
The Impact of Genetics on Gut Microbiota of Growing and Fattening Pigs under Moderate N Restriction



Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Animals, Diets and Sampling Procedure
2.2. Genomic DNA Extraction, 16S rRNA Amplicon Sequencing and Bioinformatics
2.3. Statistical Analysis
3. Results
3.1. Dataset Features
3.2. Alpha Diversity
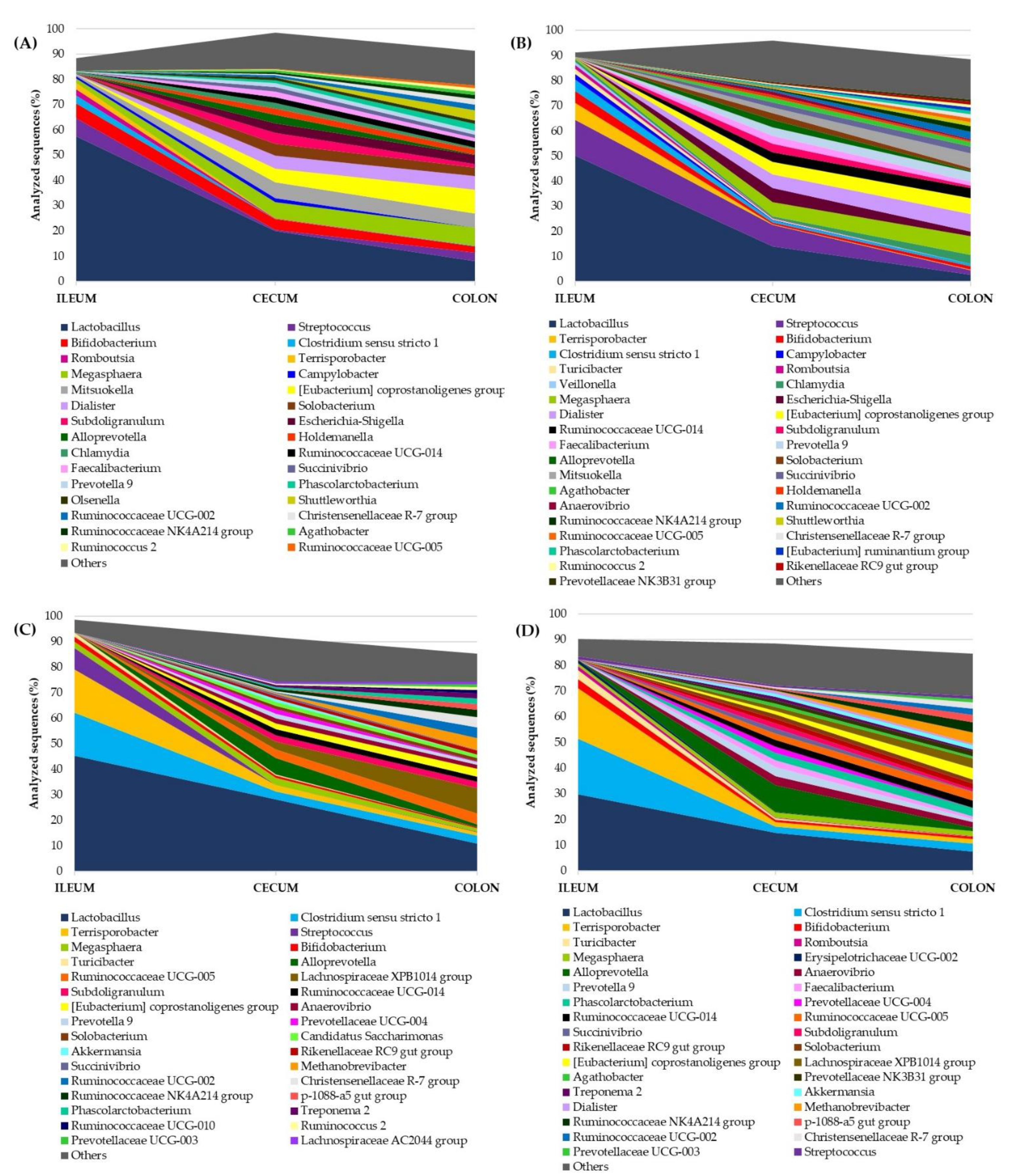
3.3. Microbial Composition throughout the Intestinal Tract
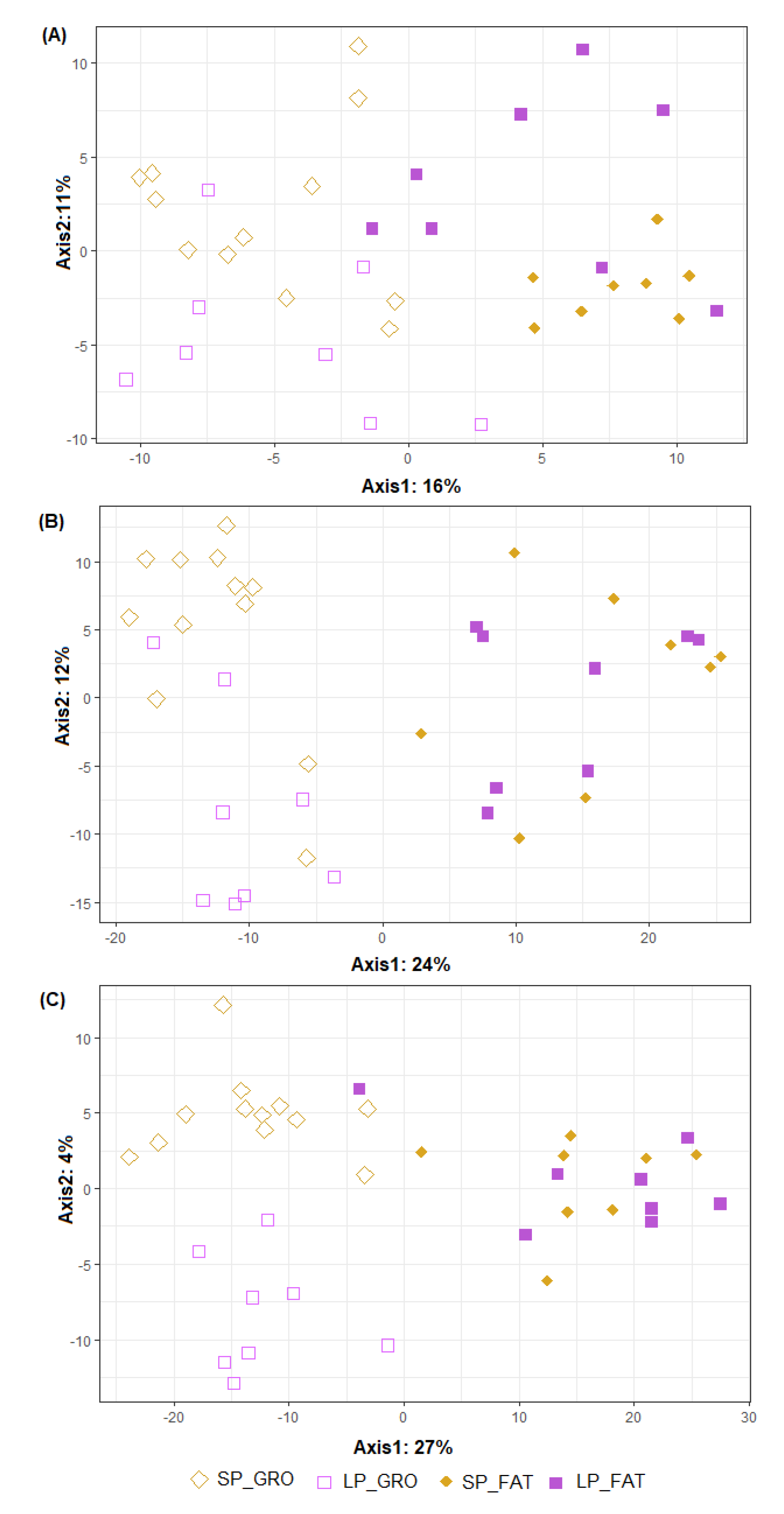
3.4. Beta Diversity
3.5. Microbial Networks
4. Discussion
4.1. Effect of Production Phase
4.2. Effect of Producing Type of Pig
4.3. Effect of Dietary CP Content
4.4. Microbiota Composition throughout the Intestinal Tract
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Pomar, C.; Remus, A. Precision pig feeding: A breakthrough toward sustainability. Anim. Front. 2019, 9, 52–59. [Google Scholar] [CrossRef]
- Gardiner, G.E.; Metzler-Zebeli, B.U.; Lawlor, P.G. Impact of intestinal microbiota on growth and feed efficiency in pigs: A review. Microorganisms 2020, 8, 1886. [Google Scholar] [CrossRef]
- Wang, Y.; Zhou, J.; Wang, G.; Cai, S.; Zeng, X.; Qiao, S. Advances in low-protein diets for swine. J. Anim. Sci. Biotechnol. 2018, 9. [Google Scholar] [CrossRef]
- Directive (EU) 1991/676 of the European Parliament and of the Council. Available online: https://eur-lex.europa.eu/legal-content/EN/TXT/?uri=CELEX%3A31991L0676&qid=1623746555297 (accessed on 15 June 2021).
- Díaz-Caro, C.; García-Torres, S.; Elghannam, A.; Tejerina, D.; Mesias, F.J.; Ortiz, A. Is production system a relevant attribute in consumers’ food preferences? The case of Iberian dry-cured ham in Spain. Meat Sci. 2019, 158. [Google Scholar] [CrossRef]
- Yang, H.; Xiang, Y.; Robinson, K.; Wang, J.; Zhang, G.; Zhao, J.; Xiao, Y. Gut microbiota is a major contributor to adiposity in pigs. Front. Microbiol. 2018, 9. [Google Scholar] [CrossRef]
- Niederwerder, M.C.; Jaing, C.J.; Thissen, J.B.; Cino-Ozuna, A.G.; McLoughlin, K.S.; Rowland, R.R.R. Microbiome associations in pigs with the best and worst clinical outcomes following co-infection with porcine reproductive and respiratory syndrome virus (PRRSV) and porcine circovirus type 2 (PCV2). Vet. Microbiol. 2016, 188. [Google Scholar] [CrossRef] [Green Version]
- Quan, J.; Cai, G.; Yang, M.; Zeng, Z.; Ding, R.; Wang, X.; Zhuang, Z.; Zhou, S.; Li, S.; Yang, H.; et al. Exploring the fecal microbial composition and metagenomic functional capacities associated with feed efficiency in commercial DLY pigs. Front. Microbiol. 2019, 10. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wu, C.; Lyu, W.; Hong, Q.; Zhang, X.; Yang, H.; Xiao, Y. Gut microbiota influence lipid metabolism of skeletal muscle in pigs. Front. Nutr. 2021, 8. [Google Scholar] [CrossRef] [PubMed]
- Guo, X.; Xia, X.; Tang, R.; Zhou, J.; Zhao, H.; Wang, K. Development of a real-time PCR method for Firmicutes and Bacteroidetes in faeces and its application to quantify intestinal population of obese and lean pigs. Lett. Appl. Microbiol. 2008, 47, 367–373. [Google Scholar] [CrossRef] [PubMed]
- Richards, J.D.; Gong, J.; de Lange, C.F.M. The gastrointestinal microbiota and its role in monogastric nutrition and health with an emphasis on pigs: Current understanding, possible modulations, and new technologies for ecological studies. Can. J. Anim. Sci. 2005, 85, 421–435. [Google Scholar] [CrossRef]
- Zhao, W.; Wang, Y.; Liu, S.; Huang, J.; Zhai, Z.; He, C.; Ding, J.; Wang, J.; Wang, H.; Fan, W.; et al. The dynamic distribution of porcine microbiota across different ages and gastrointestinal tract segments. PLoS ONE 2015, 10, 117441. [Google Scholar] [CrossRef] [Green Version]
- Trevisi, P.; Luise, D.; Correa, F.; Bosi, P. Timely control of gastrointestinal eubiosis: A strategic pillar of pig health. Microorganisms 2021, 9, 313. [Google Scholar] [CrossRef]
- de Blas, C.; Gasa, J.; Mateos, G.G. Necesidades Nutricionales Para Ganado Porcino. Normas FEDNA, 2nd ed.; Normas FEDNA: Madrid, Spain, 2013. [Google Scholar]
- Sarri, L.; Balcells, J.; de la Fuente, G.; Tor, M.; Gómez-Arrue, J.; Seradj, A.R. Evolution of viscera and muscle fractional protein synthesis rate in lean meat selected hybrids and castrated Duroc pigs fed under moderate crude protein restriction. Animal 2021, 15. [Google Scholar] [CrossRef]
- Callahan, B.J.; McMurdie, P.J.; Rosen, M.J.; Han, A.W.; Johnson, A.J.A.; Holmes, S.P. DADA2: High-resolution sample inference from Illumina amplicon data. Nat. Methods 2016, 13, 581–583. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, Q.; Garrity, G.M.; Tiedje, J.M.; Cole, J.R. Naïve Bayesian classifier for rapid assignment of rRNA sequences into the new bacterial taxonomy. Appl. Environ. Microbiol. 2007, 73, 5261–5267. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Calle, M.L. Statistical analysis of metagenomics data. Genom. Inform. 2019, 17. [Google Scholar] [CrossRef]
- Gloor, G.B.; Macklaim, J.M.; Pawlowsky-Glahn, V.; Egozcue, J.J. Microbiome datasets are compositional: And this is not optional. Front. Microbiol. 2017, 8. [Google Scholar] [CrossRef] [Green Version]
- Fernandes, A.D.; Macklaim, J.M.; Linn, T.G.; Reid, G.; Gloor, G.B. ANOVA-like differential expression (ALDEx) analysis for mixed population RNA-Seq. PLoS ONE 2013, 8, 067019. [Google Scholar] [CrossRef]
- Friedman, J.; Alm, E.J. Inferring correlation networks from genomic survey data. PLoS Comput. Biol. 2012, 8. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ke, S.; Fang, S.; He, M.; Huang, X.; Yang, H.; Yang, B.; Chen, C.; Huang, L. Age-based dynamic changes of phylogenetic composition and interaction networks of health pig gut microbiome feeding in a uniformed condition. BMC Vet. Res. 2019, 15. [Google Scholar] [CrossRef] [Green Version]
- Wang, X.; Tsai, T.; Deng, F.; Wei, X.; Chai, J.; Knapp, J.; Apple, J.; Maxwell, C.V.; Lee, J.A.; Li, Y.; et al. Longitudinal investigation of the swine gut microbiome from birth to market reveals stage and growth performance associated bacteria. Microbiome 2019, 7. [Google Scholar] [CrossRef] [Green Version]
- Han, G.G.; Lee, J.-Y.; Jin, G.-D.; Park, J.; Choi, Y.H.; Kang, S.-K.; Chae, B.J.; Kim, E.B.; Choi, Y.-J. Tracing of the fecal microbiota of commercial pigs at five growth stages from birth to shipment. Sci. Rep. 2018, 8. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- De Rodas, B.; Youmans, B.P.; Danzeisen, J.L.; Tran, H.; Johnson, T.J. Microbiome profiling of commercial pigs from farrow to finish. J. Anim. Sci. 2018, 96, 1778–1794. [Google Scholar] [CrossRef]
- Crespo-Piazuelo, D.; Estellé, J.; Revilla, M.; Criado-Mesas, L.; Ramayo-Caldas, Y.; Óvilo, C.; Fernández, A.I.; Ballester, M.; Folch, J.M. Characterization of bacterial microbiota compositions along the intestinal tract in pigs and their interactions and functions. Sci. Rep. 2018, 8. [Google Scholar] [CrossRef] [PubMed]
- Hutchens, L.K.; Hintz, R.L.; Johnson, R.K. Breed comparisons for age and weight at puberty in gilts. J. Anim. Sci. 1982, 55, 60–66. [Google Scholar] [CrossRef]
- Len, N.T.; Hong, T.T.T.; Ogle, B.; Lindberg, J.E. Comparison of total tract digestibility, development of visceral organs and digestive tract of Mong Cai and Yorkshire x Landrace piglets fed diets with different fibre sources. J. Anim. Physiol. Anim. Nutr. (Berl.) 2009, 93, 181–191. [Google Scholar] [CrossRef]
- Freire, J.P.B.; Peiniau, J.; Cunha, L.F.; Almeida, J.A.A.; Aumaitre, A. Comparative effects of dietary fat and fibre in Alentejano and Large White piglets: Digestibility, digestive enzymes and metabolic data. Livest. Prod. Sci. 1998, 53, 37–47. [Google Scholar] [CrossRef]
- Kostic, A.D.; Howitt, M.R.; Garrett, W.S. Exploring host-microbiota interactions in animal models and humans. Genes Dev. 2013, 27, 701–718. [Google Scholar] [CrossRef] [Green Version]
- Wood, J.D.; Nute, G.R.; Richardson, R.I.; Whittington, F.M.; Southwood, O.; Plastow, G.; Mansbridge, R.; da Costa, N.; Chang, K.C. Effects of breed, diet and muscle on fat deposition and eating quality in pigs. Meat Sci. 2004, 67, 651–667. [Google Scholar] [CrossRef]
- Pajarillo, E.A.B.; Chae, J.P.; Balolong, M.P.; Kim, H.B.; Seo, K.-S.; Kang, D.-K. Pyrosequencing-based analysis of fecal microbial communities in three purebred pig lines. J. Microbiol. 2014, 52, 646–651. [Google Scholar] [CrossRef]
- Bergamaschi, M.; Tiezzi, F.; Howard, J.; Huang, Y.J.; Gray, K.A.; Schillebeeckx, C.; McNulty, N.P.; Maltecca, C. Gut microbiome composition differences among breeds impact feed efficiency in swine. Microbiome 2020, 8. [Google Scholar] [CrossRef]
- Seradj, A.R.; Balcells, J.; Sarri, L.; Fraile, L.J.; de la fuente, G. The impact of producing type and dietary crude protein on animal performances and microbiota together with greenhouse gases emissions in growing pigs. Animals 2020, 10, 1742. [Google Scholar] [CrossRef]
- Xiao, Y.; Kong, F.; Xiang, Y.; Zhou, W.; Wang, J.; Yang, H.; Zhang, G.; Zhao, J. Comparative biogeography of the gut microbiome between Jinhua and Landrace pigs. Sci. Rep. 2018, 8. [Google Scholar] [CrossRef]
- Xiao, Y.; Li, K.; Xiang, Y.; Zhou, W.; Gui, G.; Yang, H. The fecal microbiota composition of boar Duroc, Yorkshire, Landrace and Hampshire pigs. Asian-Australas. J. Anim. Sci. 2017, 30, 1456–1463. [Google Scholar] [CrossRef] [Green Version]
- Ju, Y.C.; Antonopoulos, D.A.; Kalra, A.; Tonelli, A.; Khalife, W.T.; Schmidt, T.M.; Young, V.B. Decreased diversity of the fecal microbiome in recurrent Clostridium difficile-associated diarrhea. J. Infect. Dis. 2008, 197, 435–438. [Google Scholar] [CrossRef] [Green Version]
- Dunne, J.A.; Williams, R.J.; Martinez, N.D. Network structure and biodiversity loss in food webs: Robustness increases with connectance. Ecol. Lett. 2002, 5, 558–567. [Google Scholar] [CrossRef] [Green Version]
- Costa-Roura, S. Alternatives for the Redesign of Beef Cattle Production: Dietary Protein, Forage Intake and Feed Efficiency. Ph.D. Thesis, Universitat de Lleida, Lleida, Spain, 2021. [Google Scholar]
- Wang, X.; Zhang, Y.; Wen, Q.; Wang, Y.; Wang, Z.; Tan, Z.; Wu, K. Sex differences in intestinal microbial composition and function of Hainan special wild boar. Animals 2020, 10, 1553. [Google Scholar] [CrossRef] [PubMed]
- Harada, N.; Hanaoka, R.; Horiuchi, H.; Kitakaze, T.; Mitani, T.; Inui, H.; Yamaji, R. Castration influences intestinal microflora and induces abdominal obesity in high-fat diet-fed mice. Sci. Rep. 2016, 6. [Google Scholar] [CrossRef] [PubMed]
- Kovacs, A.; Ben-Jacob, N.; Tayem, H.; Halperin, E.; Iraqi, F.A.; Gophna, U. Genotype is a stronger determinant than sex of the mouse gut microbiota. Microb. Ecol. 2011, 61, 423–428. [Google Scholar] [CrossRef] [PubMed]
- Guo, X.; Xia, X.; Tang, R.; Wang, K. Real-time PCR quantification of the predominant bacterial divisions in the distal gut of Meishan and Landrace pigs. Anaerobe 2008, 14, 224–228. [Google Scholar] [CrossRef]
- Park, S.-J.; Kim, J.; Lee, J.-S.; Rhee, S.-K.; Kim, H. Characterization of the fecal microbiome in different swine groups by high-throughput sequencing. Anaerobe 2014, 28, 157–162. [Google Scholar] [CrossRef]
- Gloaguen, M.; Floc’h, N.L.; Corrent, E.; Primot, Y.; van Milgen, J. The use of free amino acids allows formulating very low crude protein diets for piglets. Am. Soc. Anim. Sci. 2014, 92, 637–644. [Google Scholar] [CrossRef]
- Qiu, K.; Zhang, X.; Jiao, N.; Xu, D.; Huang, C.; Wang, Y.; Yin, J. Dietary protein level affects nutrient digestibility and ileal microbiota structure in growing pigs. Anim. Sci. J. 2017, 89, 537–546. [Google Scholar] [CrossRef] [PubMed]
- Yan, H.; Potu, R.; Lu, H.; Vezzoni de Almeida, V.; Stewart, T.; Ragland, D.; Armstrong, A.; Adeola, O.; Nakatsu, C.H.; Ajuwon, K.M. Dietary fat content and fiber type modulate hind gut microbial community and metabolic markers in the pig. PLoS ONE 2013, 8, 059581. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Costa-Roura, S.; Balcells, J.; de la Fuente, G.; Mora-Gil, J.; Llanes, N.; Villalba, D. Effects of protein restriction on performance, ruminal fermentation and microbial community in Holstein bulls fed high-concentrate diets. Anim. Feed Sci. Technol. 2020, 264, 114479. [Google Scholar] [CrossRef]
- Ghosh, T.S.; Gupta, S.S.; Bhattacharya, T.; Yadav, D.; Barik, A.; Chowdhury, A.; Das, B.; Mande, S.S.; Nair, G.B. Gut microbiomes of Indian children of varying nutritional status. PLoS ONE 2014, 9, 095547. [Google Scholar] [CrossRef]
- Zhou, L.; Fang, L.; Sun, Y.; Su, Y.; Zhu, W. Effects of the dietary protein level on the microbial composition and metabolomic profile in the hindgut of the pig. Anaerobe 2016, 38, 61–69. [Google Scholar] [CrossRef]
- Gresse, R.; Chaucheyras-Durand, F.; Dunière, L.; Blanquet-Diot, S.; Forano, E. Microbiota composition and functional profiling throughout the gastrointestinal tract of commercial weaning piglets. Microorganisms 2019, 7, 343. [Google Scholar] [CrossRef] [Green Version]
- Wang, J.; Han, Y.; Zhao, J.Z.; Zhou, Z.J.; Fan, H. Pyrosequencing-based analysis of the complex microbiota located in the gastrointestinal tracts of growing-finishing pigs. Anim. Prod. Sci. 2019, 59, 870–878. [Google Scholar] [CrossRef]
- Niu, Q.; Li, P.; Hao, S.; Zhang, Y.; Kim, S.W.; Li, H.; Ma, X.; Gao, S.; He, L.; Wu, W.; et al. Dynamic distribution of the gut microbiota and the relationship with apparent crude fiber digestibility and growth stages in pigs. Sci. Rep. 2015, 5. [Google Scholar] [CrossRef] [Green Version]


{kind=link}
{kind=link}
{kind=link}
| Items 1 | Growing Phase | Fattening Phase | ||
|---|---|---|---|---|
| Low Protein | Standard Protein | Low Protein | Standard Protein | |
| Ingredients (g/kg DM) | ||||
| Corn | 294.8 | 246.5 | 190.3 | 99.2 |
| Barley | 290.0 | 287.8 | 466.0 | 512.8 |
| Wheat | 200.0 | 200.0 | 200.0 | 200.0 |
| Soybean meal | 137.6 | 195.0 | 65.8 | 113.6 |
| Beet pulp dehydrated | 30.0 | 30.0 | 30.0 | 30.0 |
| Calcium carbonate | 13.4 | 13.2 | 10.0 | 10.0 |
| Mono calcium phosphate | 9.4 | 8.8 | 7.0 | 6.4 |
| Soybean oil | 9.0 | 6.3 | 17.0 | 17.1 |
| L-Valine | 6.8 | 8.0 | 0.0 | 0.0 |
| Sodium chloride | 4.6 | 4.6 | 4.1 | 4.1 |
| L-Lysine HCL | 4.2 | 2.4 | 3.7 | 2.1 |
| Vitamin-Mineral mix 2 | 4.0 | 4.0 | 4.0 | 4.0 |
| L-Threonine | 1.6 | 0.8 | 1.2 | 0.5 |
| DL- Methionine | 1.0 | 0.5 | 0.6 | 0.3 |
| L-Tryptophan | 0.3 | 0.2 | 0.2 | 0.0 |
| Chemical composition (g/kg DM) | ||||
| Dry matter (g/kg FM) | 900.1 | 897.1 | 909.9 | 908.4 |
| Crude protein | 147.2 | 166.9 | 129.27 | 154.95 |
| Ether extract | 24.9 | 26.9 | 39.7 | 43.2 |
| Ash | 65.8 | 70.3 | 58.1 | 52.8 |
| NDF | 173.3 | 179.9 | 211.8 | 211.6 |
| ME (kcal/kg) | 3108 | 3094 | 3185 | 3185 |
| Item 1 | Growing Phase | Fattening Phase | SEM | p-Value | |||
|---|---|---|---|---|---|---|---|
| Duroc | F2 | Duroc | F2 | PT | PP | ||
| n | 12 | 8 | 8 | 8 | |||
| Ileum 2 | |||||||
| Shannon index | 1.53 b | 1.64 ab | 1.47 b | 2.18 a | 0.160 | 0.014 | 0.186 |
| Simpson index | 0.61 b | 0.69 ab | 0.66 ab | 0.81 a | 0.049 | 0.009 | 0.102 |
| Richness 3 | 31.48 ab | 24.38 b | 31.50 ab | 43.88 a | 3.854 | 0.446 | 0.020 |
| Evenness | 0.45 | 0.52 | 0.43 | 0.58 | 0.040 | 0.011 | 0.785 |
| Cecum | |||||||
| Shannon index | 3.18 | 3.35 | 3.22 | 3.76 | 0.177 | 0.033 | 0.189 |
| Simpson index | 0.92 | 0.92 | 0.91 | 0.95 | 0.018 | 0.422 | 0.618 |
| Richness | 90.23 b | 104.88 ab | 106.50 ab | 134.50 a | 8.973 | 0.010 | 0.013 |
| Evenness | 0.71 | 0.72 | 0.70 | 0.77 | 0.034 | 0.200 | 0.457 |
| Ratio F/B | 47.57 | 24.40 | 35.68 | 2.37 | 18.409 | 0.089 | 0.309 |
| Distal colon | |||||||
| Shannon index | 3.53 | 3.83 | 3.67 | 3.77 | 0.128 | 0.076 | 0.650 |
| Simpson index | 0.94 | 0.96 | 0.94 | 0.95 | 0.009 | 0.047 | 0.742 |
| Richness | 123.64 | 140.00 | 146.13 | 153.72 | 9.595 | 0.138 | 0.045 |
| Evenness | 0.74 | 0.78 | 0.74 | 0.78 | 0.017 | 0.011 | 0.820 |
| Ratio F/B | 28.73 | 10.92 | 7.65 | 11.49 | 8.110 | 0.185 | 0.112 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sarri, L.; Costa-Roura, S.; Balcells, J.; Seradj, A.R.; de la Fuente, G. The Impact of Genetics on Gut Microbiota of Growing and Fattening Pigs under Moderate N Restriction. Animals 2021, 11, 2846. https://doi.org/10.3390/ani11102846
Sarri L, Costa-Roura S, Balcells J, Seradj AR, de la Fuente G. The Impact of Genetics on Gut Microbiota of Growing and Fattening Pigs under Moderate N Restriction. Animals. 2021; 11(10):2846. https://doi.org/10.3390/ani11102846
Chicago/Turabian StyleSarri, Laura, Sandra Costa-Roura, Joaquim Balcells, Ahmad Reza Seradj, and Gabriel de la Fuente. 2021. "The Impact of Genetics on Gut Microbiota of Growing and Fattening Pigs under Moderate N Restriction" Animals 11, no. 10: 2846. https://doi.org/10.3390/ani11102846
APA StyleSarri, L., Costa-Roura, S., Balcells, J., Seradj, A. R., & de la Fuente, G. (2021). The Impact of Genetics on Gut Microbiota of Growing and Fattening Pigs under Moderate N Restriction. Animals, 11(10), 2846. https://doi.org/10.3390/ani11102846