of mercury. Arch. Environ. Contam. Toxicol. 2013, 64, 327–336. [Google Scholar] [CrossRef] [PubMed]
- Hui, L.L.; Chan, M.H.M.; Lam, H.S.; Chan, P.H.Y.; Kwok, K.M.; Chan, I.H.S.; Li, A.M.; Fok, T.F. Impact of fetal and childhood mercury exposure on immune status in children. Environ. Res. 2016, 144, 66–72. [Google Scholar] [CrossRef]
- Zhang, Q.-F.; Li, Y.-W.; Liu, Z.-H.; Chen, Q.-L. Exposure to mercuric chloride induces developmental damage, oxidative stress and immunotoxicity in zebrafish embryos-larvae. Aquat. Toxicol. 2016, 181, 76–85. [Google Scholar] [CrossRef]
- Tchounwou, P.B.; Ayensu, W.K.; Ninashvili, N.; Sutton, D. Environmental exposure to mercury and its toxicopathologic implications for public health. Environ. Toxicol. Int. J. 2003, 18, 149–175. [Google Scholar] [CrossRef]
- Basu, N.; Stamler, C.J.; Loua, K.M.; Chan, H.M. An interspecies comparison of mercury inhibition on muscarinic acetylcholine receptor binding in the cerebral cortex and cerebellum. Toxicol. Appl. Pharmacol. 2005, 205, 71–76. [Google Scholar] [CrossRef]
- Clarkson, T.W.; Magos, L. The Toxicology of Mercury and Its Chemical Compounds. Crit. Rev. Toxicol. 2006, 36, 609–662. [Google Scholar] [CrossRef]
- Mergler, D.; Anderson, H.A.; Chan, L.H.; Mahaffey, K.R.; Murray, M.; Sakamoto, M.; Stern, A.H. Methylmercury exposure and health effects in humans: A worldwide concern. Ambio 2007, 36, 3–11. [Google Scholar] [CrossRef]
- Depew, D.C.; Basu, N.; Burgess, N.M.; Campbell, L.M.; Evers, D.C.; Grasman, K.A.; Scheuhammer, A.M. Derivation of screening benchmarks for dietary methylmercury exposure for the common loon (Gavia immer): Rationale for use in ecological risk assessment. Environ. Toxicol. Chem. 2012, 31, 2399–2407. [Google Scholar] [CrossRef] [PubMed]
- Bridges, K.N.; Soulen, B.K.; Overturf, C.L.; Drevnick, P.E.; Roberts, A.P. Embryotoxicity of maternally transferred methylmercury to fathead minnows (Pimephales promelas). Environ. Toxicol. Chem. 2016, 35, 1436–1441. [Google Scholar] [CrossRef] [PubMed]
- Landler, L.; Painter, M.S.; Coe, B.H.; Youmans, P.W.; Hopkins, W.A.; Phillips, J.B. High levels of maternally transferred mercury disrupt magnetic responses of snapping turtle hatchlings (Chelydra serpentina). Environ. Pollut. 2017, 228, 19–25. [Google Scholar] [CrossRef] [PubMed]
- Wada, H.; Cristol, D.A.; McNabb, F.A.; Hopkins, W.A. Suppressed adrenocortical responses and thyroid hormone levels in birds near a mercury-contaminated river. Environ. Sci. Technol. 2009, 43, 6031–6038. [Google Scholar] [CrossRef] [PubMed]
- Meyer, E.; Eagles-Smith, C.A.; Sparling, D.; Blumenshine, S. Mercury exposure associated with altered plasma thyroid hormones in the declining western pond turtle (Emys marmorata) from California mountain streams. Environ. Sci. Technol. 2014, 48, 2989–2996. [Google Scholar] [CrossRef] [PubMed]
- Klaper, R.; Rees, C.B.; Drevnick, P.; Weber, D.; Sandheinrich, M.; Carvan, M.J. Gene expression changes related to endocrine function and decline in reproduction in fathead minnow (Pimephales promelas) after dietary methylmercury exposure. Environ. Health Perspect. 2006, 114, 1337–1343. [Google Scholar] [CrossRef]
- Burgess, N.M.; Meyer, M.W. Methylmercury exposure associated with reduced productivity in common loons. Ecotoxicology 2008, 17, 83–91. [Google Scholar] [CrossRef] [PubMed]
- Bergeron, C.M.; Hopkins, W.A.; Todd, B.D.; Hepner, M.J.; Unrine, J.M. Interactive effects of maternal and dietary mercury exposure have latent and lethal consequences for amphibian larvae. Environ. Sci. Technol. 2011, 45, 3781–3787. [Google Scholar] [CrossRef]
- Hopkins, B.C.; Willson, J.D.; Hopkins, W.A. Mercury exposure is associated with negative effects on turtle reproduction. Environ. Sci. Technol. 2013, 47, 2416–2422. [Google Scholar] [CrossRef]
- Tartu, S.; Goutte, A.; Bustamante, P.; Angelier, F.; Moe, B.; Clément-Chastel, C.; Bech, C.; Gabrielsen, G.W.; Bustnes, J.O.; Chastel, O. To breed or not to breed: Endocrine response to mercury contamination by an Arctic seabird. Biol. Lett. 2013, 9, 20130317. [Google Scholar] [CrossRef] [PubMed]
- Wiener, J.G.; Krabbenhoft, D.P.; Heinz, G.H.; Scheuhammer, A.M. Ecotoxicology of mercury. Handb. Ecotoxicol. 2003, 2, 409–463. [Google Scholar]
- USEPA. Mercury Study Report to Congress Volume VI: An Ecological Assessment for Anthropogenic Mercury Emissions in the United States; U.S. Environmental Protection Agency (EPA): Washington, DC, USA, 1997. [Google Scholar]
- Shore, R.F.; Pereira, M.G.; Walker, L.A.; Thompson, D.R. Mercury in Nonmarine Birds and Mammals. In Environmental Contaminants in Biota: Interpreting Tissue Concentrations, 2nd ed.; Nelson, B.W., James, P.M., Eds.; CRC Press: Boca Raton, FL, USA, 2011; pp. 609–624. [Google Scholar]
- Vieira, H.C.; de Abreu, S.N.; Morgado, F. Field Caging Transplants of Fish for Assessment and Monitoring in Aquatic Toxicology. In Life Below Water; Leal Filho, W., Azul, A.M., Brandli, L., Lange Salvia, A., Wall, T., Eds.; Springer International Publishing: Cham, Switzerland, 2020; pp. 1–15. [Google Scholar]
- Clackett, S.P.; Porter, T.J.; Lehnherr, I. 400-year record of atmospheric mercury from tree-rings in Northwestern Canada. Environ. Sci. Technol. 2018, 52, 9625–9633. [Google Scholar] [CrossRef]
- Enrico, M.; Le Roux, G.; Heimbürger, L.-E.; Van Beek, P.; Souhaut, M.; Chmeleff, J.; Sonke, J.E. Holocene atmospheric mercury levels reconstructed from peat bog mercury stable isotopes. Environ. Sci. Technol. 2017, 51, 5899–5906. [Google Scholar] [CrossRef] [PubMed]
- de Lacerda, L.D.; Turcq, B.; Sifeddine, A.; Cordeiro, R.C. Mercury accumulation rates in Caço Lake, NE Brazil during the past 20.000 years. J. S. Am. Earth Sci. 2017, 77, 42–50. [Google Scholar] [CrossRef]
- Gallego, J.L.R.; Ortiz, J.E.; Sierra, C.; Torres, T.; Llamas, J. Multivariate study of trace element distribution in the geological record of Roñanzas Peat Bog (Asturias, N. Spain). Paleoenvironmental evolution and human activities over the last 8000 cal yr BP. Sci. Total Environ. 2013, 454, 16–29. [Google Scholar] [CrossRef]
- Li, F.; Ma, C.; Zhang, P. Mercury deposition, climate change and anthropogenic activities: A review. Front. Earth Sci. 2020, 8, 316. [Google Scholar] [CrossRef]
- Li, Y.; Ma, C.; Zhu, C.; Huang, R.; Zheng, C. Historical anthropogenic contributions to mercury accumulation recorded by a peat core from Dajiuhu montane mire, central China. Environ. Pollut. 2016, 216, 332–339. [Google Scholar] [CrossRef]
- Pérez-Rodríguez, M.; Horák-Terra, I.; Rodríguez-Lado, L.; Aboal, J.s.R.; Martinez Cortizas, A. Long-term (~57 ka) controls on mercury accumulation in the souther hemisphere reconstructed using a peat record from Pinheiro Mire (Minas Gerais, Brazil). Environ. Sci. Technol. 2015, 49, 1356–1364. [Google Scholar] [CrossRef]
- Guédron, S.; Ledru, M.-P.; Escobar-Torrez, K.; Develle, A.; Brisset, E. Enhanced mercury deposition by Amazonian orographic precipitation: Evidence from high-elevation Holocene records of the Lake Titicaca region (Bolivia). Palaeogeogr. Palaeoclimatol. Palaeoecol. 2018, 511, 577–587. [Google Scholar] [CrossRef]
- WHO/UNICEF. Progress on Sanitation and Drinking Water: 2015 Update and MDG Assessment; World Health Organization: Geneva, Switzerland, 2015. [Google Scholar]
- UN-Water. Compendium of Water Quality Regulatory Frameworks: Which Water for Which Use? UN-Water: Geneva, Switzerland, 2015. [Google Scholar]
- USEPA. Mercury; initial inventory report of supply, use, and trade. Fed. Regist. 2017, 82, 15522–15523. [Google Scholar]
- Dijkstra, J.A.; Westerman, E.L.; Harris, L.G. The effects of climate change on species composition, succession and phenology: A case study. Glob. Chang. Biol. 2011, 17, 2360–2369. [Google Scholar] [CrossRef]
- Lacerda, L.D.; Dias, F.J.; Marins, R.V.; Soares, T.M.; Godoy, J.M.O.; Godoy, M.L.D. Pluriannual watershed discharges of Hg into a tropical semi-arid estuary of the Jaguaribe River, NE Brazil. J. Braz. Chem. Soc. 2013, 24, 1719–1731. [Google Scholar] [CrossRef]
- Marengo, J.A.; Alves, L.M.; Alvala, R.; Cunha, A.P.; Brito, S.; Moraes, O.L. Climatic characteristics of the 2010-2016 drought in the semiarid Northeast Brazil region. An. Acad. Bras. Ciênc. 2018, 90, 1973–1985. [Google Scholar] [CrossRef]
- Marins, R.; Lacerda, L.; Mounier, S.; Paraquetti, H.; Marques, W. Caracterização hidroquímica, distribuição e especiação de mercúrio nos estuários dos Rios Ceará e Pacotí, Região Metropolitana de Fortaleza, Ceará, Brasil. Geochim. Bras. 2002, 16, 37–48. [Google Scholar]
- Obrist, D.; Agnan, Y.; Jiskra, M.; Olson, C.L.; Colegrove, D.P.; Hueber, J.; Moore, C.W.; Sonke, J.E.; Helmig, D. Tundra uptake of atmospheric elemental mercury drives Arctic mercury pollution. Nature 2017, 547, 201–204. [Google Scholar] [CrossRef]
- Carignan, J.; Sonke, J. The effect of atmospheric mercury depletion events on the net deposition flux around Hudson Bay, Canada. Atmos. Environ. 2010, 44, 4372–4379. [Google Scholar] [CrossRef]
- Dai, J.; Balish, R.; Meagher, R.B.; Merkle, S.A. Development of transgenic hybrid sweetgum (Liquidambar styraciflua × L. formosana) expressing γ-glutamylcysteine synthetase or mercuric reductase for phytoremediation of mercury pollution. New For. 2009, 38, 35–52. [Google Scholar] [CrossRef]
- Stern, G.A.; Macdonald, R.W.; Outridge, P.M.; Wilson, S.; Chetelat, J.; Cole, A.; Hintelmann, H.; Loseto, L.L.; Steffen, A.; Wang, F. How does climate change influence arctic mercury? Sci. Total Environ. 2012, 414, 22–42. [Google Scholar] [CrossRef] [PubMed]
- Lacerda, L.D.d.; Marins, R.V.; Dias, F.J.d.S. An Arctic Paradox: Response of Fluvial Hg inputs and bioavailability to global climate change in an extreme coastal environment. Front. Earth Sci. 2020, 8, 93. [Google Scholar] [CrossRef]
- Hsu-Kim, H.; Kucharzyk, K.H.; Zhang, T.; Deshusses, M.A. Mechanisms regulating mercury bioavailability for methylating microorganisms in the aquatic environment: A critical review. Environ. Sci. Technol. 2013, 47, 2441–2456. [Google Scholar] [CrossRef] [PubMed]
- Abreu, S.; Soares, A.; Nogueira, A.; Morgado, F. Tree rings, Populus nigra L., as mercury data logger in aquatic environments: Case study of an historically contaminated environment. Bull. Environ. Contam. Toxicol. 2008, 80, 294–299. [Google Scholar] [CrossRef] [PubMed]
- Babst, F.; Bodesheim, P.; Charney, N.; Friend, A.D.; Girardin, M.P.; Klesse, S.; Moore, D.J.; Seftigen, K.; Björklund, J.; Bouriaud, O. When tree rings go global: Challenges and opportunities for retro-and prospective insight. Quat. Sci. Rev. 2018, 197, 1–20. [Google Scholar] [CrossRef]
- Babst, F.; Poulter, B.; Bodesheim, P.; Mahecha, M.D.; Frank, D.C. Improved tree-ring archives will support earth-system science. Nat. Ecol. Evol. 2017, 1, 1–2. [Google Scholar] [CrossRef]
- Brienen, R.J.; Schöngart, J.; Zuidema, P.A. Tree rings in the tropics: Insights into the ecology and climate sensitivity of tropical trees. Trop. Tree Physiol. 2016, 6, 439–461. [Google Scholar]
- Danek, M.; Bell, T.; Laroque, C.P. Some considerations in the reconstruction of lead levels using laser ablation: Lessons from the design stage of dendrochemistry study, St. John’s, Canada. Geochronometria 2015, 42, 217–231. [Google Scholar] [CrossRef]
- Jensen, N.; Holzmueller, E.; Edwards, P.; Thomas-Van Gundy, M.; DeWalle, D.; Williard, K. Tree response to experimental watershed acidification. Water Air Soil Pollut. 2014, 225, 1–12. [Google Scholar] [CrossRef]
- Jiskra, M.; Sonke, J.E.; Obrist, D.; Bieser, J.; Ebinghaus, R.; Myhre, C.L.; Pfaffhuber, K.A.; Wängberg, I.; Kyllönen, K.; Worthy, D. A vegetation control on seasonal variations in global atmospheric mercury concentrations. Nat. Geosci. 2018, 11, 244–250. [Google Scholar] [CrossRef]
- Żywiec, M.; Muter, E.; Zielonka, T.; Delibes, M.; Calvo, G.; Fedriani, J.M. Long-term effect of temperature and precipitation on radial growth in a threatened thermo-Mediterranean tree population. Trees 2017, 31, 491–501. [Google Scholar] [CrossRef]
- Babst, F.; Poulter, B.; Trouet, V.; Tan, K.; Neuwirth, B.; Wilson, R.; Carrer, M.; Grabner, M.; Tegel, W.; Levanic, T. Site-and species-specific responses of forest growth to climate across the E uropean continent. Glob. Ecol. Biogeogr. 2013, 22, 706–717. [Google Scholar] [CrossRef]
- Charney, N.D.; Babst, F.; Poulter, B.; Record, S.; Trouet, V.M.; Frank, D.; Enquist, B.J.; Evans, M.E. Observed forest sensitivity to climate implies large changes in 21st century North American forest growth. Ecol. Lett. 2016, 19, 1119–1128. [Google Scholar] [CrossRef] [PubMed]
- George, S.S.; Ault, T.R. The imprint of climate within Northern Hemisphere trees. Quat. Sci. Rev. 2014, 89, 1–4. [Google Scholar] [CrossRef]
- Martin-Benito, D.; Pederson, N. Convergence in drought stress, but a divergence of climatic drivers across a latitudinal gradient in a temperate broadleaf forest. J. Biogeogr. 2015, 42, 925–937. [Google Scholar] [CrossRef]
- Restaino, R.M.; Walsh, L.K.; Morishima, T.; Vranish, J.R.; Martinez-Lemus, L.A.; Fadel, P.J.; Padilla, J. Endothelial dysfunction following prolonged sitting is mediated by a reduction in shear stress. Am. J. Physiol. Heart Circ. Physiol. 2016, 310, H648–H653. [Google Scholar] [CrossRef] [PubMed]
- Tei, S.; Sugimoto, A.; Liang, M.; Yonenobu, H.; Matsuura, Y.; Osawa, A.; Sato, H.; Fujinuma, J.; Maximov, T. Radial growth and physiological response of coniferous trees to Arctic amplification. J. Geophys. Res. Biogeosci. 2017, 122, 2786–2803. [Google Scholar] [CrossRef]
- Perone, A.; Cocozza, C.; Cherubini, P.; Bachmann, O.; Guillong, M.; Lasserre, B.; Marchetti, M.; Tognetti, R. Oak tree-rings record spatial-temporal pollution trends from different sources in Terni (Central Italy). Environ. Pollut. 2018, 233, 278–289. [Google Scholar] [CrossRef] [PubMed]
- Scholes, R.J. Taking the mumbo out of the jumbo: Progress towards a robust basis for ecological scaling. Ecosystems 2017, 20, 4–13. [Google Scholar] [CrossRef]
- Yang, Y.; Yanai, R.D.; Montesdeoca, M.; Driscoll, C.T. Measuring mercury in wood: Challenging but important. Int. J. Environ. Anal. Chem. 2017, 97, 456–467. [Google Scholar] [CrossRef]
- Nillin, J. Avaliação da Qualidade do Sedimento do Estuário do rio Ceará. Dissertação de Mestrado (Engenharia de Pesca), Universidade Federal do Ceará, Fortaleza, Brazil, 2008. [Google Scholar]
- SEMACE. Área de Proteção Ambiental do Estuário do Rio Ceará. 2010. Available online: https://www.semace.ce.gov.br/2010/12/08/area-de-protecao-ambiental-do-estuario-do-rio-ceara/ (accessed on 1 May 2021).
- Davanso, M.B. Abordagens Biológicas Como Instrumento de Avaliação de Risco Ambiental em Estuários da Região Metropolitana de Fortaleza-CE: Biomarcadores e Toxicidade de Sedimentos. Dissertação Mestrado (Ciências Marinhas Tropicais), Instituto de Ciências do Mar, Fortaleza, Brazil, 2010; 74p. [Google Scholar]
- Nilin, J.; Castro, C.d.; Pimentel, M.; Franklin Júnior, W.; Matos, R.; Lotufo, T.; Costa-Lotufo, L. Water toxicity assessment of the Ceará river estuary (Brazil). J. Braz. Soc. Ecotoxicol. 2007, 2, 107–113. [Google Scholar] [CrossRef]
- Pech, D.; Vidal-Martínez, V.M.; Aguirre-Macedo, M.L.; Gold-Bouchot, G.; Herrera-Silveira, J.; Zapata-Pérez, O.; Marcogliese, D.J. The checkered puffer (Spheroides testudineus) and its helminths as bioindicators of chemical pollution in Yucatan coastal lagoons. Sci. Total Environ. 2009, 407, 2315–2324. [Google Scholar] [CrossRef] [PubMed]
- MacDonald, J.; Shahrestani, S.; Weis, J. Behavior and space utilization of two common fishes within Caribbean mangroves: Implications for the protective function of mangrove habitats. Estuar. Coast. Shelf Sci. 2009, 84, 195–201. [Google Scholar] [CrossRef]
- Costa, B.G.B. Distribuição de Mercurio (Hg) em Sedimento, Água e Biota na Região de Influencia dos Efluentes da Carcinicultura na Gamboa do Cumbe-Rio Jaguaribe-Ceará-Brasil. Dissertação de Mestrado (Ciências Marinhas Tropicais), Instituto de Ciências do Mar, Fortaleza, Brazil, 2009; 60p. [Google Scholar]
- Quintaneiro, C.; Querido, D.; Monteiro, M.; Guilhermino, L.; Morgado, F.; Soares, A.M.V.M. Transport and acclimation conditions for the use of an estuarine fish (Pomatoschistus microps) in ecotoxicity bioassays: Effects on enzymatic biomarkers. Chemosphere 2008, 71, 1803–1808. [Google Scholar] [CrossRef]
- Vieira, L.; Gravato, C.; Soares, A.; Morgado, F.; Guilhermino, L. Acute effects of copper and mercury on the estuarine fish Pomatoschistus microps: Linking biomarkers to behaviour. Chemosphere 2009, 76, 1416–1427. [Google Scholar] [CrossRef]
- Green-Ruiz, C.; Ruelas-Inzunza, J.; Páez-Osuna, F. Mercury in surface sediments and benthic organisms from Guaymas Bay, east coast of the Gulf of California. Environ. Geochem. Health 2005, 27, 321. [Google Scholar] [CrossRef]
- Mohsen, M.; Wang, Q.; Zhang, L.; Sun, L.; Lin, C.; Yang, H. Heavy metals in sediment, microplastic and sea cucumber Apostichopus japonicus from farms in China. Mar. Pollut. Bull. 2019, 143, 42–49. [Google Scholar] [CrossRef] [PubMed]
- Müller, G. Schadstoffe in sedimenten-sedimente als schadstoffe. Mitt Osterr. Geol. Ges. 1986, 79, 107–126. [Google Scholar]
- da Silva Dias, F.J.; Castro, B.M.; Lacerda, L.D. Tidal and low-frequency currents off the Jaguaribe River estuary (4° S, 37° 4′ W), northeastern Brazil. Ocean Dyn. 2018, 68, 967–985. [Google Scholar] [CrossRef]
- da Silva Dias, F.J.; Castro, B.M.; Lacerda, L.D.; Miranda, L.B.; Marins, R.V. Physical characteristics and discharges of suspended particulate matter at the continent-ocean interface in an estuary located in a semiarid region in northeastern Brazil. Estuar. Coast. Shelf Sci. 2016, 180, 258–274. [Google Scholar] [CrossRef]
- da Silva Dias, F.J.; Lacerda, L.D.; Marins, R.V.; de Paula, F.C.F. Comparative analysis of rating curve and ADP estimates of instantaneous water discharge through estuaries in two contrasting Brazilian rivers. Hydrol. Process. 2011, 25, 2188–2201. [Google Scholar] [CrossRef]
- Dias, F.; Castro, B.; Lacerda, L. Continental shelf water masses off the Jaguaribe River (4S), northeastern Brazil. Cont. Shelf Res. 2013, 66, 123–135. [Google Scholar] [CrossRef]
- Alvala, R.; Cunha, A.P.; Brito, S.S.; Seluchi, M.E.; Marengo, J.A.; Moraes, O.L.; Carvalho, M.A. Drought monitoring in the Brazilian Semiarid region. An. Acad. Bras. Ciências 2019, 91, e20170209. [Google Scholar] [CrossRef]
- Carvalho, A.; Marins, R.V.; Dias, F.; Rezende, C.E.; Lefèvre, N.; Cavalcante, M.; Eschrique, S. Air-sea CO2 fluxes for the Brazilian northeast continental shelf in a climatic transition region. J. Mar. Syst. 2017, 173, 70–80. [Google Scholar] [CrossRef]
- Marins, R.V.; Paula Filho, F.J.d.; Maia, S.R.R.; Lacerda, L.D.d.; Marques, W.S. Distribuição de mercúrio total como indicador de poluição urbana e industrial na costa brasileira. Química Nova 2004, 27, 763–770. [Google Scholar] [CrossRef]
- Maia, S.R.R. Distribuição e Partição Geoquímica de Metais Traço na Costa Norte de Fortaleza, CE. Dissertação (Mestrado em Ciências Marinhas Tropicais), Instituto de Ciências do Mar, Universidade Federal do Ceará, Fortaleza, Brazil, 2004; 105p. [Google Scholar]
- Mirlean, N.; Andrus, V.E.; Baisch, P. Mercury pollution sources in sediments of Patos Lagoon Estuary, Southern Brazil. Mar. Pollut. Bull. 2003, 46, 331–334. [Google Scholar] [CrossRef]
- Kehrig, H.A.; Pinto, F.N.; Moreira, I.; Malm, O. Heavy metals and methylmercury in a tropical coastal estuary and a mangrove in Brazil. Org. Geochem. 2003, 34, 661–669. [Google Scholar] [CrossRef]
- Garcia-Ordiales, E.; Covelli, S.; Rico, J.M.; Roqueñí, N.; Fontolan, G.; Flor-Blanco, G.; Cienfuegos, P.; Loredo, J. Occurrence and speciation of arsenic and mercury in estuarine sediments affected by mining activities (Asturias, northern Spain). Chemosphere 2018, 198, 281–289. [Google Scholar] [CrossRef] [PubMed]
- Cukrov, N.; Doumandji, N.; Garnier, C.; Tucaković, I.; Dang, D.H.; Omanović, D.; Cukrov, N. Anthropogenic mercury contamination in sediments of Krka River estuary (Croatia). Environ. Sci. Pollut. Res. 2020, 27, 7628–7638. [Google Scholar] [CrossRef]
- Vane, C.H.; Kim, A.W.; Emmings, J.F.; Turner, G.H.; Moss-Hayes, V.; Lort, J.A.; Williams, P.J. Grain size and organic carbon controls polyaromatic hydrocarbons (PAH), mercury (Hg) and toxicity of surface sediments in the River Conwy Estuary, Wales, UK. Mar. Pollut. Bull. 2020, 158, 111412. [Google Scholar] [CrossRef] [PubMed]
- Acquavita, A.; Covelli, S.; Emili, A.; Berto, D.; Faganeli, J.; Giani, M.; Horvat, M.; Koron, N.; Rampazzo, F. Mercury in the sediments of the Marano and Grado Lagoon (northern Adriatic Sea): Sources, distribution and speciation. Estuar. Coast. Shelf Sci. 2012, 113, 20–31. [Google Scholar] [CrossRef]
- Vane, C.H.; Beriro, D.J.; Turner, G.H. Rise and fall of mercury (Hg) pollution in sediment cores of the Thames Estuary, London, UK. Earth Environ. Sci. Trans. R. Soc. Edinb. 2015, 105, 285–296. [Google Scholar] [CrossRef]
- Pereira, E.; Abreu, S.N.; Coelho, J.P.; Lopes, C.B.; Pardal, M.A.; Vale, C.; Duarte, A.C. Seasonal fluctuations of tissue mercury contents in the European shore crab Carcinus maenas from low and high contamination areas (Ria de Aveiro, Portugal). Mar. Pollut. Bull. 2006, 52, 1450–1457. [Google Scholar] [CrossRef]
- Silva, L.; Machado, W.; Lisboa Filho, S.; Lacerda, L. Mercury accumulation in sediments of a mangrove ecosystem in SE Brazil. Water Air Soil Pollut. 2003, 145, 67–77. [Google Scholar] [CrossRef]
- Abreu, S.; Rodrigues, A.; Gadelha, J.; Morgado, F.; Soares, A. Mercury depuration in Pomatoschistus microps during acclimatation. Interdiscip. Stud. Environ. Chem. Biol. Responses Contam. Mol. Community Level 2010, 3, 159–164. [Google Scholar]
- Stoyanova, S.; Georgieva, E.; Velcheva, I.; Iliev, I.; Vasileva, T.; Bivolarski, V.; Tomov, S.; Nyeste, K.; Antal, L.; Yancheva, V. Multi-Biomarker Assessment in Common Carp (Cyprinus carpio, Linnaeus 1758) Liver after Acute Chlorpyrifos Exposure. Water 2020, 12, 1837. [Google Scholar] [CrossRef]
- Georgieva, E.; Yancheva, V.; Stoyanova, S.; Velcheva, I.; Iliev, I.; Vasileva, T.; Bivolarski, V.; Petkova, E.; László, B.; Nyeste, K.; et al. Which Is More Toxic? Evaluation of the Short-Term Toxic Effects of Chlorpyrifos and Cypermethrin on Selected Biomarkers in Common Carp (Cyprinus carpio, Linnaeus 1758). Toxics 2021, 9, 125. [Google Scholar] [CrossRef]
- Vieira, H.C.; Bordalo, M.D.; Rodrigues, A.C.M.; Pires, S.F.S.; Rocha, R.J.M.; Soares, A.M.V.M.; Rendón-von Osten, J.; Abreu, S.N.; Morgado, F. Water temperature modulates mercury accumulation and oxidative stress status of common goby (Pomatoschistus microps). Environ. Res. 2021, 193, 110585. [Google Scholar] [CrossRef] [PubMed]
- Nyeste, K.; Dobrocsi, P.; Czeglédi, I.; Czédli, H.; Harangi, S.; Baranyai, E.; Simon, E.; Nagy, S.A.; Antal, L. Age and diet-specific trace element accumulation patterns in different tissues of chub (Squalius cephalus): Juveniles are useful bioindicators of recent pollution. Ecol. Indic. 2019, 101, 1–10. [Google Scholar] [CrossRef]
- Dillon, M.E.; Wang, G.; Huey, R.B. Global metabolic impacts of recent climate warming. Nature 2010, 467, 704–706. [Google Scholar] [CrossRef]
- Ward, D.M.; Nislow, K.H.; Folt, C.L. Bioaccumulation syndrome: Identifying factors that make some stream food webs prone to elevated mercury bioaccumulation. Ann. N. Y. Acad. Sci. 2010, 1195, 62. [Google Scholar] [CrossRef]
- UN Environment. Global Mercury Assessment 2018. UN Environment Programme, Chemicals and Health Branch; UN Press: Geneva, Switzerland, 2019. [Google Scholar]
- IPCC. Climate Change 2007: The Physical Science Basis. Contribution of Working Group I to the Fourth Assessment Report of the Intergovernmental Panel on Climate Change; Solomon, S., Qin, D., Manning, M., Chen, Z., Marquis, M., Averyt, K.B., Tignor, M., Miller, H.L., Eds.; Cambridge University Press: Cambridge, UK; New York, NY, USA, 2007; p. 996. [Google Scholar]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Igeo | Class | Pollution Intensity |
|---|---|---|
| <0 | 1 | unpolluted |
| 0–1 | 2 | unpolluted to moderately polluted |
| 1–2 | 3 | moderately polluted |
| 2–3 | 4 | moderately to strongly polluted |
| 3–4 | 5 | strongly polluted |
| 4–5 | 6 | strongly to very strongly polluted |
| >5 | 7 | very strongly polluted |
| International Hg Environmental Guidelines | Hg Source | [Hg] (µg g−1) | Guidelines Standards |
|---|---|---|---|
| EQSBiota (1) | - | 0.02 | |
| Limit for human consumption (2) | - | 0.5 | EPA (3); FDA (4) |
| Limit of Mercury for Water for Protection of Aquatic Life | Inorganic Hg Methyl mercury | 0.026 0.004 | EPA (3) |
| Limit of Mercury for Fish (4) | 1 | FDA action level for methyl mercury | |
| Drinking water | - | 1 mg/L (total Hg) 2 mg/L (Inorganic Hg) | WHO (9); EPA |
| Baseline Values Standard for Soil and Sediments | Marine water | 0.13 | ISQG (7) |
| Baseline Values Standard for Soil and Sediments | Baseline | 0.4 | World Shale Value (µg/g) (6) |
| Baseline Values Standard for Soil and Sediments | Marine water | 0.70 | PEL (8) |
| Baseline Values Standard for Soil and Sediments (Soils) | Residential | 6.6 | Guideline value EPA, WHO (5) |
| Baseline Values Standard for Soil and Sediments (Soils) | Industrial | 50 | Guideline value EPA, WHO |
| Baseline Values Standard for Soil and Sediments (Soils) | Agricultural | 6.6 | Guideline value EPA, WHO |
| Pacoti River (Ceará, Brazil) | Fine sediments (<63 µm) | 0.02–0.02 | This study |
| Ceará River (Ceará, Brazil) | Fine sediments (<63 µm) | 0.01–0.07 | This study |
| Pacoti River (Ceará, Brazil) | Hg in muscles (S. testudineus) | 0.03–0.10 | This study |
| Ceará River (Ceará, Brazil) | Hg in muscles (S. testudineus) | 0.03–0.10 | This study |
| Pacoti River (Ceará, Brazil) | Hg in liver S. testudineus) | 0.08–0.18 | This study |
| Ceará River (Ceará, Brazil) | Hg in liver (S. testudineus) | 0.07–0.13 | This study |
| BASF—Pacoti River (Ceará, Brazil) | 1.35–4.04 | This study | |
| BASF—Ceará River (Ceará, Brazil) | 1.43–3.70 | This study |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Morgado, F.; Santos, R.M.A.L.; Sampaio, D.; de Lacerda, L.D.; Soares, A.M.V.M.; Vieira, H.C.; Abreu, S. Chronological Trends and Mercury Bioaccumulation in an Aquatic Semiarid Ecosystem under a Global Climate Change Scenario in the Northeastern Coast of Brazil. Animals 2021, 11, 2402. https://doi.org/10.3390/ani11082402
Morgado F, Santos RMAL, Sampaio D, de Lacerda LD, Soares AMVM, Vieira HC, Abreu S. Chronological Trends and Mercury Bioaccumulation in an Aquatic Semiarid Ecosystem under a Global Climate Change Scenario in the Northeastern Coast of Brazil. Animals. 2021; 11(8):2402. https://doi.org/10.3390/ani11082402
Chicago/Turabian StyleMorgado, Fernando, Ruy M. A. L. Santos, Daniela Sampaio, Luiz Drude de Lacerda, Amadeu M. V. M. Soares, Hugo C. Vieira, and Sizenando Abreu. 2021. "Chronological Trends and Mercury Bioaccumulation in an Aquatic Semiarid Ecosystem under a Global Climate Change Scenario in the Northeastern Coast of Brazil" Animals 11, no. 8: 2402. https://doi.org/10.3390/ani11082402
APA StyleMorgado, F., Santos, R. M. A. L., Sampaio, D., de Lacerda, L. D., Soares, A. M. V. M., Vieira, H. C., & Abreu, S. (2021). Chronological Trends and Mercury Bioaccumulation in an Aquatic Semiarid Ecosystem under a Global Climate Change Scenario in the Northeastern Coast of Brazil. Animals, 11(8), 2402. https://doi.org/10.3390/ani11082402