1. Introduction
The gastrointestinal (GI) tract digests and absorbs nutrients and acts as a barrier against toxic compounds and pathogens from the diets taken. Moreover, intestinal epithelial cells are sealed with paracellular tight junctions to regulate the permeability of the intestinal barrier, which is associated with nutrient absorption and immune activity [
1]. The epithelial barrier in newborn pigs is rapidly formed with decreased intestinal permeability and optimized liquid-form nutrient absorption during the lactation period. After weaning, piglets undergo several challenges, such as the separation from their mothers, mixing with other piglets, transferring to new environments, switching from digestible liquid diets to solid diets, and immature digestive and immune systems [
2,
3]. Generally, the weaning stress induces the intestinal dysfunction of weaned pigs, leading to reduced growth performance and increased intestinal problems and diseases that are the major concerns in swine production [
4].
It is known that glutamic acid (GLU) is a non-essential amino acid that is abundantly present in the body and plays an essential function in cellular metabolism and immune responses [
5]. The GLU is a precursor for protein synthesis and has been attracting attention for its diverse metabolic functions in weaned pigs [
6,
7]. Moreover, GLU is an important precursor for intestinal formation and immune system in the small intestine and can contribute to the immediate effect on nutrient absorption [
7]. Previous studies reported that GLU improved the performance of weaned pigs [
7,
8,
9]. The potential mechanisms of its action are as follows: (1) stimulation of oral and visceral sensory fibers [
10], (2) an energy source to form the intestinal barrier structure and functions [
11], and (3) regulation of intestinal nervous and immune systems as a signaling compound [
12]. However, the connection between dietary GLU and gut physiological benefits under weaning stress is not fully understood. Therefore, the objective of this study was to investigate the effects of dietary GLU on the growth performance, nutrient digestibility, immune responses, and intestinal health of weaned pigs.
4. Discussion
The post-weaning period is critical for the intestinal development of weaned pigs, as weaning stress can lead to intestinal barrier dysfunction, immune system destruction, and intestinal microbiota disruption, thereby reducing the absorption of nutrients and causing intestinal diseases such as diarrhea [
4]. Therefore, appropriate nutrient supply for weaned pigs is one of essential factors to improve the intestinal health. Several studies reported that some feed additives, such as plant extracts, exogenous enzymes, and GLU, had beneficial effects on growth performance and gut barrier functions of pigs [
8,
14,
15,
28]. In particular, GLU is regarded as one of the most promising performance-enhancing additives with amino acid synthesis properties [
8]. However, the correlations of dietary GLU on growth performance and intestinal barrier and immune functions of pigs are not fully understood. In the present study, we demonstrated that dietary GLU efficiently influenced growth rate and gut health of weaned pigs.
The present study showed dietary GLU enhanced the ADG and ADFI of weaned pigs for the first two weeks after weaning. These observations are in agreement with previous studies, suggesting that dietary GLU may give more beneficial effects on growth performance of weaned pigs around the first 10 to 21 days after weaning [
9,
29,
30]. These results indicate that dietary GLU promotes the growth performance of pigs by increasing feed intake and performing as a metabolic fuel that more effectively increases cell turnover rate during intestinal development [
8,
9]. Moreover, it is well known that GLU has umami properties that may have an important role in the taste, palatability, and acceptability of the feeds [
6,
31]. The GLU not only stimulates taste and oral sensation by binding with umami receptors in the oral cavity, but is also involved in signaling in the brain for the autonomic reflex of the digestive responses associated with salivation for chewing and swallowing [
11]. In the present study, there was no dietary GLU effect on the frequency of diarrhea during the first two weeks after weaning that may be caused by the weaning events. However, a previous study reported that supplementation with 4% dietary GLU in weaner diets alleviated diarrhea in the
Escherichia coli infected pigs [
32]. This is because the metabolites that produced by GLU, such as N-acetyl glucosamine and N-acetylgalactosamine residues, bind to same receptors for
Escherichia coli in intestinal epithelial cells to relieve diarrhea [
33]. These findings suggested that more nutrient supply is required for the growth of weaned pigs in early intestinal development and dietary GLU may be involved in amino acid metabolisms in the initial phase [
32,
33].
A previous study reported that several stress factors, such as maternal separation, mixing/crowding, dietary change, and GI infection after weaning, could cause intestinal dysfunction which inhibits the absorption of nutrients in the small intestine [
34]. During the weaning period, the intestinal structure and function of pigs are immature by declining the reduction in villus enzyme activity, VH, and transepithelial electrical resistance [
35]. In the results from the present study, dietary GLU enhanced the AID and ATTD of DM and energy. These results are in agreement with previous studies, suggesting that GLU improves the intestinal digestive process by acting as a signaling compound in the intestinal nervous system and regulating the neuroendocrine functions of the GI tract [
11,
36]. The GLU-related receptors, such as T1R1, T1R3, mGluR4 and mGluR1, are distributed in the stomach and intestine, regulating the mucosal responses in the stomach and duodenum [
36]. Moreover, GLU is involved in maintaining the intestinal barrier functions and structure of weaned pigs. The present study also observed that dietary GLU improved the intestinal morphology by increasing VH:CD and number of goblet cells. These observations are in agreement with several in vivo and in vitro studies, reporting that GLU regulates the proliferation and differentiation of intestinal epithelial cells [
36,
37,
38]. The GLU serves as an important energy source for cell division in porcine mucosa and inhibits the activities of apoptotic proteins, such as caspase families, phospho-p53, and cytochrome, which induce intestinal cell apoptosis in weaned pigs [
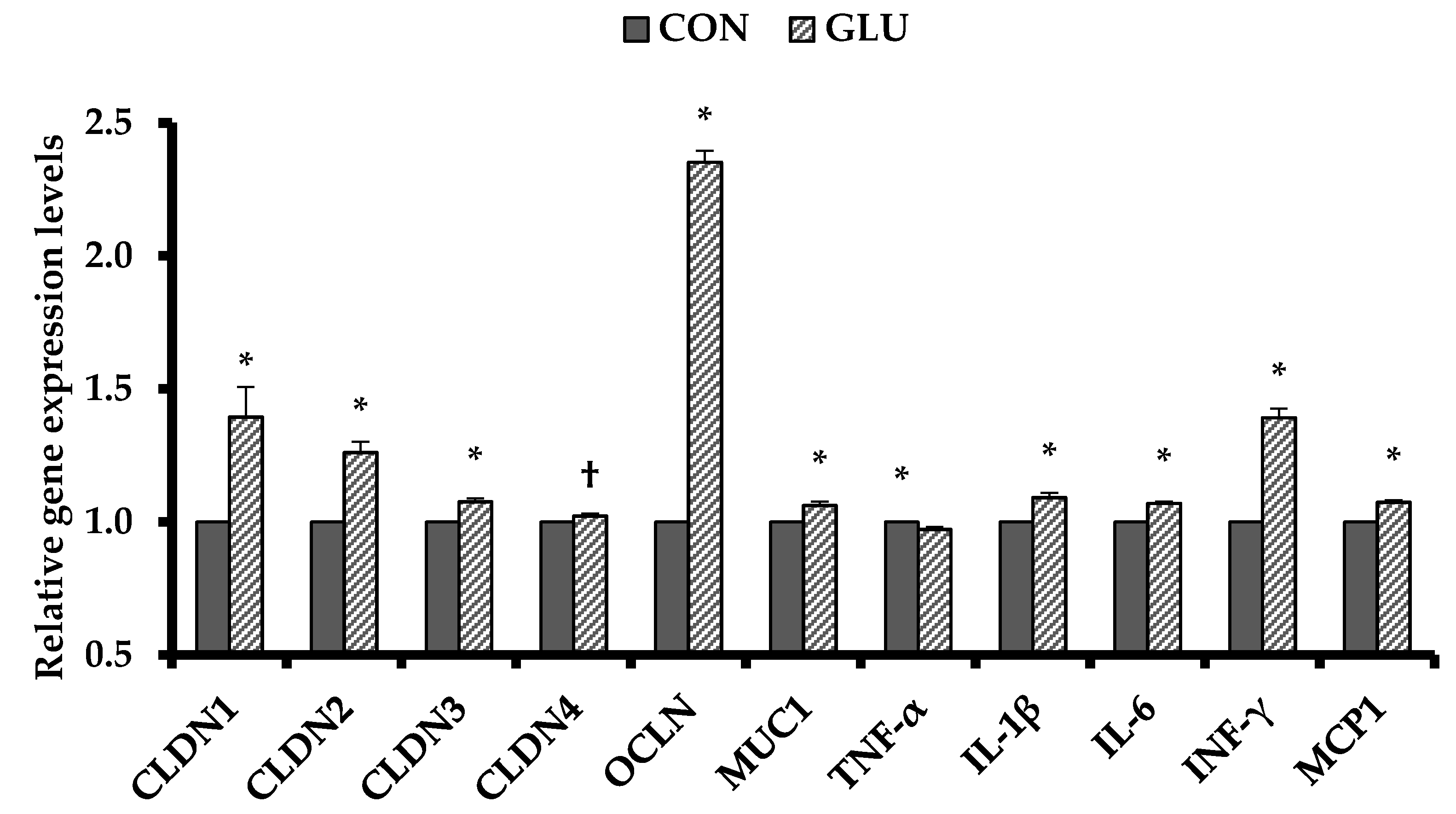
7]. Furthermore, the intestinal barrier functions in the intestinal epithelium play a critical role in nutrient absorption by pump action via the enzyme activity of the electrolyte transporters distributed on the brush border. In addition, the tight junction between the epithelial cells regulates gate function through intercellular membrane proteins, such as ZO-1, occludin, and claudins [
34]. Similarly, the present study verified that dietary GLU improved the intestinal barrier functions by improving gene expressions of tight junction proteins in the ileal tissues of weaned pigs. These results are in agreement with previous studies on nutrient digestibility and gut barrier functions [
8,
39,
40]. These physiological benefits suggest that dietary GLU improved nutrient digestibility directly by strengthening the intestinal barrier functions.
Weaning stress overreacts the hypothalamic pituitary adrenal axis and corticotropin releasing factor system to release mast cell proteases and TNF-α in the intestine, which increase intestinal epithelium permeability and induce the penetration of external antigens [
34]. Inflammatory cytokines play the most important part in the regulation of immune system and are closely related to weaning stress [
4]. In the present study, dietary GLU decreased TNF-α, IL-1β, and IL-6 levels in the serum and downregulated TNF-α gene expression levels in the ileal tissue, suggesting that dietary GLU may have anti-inflammatory effects. Previous studies indicated that GLU stimulated the synthesis of intestinal mucosa cells and might be used as major fuel and substrates for metabolism and immune responses [
5,
7,
41]. Like other immune response-related amino acids, GLU also plays a role in regulating the immune system. Previous studies showed that dietary GLU alleviated the incidence of diarrhea and had anti-oxidative stress effects in weaned pigs [
42,
43,
44]. The results from the present study are consistent with these previous observations. Therefore, the immune responses by dietary GLU in weaner diets supported that dietary GLU might contribute to improvement of the intestinal health and development during the weaning stress.
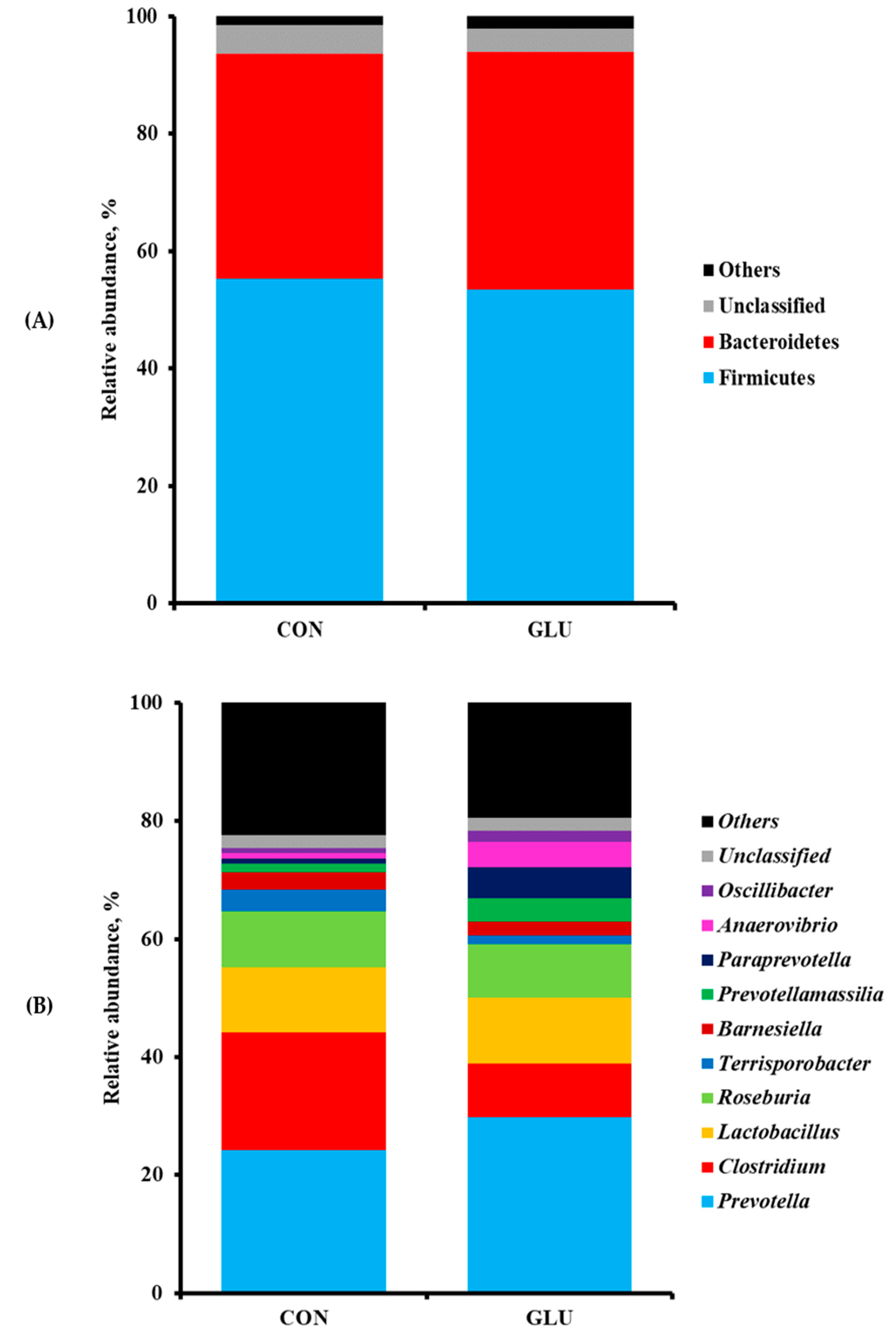
Weaning is a major challenge for piglets to adapt to the GI microbial colonization and physiological changes in their intestine, following the transition of diets from milk to feed as well as other changes. Therefore, the gut microbial analysis was performed as a pilot study using small numbers of animals to assess whether dietary GLU changed microbial population of weaned pigs. Our results showed that the phyla Firmicutes and Bacteroidetes comprised over 90% of all bacteria in the microbiomes of pigs in both CON and GLU. Previous studies reported that both phyla Firmicutes and Bacteroidetes contributed to the production of volatile fatty acids, such as acetate, propionate, and butyrate, through the digestible fiber in plant-derived weaning diets compared with the pre-weaning milk diets [
45,
46]. In addition, higher proportions of bacteria from both phyla Firmicutes and Bacteroidetes were associated with an increase in BW and these phyla improved the energy production capacity of pigs during the digestion process [
47].
The present study also showed pigs fed GLU had more
Prevotella and
Anaerovibrio than those fed CON. The previous study mentioned that
Prevotella was more common in the gut microbiota of pigs after weaning than in that of breast-fed piglets because they could ferment the indigestible polysaccharides into short-chain fatty acids in the gut [
17]. In addition,
Anaerovibrio strains, such as
A. lipolytica, are linked to fat metabolism with the production of lipase to hydrolyze triglycerides [
48]. In the present study, the relative abundances of genera
Clostridium and
Terrisporobacter were also lower in the gut microbiota of weaned pigs fed GLU than in that of weaned pigs fed CON. Generally, spore-forming
Clostridium strains are recognized as the primary cause of diarrhea in neonatal and weaned piglets [
49]. Previous studies reported that the spores and toxins from
C. difficile and
C. perfringens originated during the lactation period induced high rates of diarrhea and mortality after weaning [
49,
50,
51]. Moreover,
Terrisporobacter produces the urinary toxin, such as trimethylamine-
N-oxide, which is associated with oxidative stress and inflammation in the gut of weaned pigs [
52]. Therefore, it is plausible that the increased abundances of
Prevotella and
Anaerovibrio in the gut microbiota of weaned pigs fed GLU may contribute to the reduced abundances of
Clostridium and
Terrisporobacter compared with those of weaned pigs fed CON, thereby improving the intestinal health of weaned pigs by stabilizing the immune state and intestinal environment.


 ,
, 



{kind=link}
{kind=link}
{kind=link}