
Effects of Exogenous 6-Phytase (EC 3.1.3.26) Supplementation on Performance, Calcium and Phosphorous Digestibility, and Bone Mineralisation and Density in Weaned Piglets

 , , ,
, , ,
Abstract
Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Animals, Dietary Treatments, and Experimental Design
2.2. Experimental Procedures, Sampling, and Analyses
2.3. Computed Tomography Scanning of the Bones
2.4. Statistics
3. Results
3.1. Performance
3.2. Apparent Total Tract Digestibility of P and Ca
3.3. Bone Mineralization and Bone Density
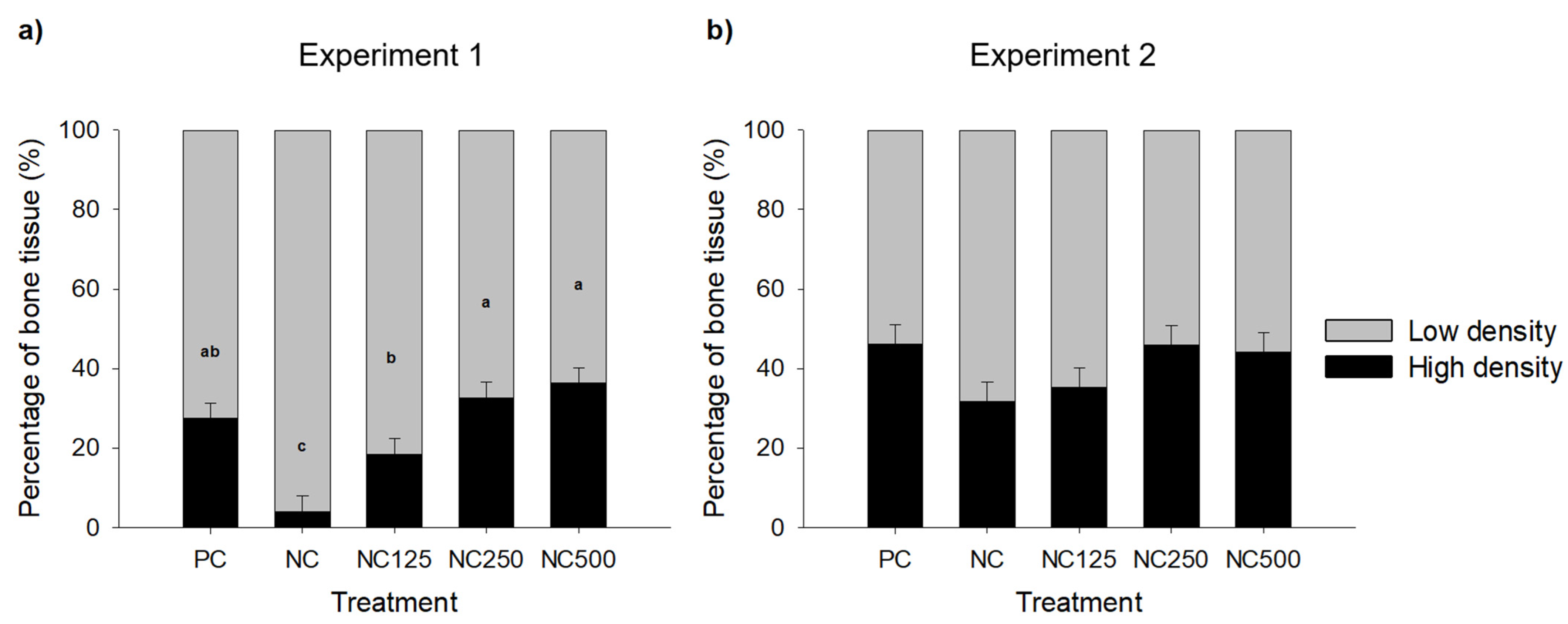
3.4. Image Analysis of Bones by Computed Tomography Scanning
3.5. Concentration of Ca and P and Alkaline Phosphatase Activity in Blood Plasma
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- O’Dell, B.L.; Sunde, R.A. Handbook of Nutritionally Essential Mineral Elements, 1st ed.; CRC Press: New York, NY, USA, 1997. [Google Scholar]
- Angel, R.; Tamim, N.; Applegate, T.; Dhandu, A.; Ellestad, L. Phytic acid chemistry: Influence on phytin-phosphorus availability and phytase efficacy. J. Appl. Poult. Res. 2002, 11, 471–480. [Google Scholar] [CrossRef]
- Ravindran, V.; Bryden, W.; Kornegay, E. Phytates: Occurrence, bioavailability and implications in poultry nutrition. Poult. Avian. Biol. Rev. 1995, 6, 125–143. [Google Scholar]
- Jongbloed, A.W.; Mroz, Z.; Kemme, P.A. The effect of supplementary Aspergillus niger phytase in diets for pigs on concentration and apparent digestibility of dry matter, total phosphorus, and phytic acid in different sections of the alimentary tract. J. Anim. Sci. 1992, 70, 1159–1168. [Google Scholar] [CrossRef]
- Pointillart, A. Enhancement of phosphorus utilization in growing pigs fed phytate-rich diets by using rye bran. J. Anim. Sci. 1991, 69, 1109–1115. [Google Scholar] [CrossRef] [PubMed]
- Selle, P.; Ravindran, V.; Caldwell, A.; Bryden, W. Phytate and phytase: Consequences for protein utilisation. Nutr. Res. Rev. 2000, 13, 255–278. [Google Scholar] [CrossRef] [PubMed]
- Torrallardona, D.; Tarradas, J.; Tous, N. The use of exogenous enzymes to optimise gut function in pigs. In Understanding Gut Microbiomes as Targets for Improving Gut Health; Bailey, M., Stokes, C., Eds.; Burleigh Dodds Science Publishing Limited: Sawston, Cambridge, UK, 2021; Volume 1, p. 50. [Google Scholar]
- Cheryan, M.; Rackis, J.J. Phytic acid interactions in food systems. Crit. Rev. Food Sci. Nutr. 1980, 13, 297–335. [Google Scholar] [CrossRef] [PubMed]
- Cowieson, A.; Ravindran, V.; Selle, P. Influence of dietary phytic acid and source of microbial phytase on ileal endogenous amino acid flows in broiler chickens. Poult. Sci. J. 2008, 87, 2287–2299. [Google Scholar] [CrossRef]
- Reddy, N.; Sathe, S.; Salunkhe, D. Phytates in legumes and cereals. Adv. Food Nutr. Res. 1982, 28, 1–92. [Google Scholar]
- Woyengo, T.; Nyachoti, C. Anti-nutritional effects of phytic acid in diets for pigs and poultry-current knowledge and directions for future research. Can. J. Anim. Sci. 2013, 93, 9–21. [Google Scholar] [CrossRef]
- Woyengo, T.A.; Cowieson, A.J.; Adeola, O.; Nyachoti, C.M. Ileal digestibility and endogenous flow of minerals and amino acids: Responses to dietary phytic acid in piglets. Br. J. Nutr. 2009, 102, 428–433. [Google Scholar] [CrossRef] [PubMed]
- Deshpande, S.; Cheryan, M. Effects of Phytic Acid, Divalent Cations, and Their Interactions on α-Amylase Activity. J. Food Sci. 1984, 49, 516–519. [Google Scholar] [CrossRef]
- Singh, M.; Krikorian, A. Inhibition of trypsin activity in vitro by phytate. J. Agric. Food Chem. 1982, 30, 799–800. [Google Scholar] [CrossRef]
- Dersjant-Li, Y.; Awati, A.; Schulze, H.; Partridge, G. Phytase in non-ruminant animal nutrition: A critical review on phytase activities in the gastrointestinal tract and influencing factors. J. Sci. Food Agric. 2015, 95, 878–896. [Google Scholar] [CrossRef] [PubMed]
- Kornegay, E.; Harper, A.; Jones, R.; Boyd, L. Environmental nutrition: Nutrient management strategies to reduce nutrient excretion of swine. Prof. Anim. Sci. 1997, 13, 99–111. [Google Scholar] [CrossRef]
- Correll, D.L. The role of phosphorus in the eutrophication of receiving waters: A review. J. Environ. Qual. 1998, 27, 261–266. [Google Scholar] [CrossRef]
- Smith, V.H.; Tilman, G.D.; Nekola, J.C. Eutrophication: Impacts of excess nutrient inputs on freshwater, marine, and terrestrial ecosystems. Environ. Pollut. 1999, 100, 179–196. [Google Scholar] [CrossRef]
- Selle, P.H.; Ravindran, V. Phytate-degrading enzymes in pig nutrition. Livest. Sci. 2008, 113, 99–122. [Google Scholar] [CrossRef]
- Schlemmer, U.; Jany, K.D.; Berk, A.; Schulz, E.; Rechkemmer, G. Degradation of phytate in the gut of pigs-Pathway of gastrointestinal inositol phosphate hydrolysis and enzymes involved. Arch. Anim. Nutr. Arch. Fur Tierernahr. 2001, 55, 255–280. [Google Scholar] [CrossRef]
- Wyss, M.; Brugger, R.; Kronenberger, A.; Rémy, R.; Fimbel, R.; Oesterhelt, G.; Lehmann, M.; van Loon, A.P. Biochemical characterization of fungal phytases (myo-inositol hexakisphosphate phosphohydrolases): Catalytic properties. Appl. Environ. Microbiol. 1999, 65, 367–373. [Google Scholar] [CrossRef]
- Yu, S.; Cowieson, A.; Gilbert, C.; Plumstead, P.; Dalsgaard, S. Interactions of phytate and myo-inositol phosphate esters (IP) including IP isomers with dietary protein and iron and inhibition of pepsin. Anim. Sci. J. 2012, 90, 1824–1832. [Google Scholar] [CrossRef]
- Lei, X.G.; Weaver, J.D.; Mullaney, E.; Ullah, A.H.; Azain, M.J. Phytase, a new life for an “old” enzyme. Annu. Rev. Anim. Biosci. 2013, 1, 283–309. [Google Scholar] [CrossRef]
- Zeller, E.; Schollenberger, M.; Kühn, I.; Rodehutscord, M. Hydrolysis of phytate and formation of inositol phosphate isomers without or with supplemented phytases in different segments of the digestive tract of broilers. J. Nutr. Sci 2015, 4, e1. [Google Scholar] [CrossRef] [PubMed]
- Torrallardona, D.; Ader, P. Effects of a novel 6-phytase (EC 3.1. 3.26) on performance, phosphorus and calcium digestibility, and bone mineralization in weaned piglets. Anim. Sci. J. 2016, 94, 194–197. [Google Scholar] [CrossRef]
- Skiba, G.; Sobol, M.; Raj, S. Femur morphometry, densitometry, geometry and mechanical properties in young pigs fed a diet free of inorganic phosphorus and supplemented with phytase. Arch. Anim. Nutr. 2017, 71, 81–92. [Google Scholar] [CrossRef]
- Nielsen, D.H.; McEvoy, F.J.; Madsen, M.T.; Jensen, J.B.; Svalastoga, E. Relationship between bone strength and dual-energy X-ray absorptiometry measurements in pigs. Anim. Sci. J. 2007, 85, 667–672. [Google Scholar] [CrossRef][Green Version]
- Doroudinia, A.; Colletti, P.M. Bone mineral measurements. Clin. Nucl. Med. 2015, 40, 647–657. [Google Scholar] [CrossRef]
- Ibanez, R. Bone mineral density measurement techniques. Anales Sist. Sanitario Navar. 2003, 26, 19–27. [Google Scholar]
- du Chazaud, B.B.; Monziols, M.; Gaudre, D.; Quemeneur, B. La tomographie rayons X chez le porcelet comme prédicteur de l’état de minéralisation du squelette. In Proceedings of the Journées Recherche Porcine, Paris, France, 3–4 February 2015; pp. 121–122. [Google Scholar]
- Fabà, L.; Gasa, J.; Tokach, M.D.; Font-i-Furnols, M.; Vilarrasa, E.; Solà-Oriol, D. Effects of additional organic micro-minerals and methionine on carcass composition, gait score, bone characteristics, and osteochondrosis in replacement gilts of different growth rate. Anim. Feed Sci. Technol. 2019, 256, 114262. [Google Scholar] [CrossRef]
- Gaudré, D.; Lebas, N.; Monziols, M. Effets d’une séquence déplétion-réplétion en phosphore en engraissement. In Proceedings of the Journées Recherche Porcine, Paris, France, 4–5 February 2014; pp. 119–120. [Google Scholar]
- Engelen, A.J.; Van Der Heeft, F.C.; Randsdorp, P.H.; Smtt, E.L. Simple and rapid determination of phytase activity. J. AOAC Int. 1994, 77, 760–764. [Google Scholar] [CrossRef]
- NRC. Nutrient Requirements of Swine, 11th ed.; The National Academies of Press: Washington, DC, USA, 2012; p. 210. [Google Scholar]
- De Blas, C.; Gasa, J.; Mateos, G. Necesidades Nutricionales Para Ganado Porcino. Normas FEDNA, 2nd ed.; Fundación Española para el Desarrollo de la Nutrición Animal: Madrid, Spain, 2013; p. 36. [Google Scholar]
- Short, F.; Gorton, P.; Wiseman, J.; Boorman, K. Determination of titanium dioxide added as an inert marker in chicken digestibility studies. Anim. Feed Sci. Technol. 1996, 59, 215–221. [Google Scholar] [CrossRef]
- Adeola, O. Digestion and balance techniques in pigs. In Swine Nutrition, 2nd ed.; Lewis, A., Southern, L., Eds.; CRC Press: New York, NY, USA, 2001; pp. 903–916. [Google Scholar]
- Bardera, A.; Martίnez, R.; Boada, I.; Font-i-Furnols, M.; Gispert, M. VisualPork towards the simulation of a virtual butcher. In Proceedings of the First Annual Conference on Body and Carcass Evaluation, Meat Quality, Sofware and Traceability (FAIM I). Conference of COST Action FA1102, Ashtown, Dublin, Ireland, 24–26 September 2012; pp. 25–26. [Google Scholar]
- Boada, I.; Spinola, J.; Rodriguez, J.; Martínez, R.; Font i Furnols, M. VisualPork towards the simulation of a virtual butcher. In Proceedings of the II Workshop on the use of computed tomography (CT) in pig carcass classification. Other CT applications: Live animals and meat technology, Monells, Spain, 16–17 April 2009; pp. 16–17. [Google Scholar]
- Font-i-Furnols, M.; Carabús, A.; Pomar, C.; Gispert, M. Estimation of carcass composition and cut composition from computed tomography images of live growing pigs of different genotypes. Animal 2015, 9, 166–178. [Google Scholar] [CrossRef] [PubMed]
- Olsen, E.V.; Christensen, L.B.; Nielsen, D.B. A review of computed tomography and manual dissection for calibration of devices for pig carcass classification-Evaluation of uncertainty. Meat Sci. 2017, 123, 35–44. [Google Scholar] [CrossRef]
- Xiberta, P.; Bardera, A.; Boada, I.; Gispert, M.; Brun, A.; Font-i-Furnols, M. Evaluation of an automatic lean meat percentage quantification method based on a partial volume model from computed tomography scans. Comput. Electron. Agric. 2018, 151, 365–375. [Google Scholar] [CrossRef]
- Picouet, P.A.; Teran, F.; Gispert, M.; i Furnols, M.F. Lean content prediction in pig carcasses, loin and ham by computed tomography (CT) using a density model. Meat Sci. 2010, 86, 616–622. [Google Scholar] [CrossRef]
- Grubbs, F.E. Procedures for Detecting Outlying Observations in Samples. Technometrics 1969, 11, 1–21. [Google Scholar] [CrossRef]
- Humer, E.; Schwarz, C.; Schedle, K. Phytate in pig and poultry nutrition. J. Anim. Physiol. Anim. Nutr. 2015, 99, 605–625. [Google Scholar] [CrossRef]
- Pallauf, J.; Rimbach, G. Nutritional significance of phytic acid and phytase. Arch. Fur Tierernahr. 1997, 50, 301–319. [Google Scholar] [CrossRef] [PubMed]
- Selle, P.H.; Cowieson, A.J.; Cowieson, N.P.; Ravindran, V. Protein-phytate interactions in pig and poultry nutrition: A reappraisal. Nutr. Res. Rev. 2012, 25, 1–17. [Google Scholar] [CrossRef] [PubMed]
- Bento, M.H.; Pedersen, C.; Plumstead, P.W.; Salmon, L.; Nyachoti, C.M.; Bikker, P. Dose response of a new phytase on dry matter, calcium, and phosphorus digestibility in weaned piglets. Anim. Sci. J. 2012, 90, 245–247. [Google Scholar] [CrossRef]
- Jolliff, J.S.; Mahan, D.C. Effect of dietary inulin and phytase on mineral digestibility and tissue retention in weanling and growing swine. Anim. Sci. J. 2012, 90, 3012–3022. [Google Scholar] [CrossRef]
- Blavi, L.; Munoz, C.J.; Broomhead, J.N.; Stein, H.H. Effects of a novel corn-expressed E. coli phytase on digestibility of calcium and phosphorous, growth performance, and bone ash in young growing pigs. Anim. Sci. J. 2019, 97, 3390–3398. [Google Scholar] [CrossRef] [PubMed]
- Rosenfelder-Kuon, P.; Siegert, W.; Rodehutscord, M. Effect of microbial phytase supplementation on P digestibility in pigs: A meta-analysis. Arch. Anim. Nutr. 2020, 74, 1–18. [Google Scholar] [CrossRef] [PubMed]
- Campbell, J.M.; Crenshaw, J.D.; Polo, J. The biological stress of early weaned piglets. J. Anim. Sci. Biotechnol. 2013, 4, 19. [Google Scholar] [CrossRef] [PubMed]
- Manners, M.J. The development of digestive function in the pig. P Nutr. Soc. 1976, 35, 49–55. [Google Scholar] [CrossRef]
- Snoeck, V.; Cox, E.; Verdonck, F.; Joensuu, J.J.; Goddeeris, B.M. Influence of porcine intestinal pH and gastric digestion on antigenicity of F4 fimbriae for oral immunisation. Vet. Microbiol. 2004, 98, 45–53. [Google Scholar] [CrossRef]
- Torres-Pitarch, A.; Hermans, D.; Manzanilla, E.G.; Bindelle, J.; Everaert, N.; Beckers, Y.; Torrallardona, D.; Bruggeman, G.; Gardiner, G.E.; Lawlor, P.G. Effect of feed enzymes on digestibility and growth in weaned pigs: A systematic review and meta-analysis. Anim. Feed Sci. Tech. 2017, 233, 145–159. [Google Scholar] [CrossRef]
- Sung, J.Y.; Kim, B.G. Prediction models for apparent and standardized total tract digestible phosphorus in swine diets. Anim. Feed Sci. Tech. 2019, 255, 114224. [Google Scholar] [CrossRef]
- Calvert, C.C.; Besecker, R.J.; Plumlee, M.P.; Cline, T.R.; Forsyth, D.M. Apparent Digestibility of Phosphorus in Barley and Corn for Growing Swine. Anim. Sci. J. 1978, 47, 420–426. [Google Scholar] [CrossRef]
- Huang, K.; Allee, G. Bioavailability of Phosphorus in Selected Feedstuffs for Pigs. Kans. Agric. Exp. Stn. Res. Rep. 1980, 28–29. [Google Scholar] [CrossRef]
- Boyd, R.D.; Hall, D.; Wu, J.F. Plasma alkaline phosphatase as a criterion for determining biological availability of phosphorus for swine. J. Anim. Sci. 1983, 57, 396–401. [Google Scholar] [CrossRef][Green Version]
- Eeckhout, W.; De Paepe, M.; Warnants, N.; Bekaert, H. An estimation of the minimal P requirements for growing-finishing pigs, as influenced by the Ca level of the diet. Anim. Feed Sci. Tech. 1995, 52, 29–40. [Google Scholar] [CrossRef]
- Aasmundstad, T.; Kongsro, J.; Wetten, M.; Dolvik, N.; Vangen, O. Osteochondrosis in pigs diagnosed with computed tomography: Heritabilities and genetic correlations to weight gain in specific age intervals. Animal 2013, 7, 1576–1582. [Google Scholar] [CrossRef]
- Guggenbuhl, P.; Calvo, E.P.; Fru, F. Effects of dietary doses of three phytases on performance in pigs. J. Anim. Sci. 2016, 94, 286–288. [Google Scholar] [CrossRef]
- Huff, W.E.; Moore, P.A., Jr.; Waldroup, P.W.; Waldroup, A.L.; Balog, J.M.; Huff, G.R.; Rath, N.C.; Daniel, T.C.; Raboy, V. Effect of dietary phytase and high available phosphorus corn on broiler chicken performance. Poult. Sci. J. 1998, 77, 1899–1904. [Google Scholar] [CrossRef] [PubMed]
- Saikia, A.; Sarma, D.N.; Bhuyan, R.; Sarmah, B.C.; Kalita, D. Effect of zinc and phytase supplementation on performance, serum biochemical profiles and carcass quality of crossbred (Hampshire × Assam local) pigs. J. Appl. Anim. 2016, 44, 230–233. [Google Scholar] [CrossRef][Green Version]
- Lagos, L.V.; Walk, C.L.; Murphy, M.R.; Stein, H.H. Effects of dietary digestible calcium on growth performance and bone ash concentration in 50- to 85-kg growing pigs fed diets with different concentrations of digestible phosphorus. Anim. Feed. Sci. Tech. 2019, 247, 262–272. [Google Scholar] [CrossRef]
- Lee, S.A.; Lagos, L.V.; Bedford, M.R.; Stein, H.H. Quantities of ash, Ca, and P in metacarpals, metatarsals, and tibia are better correlated with total body bone ash in growing pigs than ash, Ca, and P in other bones. J. Anim. Sci. 2021, 99, 1–6. [Google Scholar] [CrossRef]
- Omogbenigun, F.O.; Nyachoti, C.M.; Slominski, B.A. The effect of supplementing microbial phytase and organic acids to a corn-soybean based diet fed to early-weaned pigs. J. Anim. Sci. 2003, 81, 1806–1813. [Google Scholar] [CrossRef] [PubMed]
- Zeng, Z.K.; Piao, X.S.; Wang, D.; Li, P.F.; Xue, L.F.; Salmon, L.; Zhang, H.Y.; Han, X.; Liu, L. Effect of Microbial Phytase on Performance, Nutrient Absorption and Excretion in Weaned Pigs and Apparent Ileal Nutrient Digestibility in Growing Pigs. Asian Australas. J. Anim. Sci. 2011, 24, 1164–1172. [Google Scholar] [CrossRef]
- Guggenbuhl, P.; Calvo, E.P.; Fru, F. Effect of a bacterial 6-phytase on plasma myo-inositol concentrations and P and Ca utilization in swine. J. Anim. Sci. 2016, 94, 243–245. [Google Scholar] [CrossRef]
- Guggenbuhl, P.; Calvo, E.P.; Fru, F. Effect of high dietary doses of a bacterial 6-phytase in piglets fed a corn–soybean meal diet. J. Anim. Sci. 2016, 94, 307–309. [Google Scholar] [CrossRef]
- Skiba, G.; Weremko, D.; Sobol, M.; Raj, S. Bone mineralisation of weaned piglets fed a diet free of inorganic phosphorus and supplemented with phytase, as assessed by dual-energy X-ray absorptiometry. Arch. Anim. Nutr. 2015, 69, 267–275. [Google Scholar] [CrossRef] [PubMed]
- Zeng, Z.K.; Wang, D.; Piao, X.S.; Li, P.F.; Zhang, H.Y.; Shi, C.X.; Yu, S.K. Effects of Adding Super Dose Phytase to the Phosphorus-deficient Diets of Young Pigs on Growth Performance, Bone Quality, Minerals and Amino Acids Digestibilities. Asian Australas. J. Anim. Sci. 2014, 27, 237–246. [Google Scholar] [CrossRef] [PubMed]



{kind=link}
{kind=link}
| 1–21 d (Pre-Starter) | 21–42 d (Starter) | |||||||
|---|---|---|---|---|---|---|---|---|
| Experiment 1 | Experiment 2 | Experiment 1 | Experiment 2 | |||||
| PC | NC | PC | NC | PC | NC | PC | NC | |
| Ingredients (%) | ||||||||
| Maize | 48.4 | 50.5 | 48.1 | 50.1 | 65.9 | 67.5 | 64.9 | 66.6 |
| Soybean meal (48% CP) | 24.2 | 24.2 | 24.4 | 24.4 | 27.8 | 27.8 | 28.5 | 28.5 |
| Extruded soybeans | 10.0 | 10.0 | 10.0 | 10.0 | - | - | - | - |
| Sweet milk whey | 11.0 | 11.0 | 11.0 | 11.0 | - | - | - | - |
| Lard | 2.69 | 1.86 | 2.75 | 1.91 | 2.83 | 2.14 | 2.89 | 2.20 |
| L-Lysine-HCl | 0.32 | 0.31 | 0.31 | 0.31 | 0.38 | 0.37 | 0.38 | 0.37 |
| L-Threonine | 0.13 | 0.13 | 0.13 | 0.12 | 0.15 | 0.14 | 0.15 | 0.15 |
| DL-Methionine | 0.18 | 0.17 | 0.18 | 0.17 | 0.15 | 0.15 | 0.15 | 0.14 |
| L-Tryptophan | 0.03 | 0.03 | 0.03 | 0.03 | 0.03 | 0.03 | 0.03 | 0.03 |
| Salt | 0.16 | 0.16 | 0.16 | 0.16 | 0.36 | 0.36 | 0.36 | 0.36 |
| Calcium carbonate | 0.38 | 0.55 | 0.21 | 0.38 | 0.45 | 0.59 | 0.28 | 0.42 |
| Dicalcium phosphate 2 | 1.39 | 0.05 | 1.63 | 0.29 | 1.41 | 0.30 | 1.65 | 0.54 |
| Ethoxyquin 3 | 0.02 | 0.02 | 0.02 | 0.02 | 0.02 | 0.02 | 0.02 | 0.02 |
| Vit-Min complex 4 | 0.60 | 0.60 | 0.60 | 0.60 | 0.60 | 0.60 | 0.60 | 0.60 |
| TiO2 | 0.50 | 0.50 | 0.50 | 0.50 | - | - | - | - |
| Analysed nutrient content (%) | ||||||||
| Dry matter | 89.7 | 89.3 | 88.1 | 88.0 | 89.0 | 88.4 | 87.4 | 87.3 |
| Crude Protein | 21.0 | 20.1 | 20.5 | 20.9 | 19.0 | 19.0 | 19.4 | 19.1 |
| Fat | 6.06 | 5.31 | 7.77 | 6.40 | 5.58 | 4.91 | 5.31 | 4.32 |
| Ash | 6.14 | 5.02 | 5.68 | 4.86 | 4.92 | 4.11 | 4.66 | 3.97 |
| Gross Energy (MJ/kg) | 17.4 | 17.3 | 17.0 | 16.9 | 17.3 | 17.1 | 16.9 | 16.7 |
| Calcium | 0.83 | 0.55 | 0.81 | 0.47 | 0.78 | 0.60 | 0.67 | 0.57 |
| Phosphorous | 0.57 | 0.31 | 0.66 | 0.40 | 0.56 | 0.38 | 0.63 | 0.44 |
| PC | NC | NC125 | NC250 | NC500 | RMSE | p-Value | L2 | Q3 | |
|---|---|---|---|---|---|---|---|---|---|
| Experiment 1 | |||||||||
| Body weight (kg) | |||||||||
| 0 d | 7.33 | 7.37 | 7.30 | 7.33 | 7.29 | 0.090 | 0.414 | 0.145 | 0.737 |
| 21 d | 13.0 | 12.4 | 13.4 | 12.7 | 13.3 | 1.022 | 0.302 | 0.265 | 0.623 |
| 42 d | 24.1 a,b | 21.2 c | 23.8 a,b | 23.1 b | 25.0 a | 1.609 | <0.001 | <0.001 | 0.368 |
| Weight gain (g/d) | |||||||||
| 0–21 d | 270 | 241 | 293 | 257 | 285 | 48 | 0.227 | 0.210 | 0.597 |
| 21–42 d | 527 a,b | 418 c | 494 b | 493 b | 559 a | 40 | <0.001 | <0.001 | 0.261 |
| 0–42 d | 398 a,b | 329 c | 393 a,b | 375 b | 422 a | 38 | <0.001 | <0.001 | 0.360 |
| Feed intake (g/d) | |||||||||
| 0–21 d | 387 | 383 | 393 | 372 | 414 | 59 | 0.696 | 0.356 | 0.430 |
| 21–42 d | 854 a,b | 733 c | 804 b | 835 b | 903 a | 61 | <0.001 | <0.001 | 0.361 |
| 0–42 d | 621 a,b | 558 c | 598 b,c | 603 a,b,c | 658 a | 55 | 0.019 | 0.001 | 0.937 |
| Gain/feed | |||||||||
| 0–21 d | 0.698 | 0.627 | 0.743 | 0.693 | 0.691 | 0.073 | 0.060 | 0.332 | 0.047 |
| 21–42 d | 0.618 a | 0.573 b | 0.616 a | 0.595 a,b | 0.619 a | 0.034 | 0.039 | 0.036 | 0.438 |
| 0–42 d | 0.643 a | 0.592 b | 0.658 a | 0.625 a,b | 0.642 a | 0.035 | 0.010 | 0.063 | 0.089 |
| Experiment 2 | |||||||||
| Body weight (kg) | |||||||||
| 0 d | 9.20 | 9.20 | 9.18 | 9.18 | 9.17 | 0.031 | 0.293 | 0.337 | 0.356 |
| 21 d | 15.3 | 15.0 | 15.2 | 15.3 | 14.8 | 0.817 | 0.722 | 0.561 | 0.274 |
| 42 d | 29.1 | 27.1 | 28.0 | 28.8 | 28.3 | 1.482 | 0.093 | 0.111 | 0.089 |
| Weight gain (g/d) | |||||||||
| 0–21 d | 292 | 277 | 289 | 291 | 270 | 39 | 0.729 | 0.582 | 0.261 |
| 21–42 d | 655 a | 575 b | 608 a,b | 645 a | 643 a | 57 | 0.048 | 0.021 | 0.165 |
| 0–42 d | 473 | 426 | 449 | 468 | 456 | 35 | 0.085 | 0.111 | 0.085 |
| Feed intake (g/d) | |||||||||
| 0–21 d | 416 | 397 | 397 | 395 | 389 | 46 | 0.814 | 0.729 | 0.889 |
| 21–42 d | 1017 | 938 | 963 | 1024 | 977 | 76 | 0.156 | 0.262 | 0.090 |
| 0–42 d | 717 | 668 | 680 | 710 | 683 | 50 | 0.256 | 0.484 | 0.169 |
| Gain/feed | |||||||||
| 0–21 d | 0.718 | 0.717 | 0.740 | 0.749 | 0.718 | 0.070 | 0.824 | 0.932 | 0.265 |
| 21–42 d | 0.646 | 0.613 | 0.633 | 0.629 | 0.658 | 0.030 | 0.054 | 0.007 | 0.939 |
| 0–42 d | 0.665 | 0.640 | 0.663 | 0.660 | 0.671 | 0.028 | 0.273 | 0.063 | 0.434 |
| PC | NC | NC125 | NC250 | NC500 | RMSE | p-Value | L2 | Q3 | |
|---|---|---|---|---|---|---|---|---|---|
| Experiment 1 | |||||||||
| P ATTD | 41.2 a | 9.80 d | 21.0 c | 33.3 b | 42.0 a | 6.77 | <0.001 | <0.001 | 0.023 |
| Ca ATTD | 54.2 b,c | 51.3 c | 57.9 b | 69.3 a | 68.6 a | 4.26 | <0.001 | <0.001 | <0.001 |
| Experiment 2 | |||||||||
| P ATTD | 52.0 a | 29.1 d | 28.0 d | 40.0 c | 46.8 b | 4.36 | <0.001 | <0.001 | 0.763 |
| Ca ATTD | 48.6 a | 32.6 b | 37.5 b | 48.9 a | 45.0 a | 6.37 | <0.001 | <0.001 | 0.004 |
| PC | NC | NC125 | NC250 | NC500 | RMSE | p-Value | L2 | Q3 | |
|---|---|---|---|---|---|---|---|---|---|
| Experiment 1 | |||||||||
| Dry weight (g) | 2.65 a,b | 1.85 c | 2.37 b | 2.46 a,b | 2.81 a | 0.364 | <0.001 | <0.001 | 0.185 |
| Ash (%) | 42.6 a | 30.1 d | 33.0 c,d | 36.0 b,c | 36.6 b | 3.34 | <0.001 | <0.001 | 0.083 |
| Ash (g) | 1.12 a | 0.56 d | 0.78 c | 0.88 b,c | 1.03 a,b | 0.145 | <0.001 | <0.001 | 0.095 |
| Density (g/cm3) | 1.56 a,b | 1.41 c | 1.49 b,c | 1.59 a | 1.62 a | 0.075 | <0.001 | <0.001 | 0.047 |
| Experiment 2 | |||||||||
| Dry weight (g) | 3.24 a | 2.31 c | 2.64 b,c | 2.76 b | 2.98 a,b | 0.337 | <0.001 | <0.001 | 0.310 |
| Ash (%) | 43.3 a | 36.7 c | 37.6 c | 39.8 b,c | 40.9 a,b | 3.19 | 0.002 | 0.008 | 0.602 |
| Ash (g) | 1.41 a | 0.85 d | 0.99 c | 1.09 b,c | 1.22 b | 0.135 | <0.001 | <0.001 | 0.246 |
| Density (g/cm3) | 1.70 | 1.62 | 1.68 | 1.72 | 1.71 | 0.135 | 0.595 | 0.217 | 0.306 |
| PC | NC | NC500 | RMSE | p-Value | |
|---|---|---|---|---|---|
| Experiment 1 | |||||
| Calcium (mmol/L) | 2.45 | 2.31 | 2.46 | 0.158 | 0.145 |
| Phosphate (mmol/L) | 2.79 a | 1.79 b | 2.55 a | 0.347 | <0.001 |
| Ca:P ratio | 0.89 b | 1.33 a | 0.98 b | 0.190 | 0.001 |
| Alkaline Phosphatase (I.U./L) | 414 b | 1001 a | 506 b | 227.5 | <0.001 |
| Experiment 2 | |||||
| Calcium (mmol/L) | 2.82 | 2.83 | 2.88 | 0.194 | 0.807 |
| Phosphate (mmol/L) | 2.95 a | 2.19 b | 3.18 a | 0.321 | <0.001 |
| Ca:P ratio | 0.96 b | 1.34 a | 0.92 b | 0.176 | <0.001 |
| Alkaline Phosphatase (I.U./L) | 173 b | 225 a | 168 b | 36.5 | 0.013 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Tous, N.; Tarradas, J.; Francesch, M.; Font-i-Furnols, M.; Ader, P.; Torrallardona, D. Effects of Exogenous 6-Phytase (EC 3.1.3.26) Supplementation on Performance, Calcium and Phosphorous Digestibility, and Bone Mineralisation and Density in Weaned Piglets. Animals 2021, 11, 1787. https://doi.org/10.3390/ani11061787
Tous N, Tarradas J, Francesch M, Font-i-Furnols M, Ader P, Torrallardona D. Effects of Exogenous 6-Phytase (EC 3.1.3.26) Supplementation on Performance, Calcium and Phosphorous Digestibility, and Bone Mineralisation and Density in Weaned Piglets. Animals. 2021; 11(6):1787. https://doi.org/10.3390/ani11061787
Chicago/Turabian StyleTous, Núria, Joan Tarradas, Maria Francesch, Maria Font-i-Furnols, Peter Ader, and David Torrallardona. 2021. "Effects of Exogenous 6-Phytase (EC 3.1.3.26) Supplementation on Performance, Calcium and Phosphorous Digestibility, and Bone Mineralisation and Density in Weaned Piglets" Animals 11, no. 6: 1787. https://doi.org/10.3390/ani11061787
APA StyleTous, N., Tarradas, J., Francesch, M., Font-i-Furnols, M., Ader, P., & Torrallardona, D. (2021). Effects of Exogenous 6-Phytase (EC 3.1.3.26) Supplementation on Performance, Calcium and Phosphorous Digestibility, and Bone Mineralisation and Density in Weaned Piglets. Animals, 11(6), 1787. https://doi.org/10.3390/ani11061787