SPARCL1 Influences Bovine Skeletal Muscle-Derived Satellite Cell Migration and Differentiation through an ITGB1-Mediated Signaling Pathway


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Cell Culture and Differentiation
2.2. siRNA Interference
2.3. Cell Transfection
2.4. Cell Scratch Assay
2.5. Western Blotting
2.6. Co-Immunoprecipitation
- (1)
- Cells were washed 3 times with pre-cooled PBS to completely remove the culture medium.
- (2)
- Cells were then collected with 1 mL RIPA buffer (a pre-chilled cell scraper was used to gently scrape the cells from the 10 cm dish, ensuring clean cells). EP tube of the lysate was placed on ice and then on a horizontal shaker for 15–30 min.
- (3)
- Sample was centrifuged at 12,000 rpm for 30 min at 4 °C, and the supernatant was collected, while the precipitate was discarded. A total of 20 μL from the supernatant was taken as the input group.
- (4)
- The supernatant was divided into two parts, 500 μL each, where IgG and protein A/G were added (to remove non-specific binding in the experimental group), along with 1 μg rabbit IgG to the other group and placed on a horizontal shaker at 4 °C for 30 min–2 h.
- (5)
- The experimental group was centrifuged at 2500 rpm for 5 min. The precipitate was discarded while the supernatant was retained.
- (6)
- Approximately 0.2–2 μg primary antibody was added to the supernatant and shaken slowly overnight at 4 °C.
- (7)
- The column was then washed 3 times with PBS.
- (8)
- The column was added to the experimental group and the IgG group and shaken slowly on a horizontal shaker at 4 °C for 3–5 h.
- (9)
- Following shaking, it was centrifuged at 2500 rpm for 3 min and the supernatant was discarded.
- (10)
- The precipitate was then washed with protein lysis solution for 5 times.
- (11)
- We added 20–40 μL loading buffer and boiled it together with the input group for 10 min.
- (12)
- The sample was loaded on an SDS-PAGE at a concentration of 10%, and proteins were separated and transferred to PVDF membrane via blotting.
2.7. Immunofluorescence
2.8. Antibodies and Chemicals
2.9. Statistical Analysis
3. Results
3.1. SPARCL1 Interacts with ITGB1
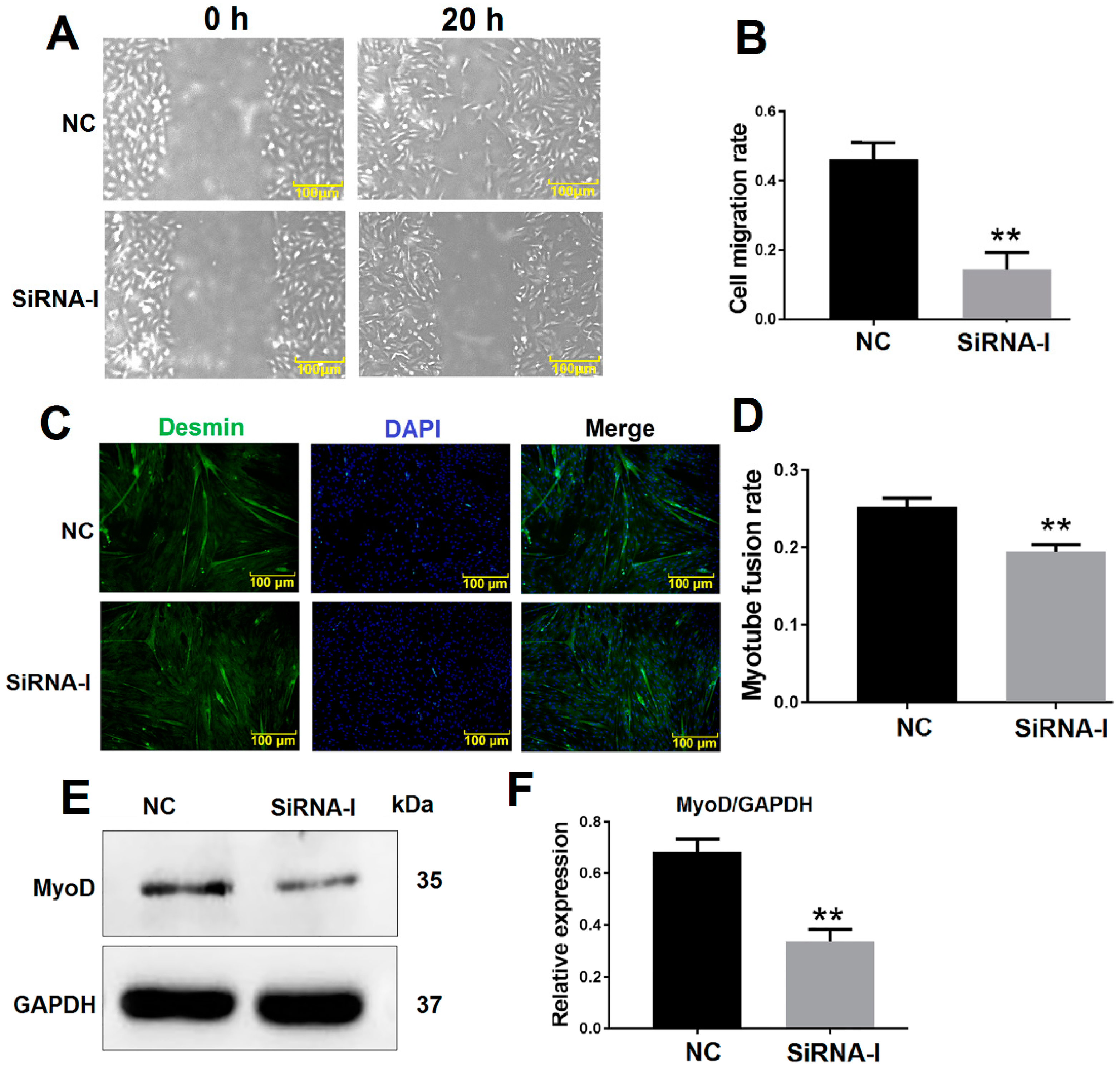
3.2. ITGB1 Influences MDSC Migration and Differentiation
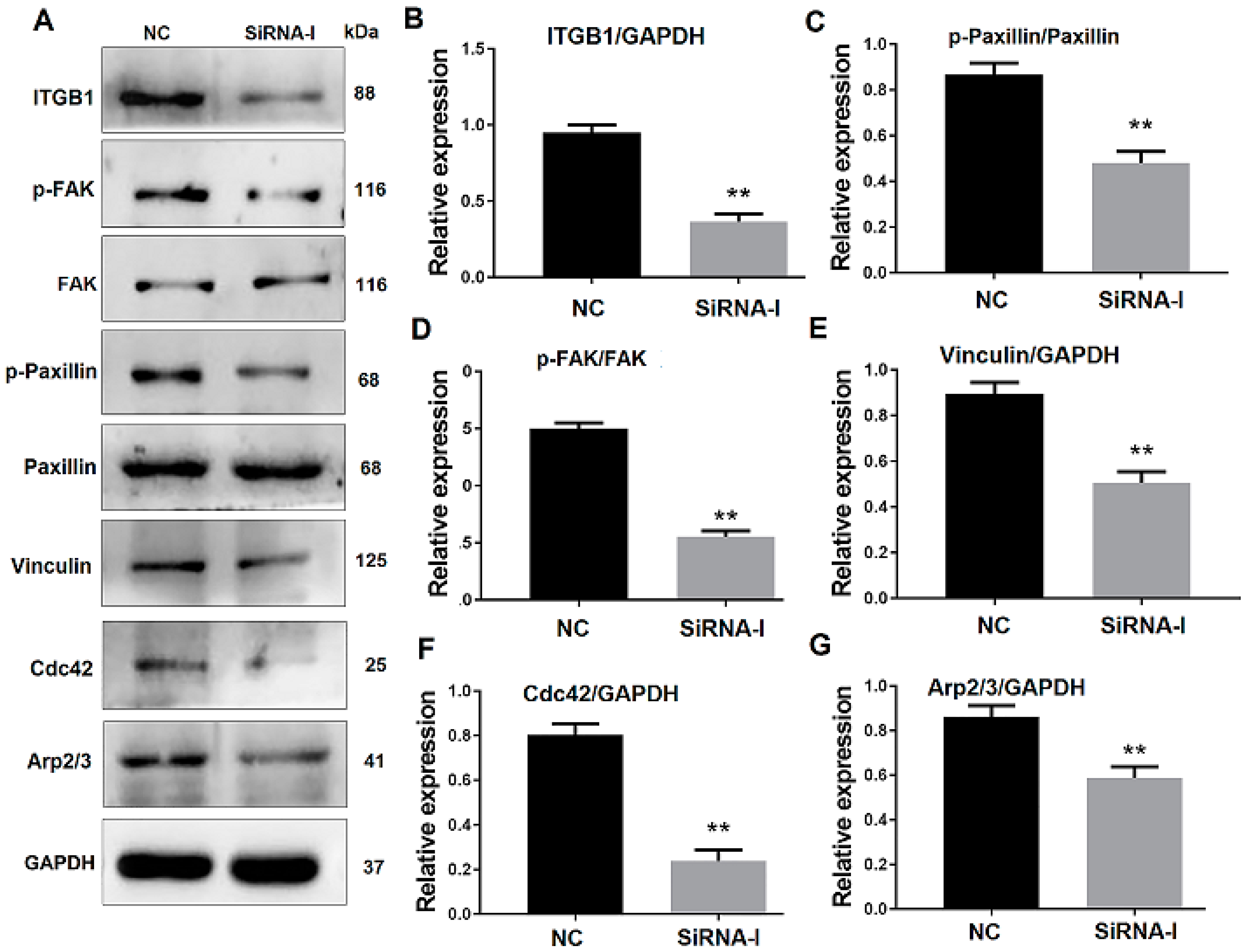
3.3. ITGB1 Inhibition Affects Downstream Protein Expression
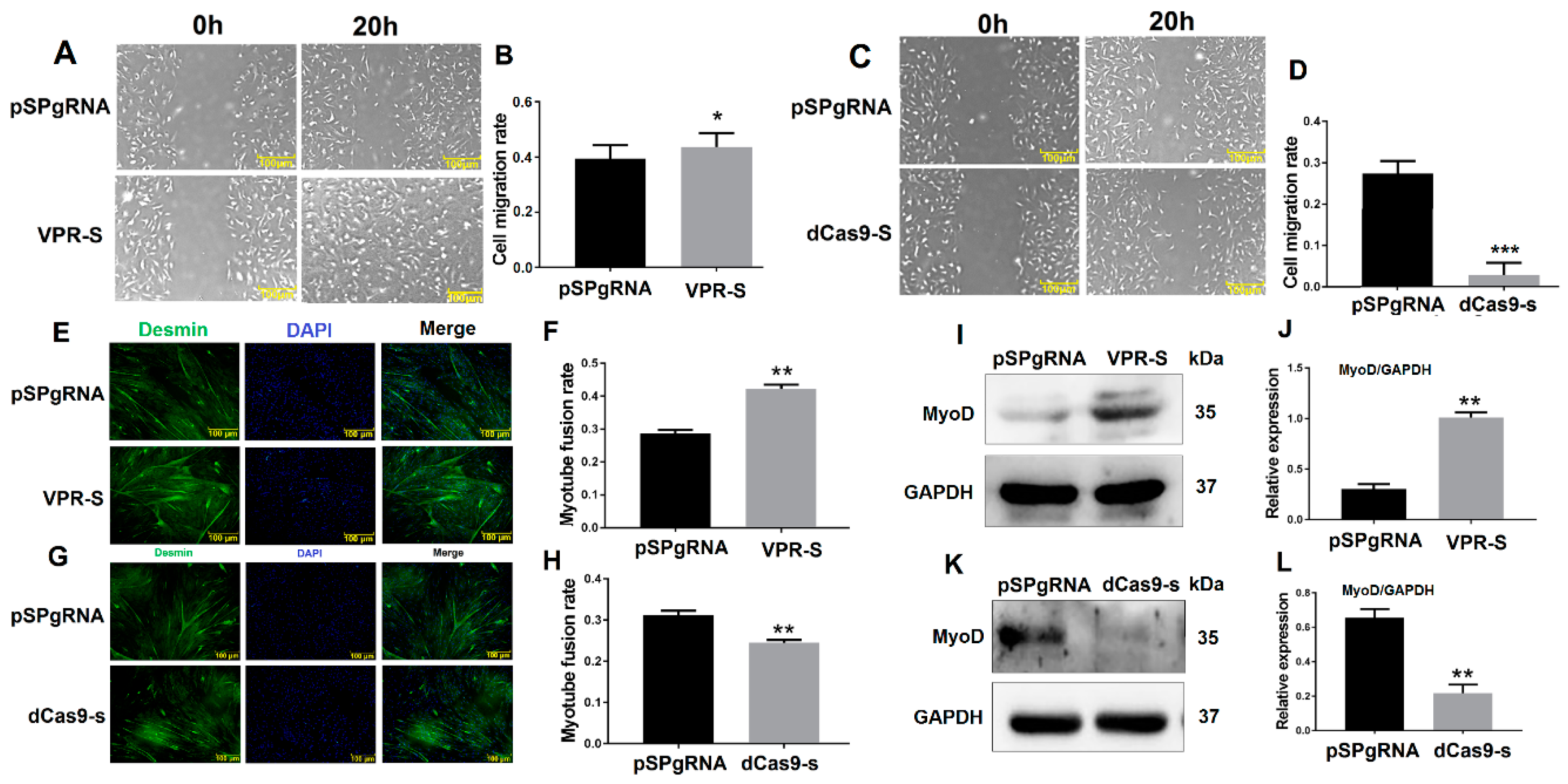
3.4. SPARCL1 Influences MDSC Migration and Differentiation
3.5. SPARCL1 Affects an ITGB1-Mediated Signaling Pathway
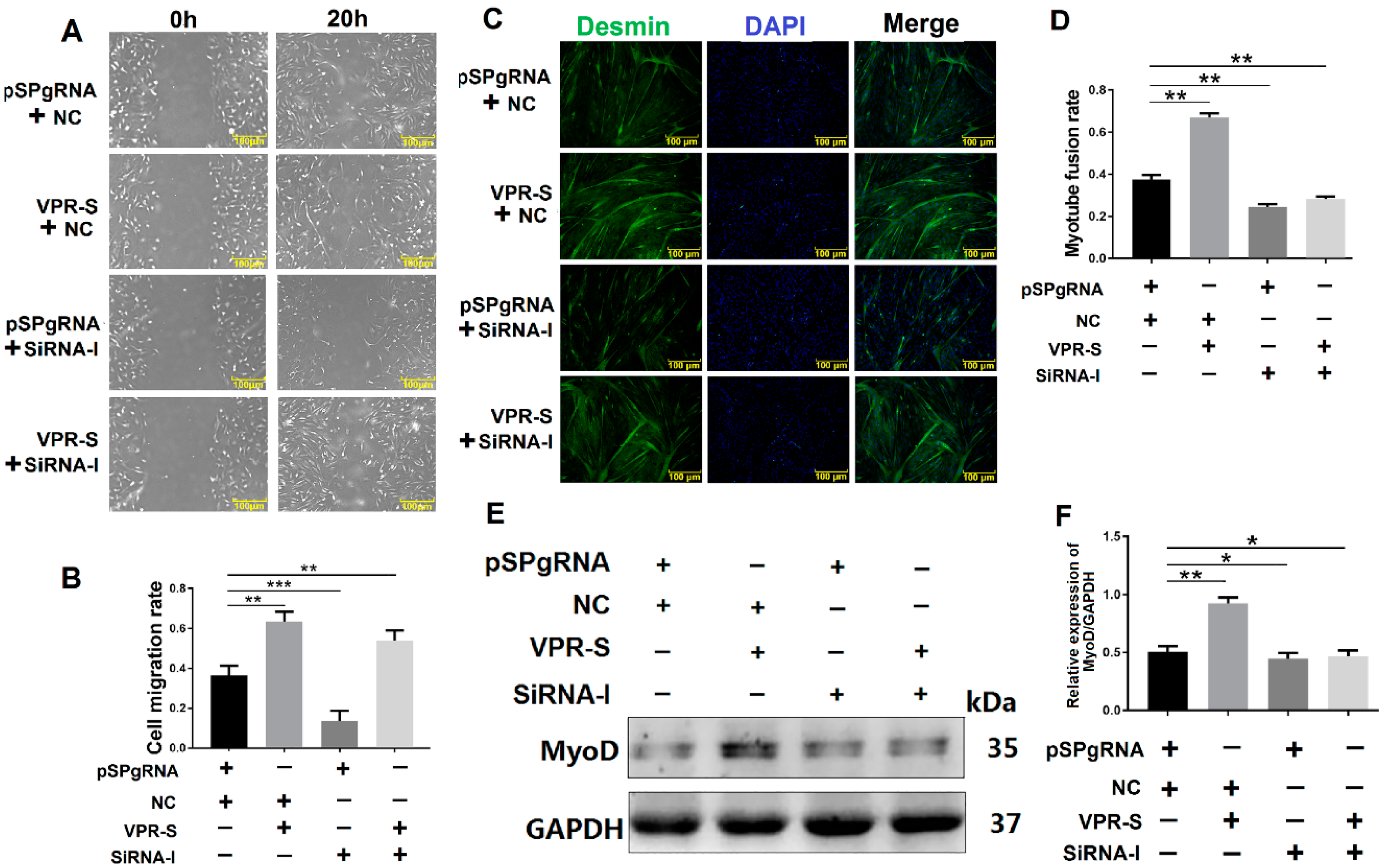
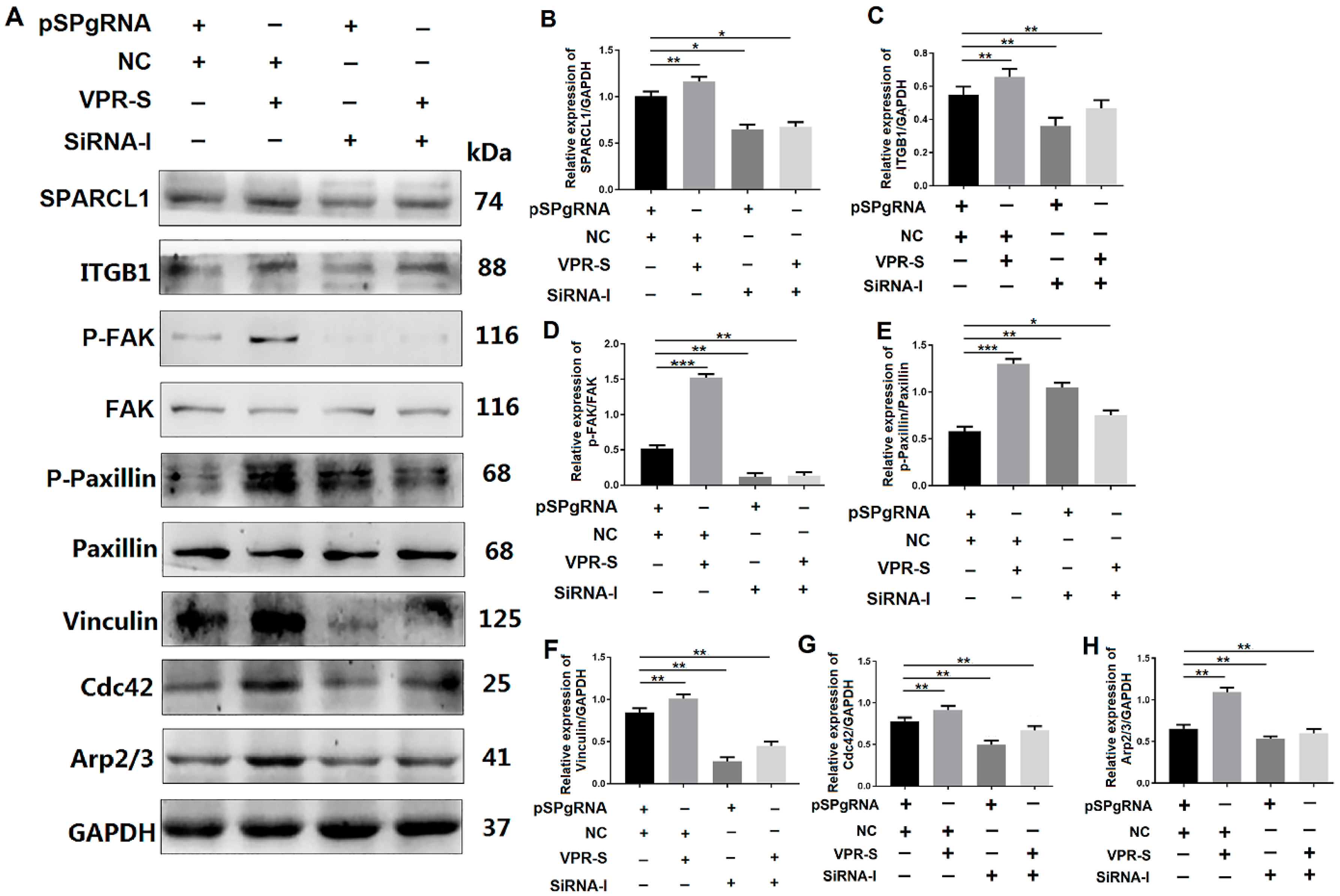
3.6. SPARCL1 Influences Cell Migration and Differentiation through an ITGB1-Mediated Cell Signaling Pathway
3.6.1. SPARCL1 Influences Cell Migration and Differentiation through ITGB1
3.6.2. SPARCL1 Regulates an ITGB1-Mediated Signaling Pathway through ITGB1
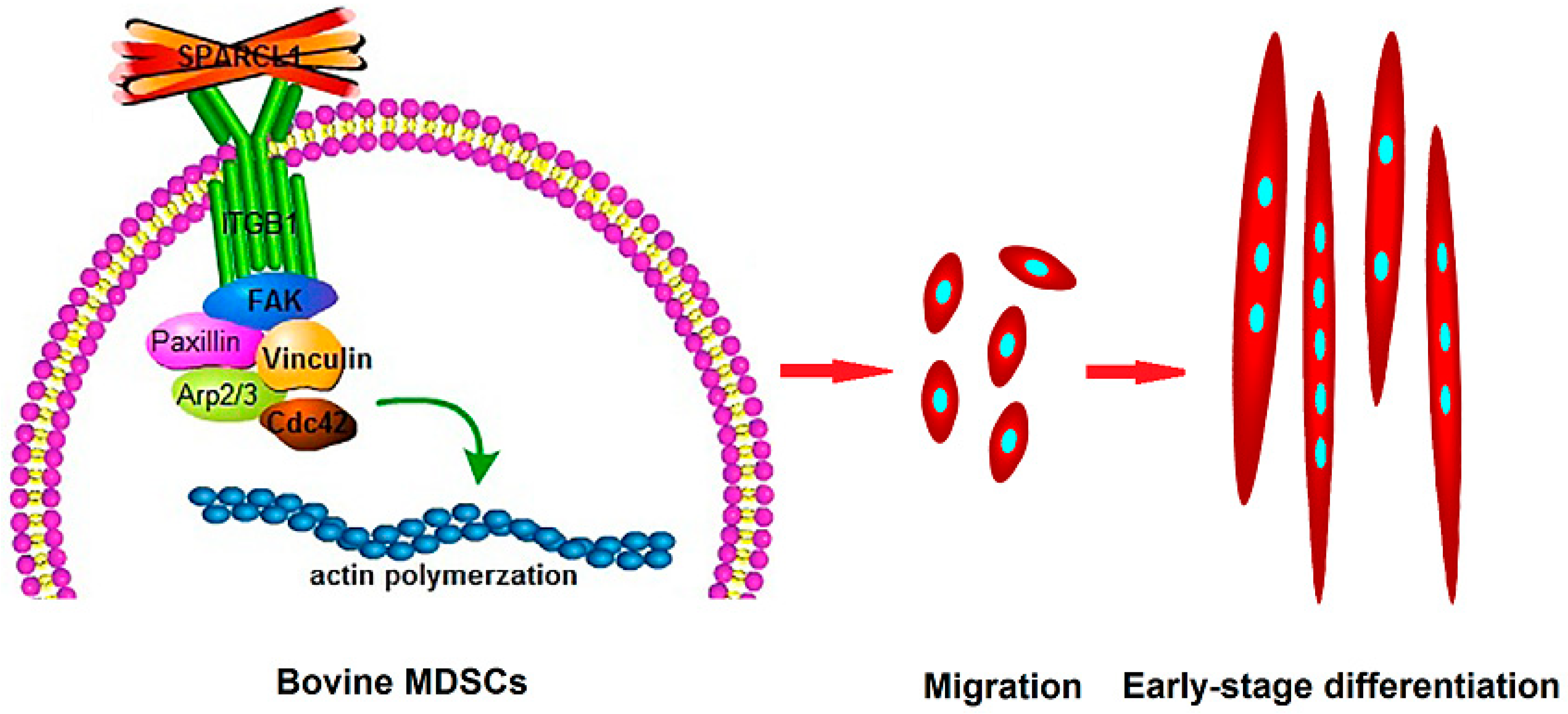
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Chetty, C.; Dontula, R.; Ganji, P.N.; Gujrati, M.; Lakka, S.S. SPARC expression induces cell cycle arrest via STAT3 signaling pathway in medulloblastoma cells. Biochem. Biophys. Res. Commun. 2012, 417, 874–879. [Google Scholar] [CrossRef] [PubMed]
- Yan, Q.; Sage, E.H. SPARC, a matricellular glycoprotein with important biological functions. J. Histochem. Cytochem. 1999, 47, 1495–1506. [Google Scholar] [CrossRef] [PubMed]
- Bradshaw, A.D.; Sage, E.H. SPARC, a matricellular protein that functions in cellular differentiation and tissue response to injury. J. Clin. Investig. 2001, 107, 1049–1054. [Google Scholar] [CrossRef] [PubMed]
- Jørgensen, L.H.; Petersson, S.J.; Sellathurai, J.; Andersen, D.C.; Thayssen, S.; Sant, D.J.; Jensen, C.H.; Schrøder, H.D. Secreted protein acidic and rich in cysteine (SPARC) in human skeletal muscle. J. Histochem. Cytochem. 2009, 57, 29–39. [Google Scholar] [CrossRef] [PubMed]
- Jørgensen, L.H.; Jepsen, P.L.; Boysen, A.; Dalgaard, L.B.; Hvid, L.G.; Ørtenblad, N.; Ravn, D.; Sellathurai, J.; Møller-Jensen, J.; Lochmüller, H.; et al. SPARC Interacts with Actin in Skeletal Muscle in Vitro and in Vivo. Am. J. Pathol. 2017, 187, 457–474. [Google Scholar] [CrossRef] [PubMed]
- Brekken, R.A.; Sullivan, M.M.; Workman, G.; Bradshaw, A.D.; Carbon, J.; Siadak, A.; Murri, C.; Framson, P.E.; Sage, E.H. Expression and characterization of murine hevin (SC1), a member of the SPARC family of matricellular proteins. J. Histochem. Cytochem. 2004, 52, 735–748. [Google Scholar] [CrossRef]
- Wang, Y.X.; Liu, S.Y.; Yan, Y.Q.; Li, S.F.; Tong, H.L. SPARCL1 promotes C2C12 cell differentiation via BMP7-mediated BMP/TGF-β cell signaling pathway. Cell Death Dis. 2019, 10, 852. [Google Scholar] [CrossRef]
- Liu, C.; Tong, H.; Li, S.; Yan, Y. Effect of ECM2 expression on bovine skeletal muscle-derived satellite cell differentiation. Cell Biol. Int. 2018, 42, 525–532. [Google Scholar] [CrossRef]
- Gros, J.; Manceau, M.; Thomé, V.; Marcelle, C. A common somatic origin for embryonic muscle progenitors and satellite cells. Nature 2005, 435, 954–958. [Google Scholar] [CrossRef]
- Dumont, N.A.; Bentzinger, C.F.; Sincennes, M.C.; Rudnicki, M.A. Satellite cells and skeletal muscle regeneration. Compr. Physiol. 2015, 5, 1027–1059. [Google Scholar]
- Mylona, E.; Jones, K.A.; Mills, S.T.; Pavlath, G.K. CD44 Regulates Myoblast Migration and Differentiation. J. Cell. Physiol. 2006, 209, 314–321. [Google Scholar] [CrossRef] [PubMed]
- Fry, C.S.; Kirby, T.J.; Kosmac, K.; McCarthy, J.J.; Peterson, C.A. Myogenic Progenitor Cells Control Extracellular Matrix Production by Fibroblasts during Skeletal Muscle Hypertrophy. Cell Stem Cell 2017, 20, 56–69. [Google Scholar] [CrossRef] [PubMed]
- Velleman, S.G.; McFarland, D.C. β1 Integrin mediation of myogenic differentiation: Implications for satellite cell differentiation. Poult. Sci. 2004, 83, 245–252. [Google Scholar] [CrossRef] [PubMed]
- Han, J.W.; Lee, H.J.; Bae, G.U.; Kang, J.S. Promyogenic function of Integrin/FAK signaling is mediated by Cdo, Cdc42 and MyoD. Cell. Signal. 2011, 23, 1162–1169. [Google Scholar] [CrossRef] [PubMed]
- Pang, Y.S.; Zhang, Z.H.; Wang, Z.; Wang, Y.X.; Yan, Y.Q.; Li, S.F.; Tong, H.L. Platelet endothelial aggregation receptor-1 regulates bovine muscle satellite cell migration and differentiation via integrin beta-1 and focal adhesion kinase. Cell Adhes. Migr. 2019, 13, 192–202. [Google Scholar] [CrossRef]
- Wu, C.; Asokan, S.B.; Berginski, M.E.; Haynes, E.M.; Sharpless, N.E.; Griffith, J.D.; Gomez, S.M.; Bear, J.E. Arp2/3 is critical for lamellipodia and response to extracellular matrix cues but is dispensable for chemotaxis. Cell 2012, 148, 973–987. [Google Scholar] [CrossRef]
- Rohatgi, R.; Ma, L.; Miki, H.; Lopez, M.; Kirchhausen, T.; Takenawa, T.; Kirschner, M.W. The interaction between N-WASP and the Arp2/3 complex links Cdc42-dependent signals to actin assembly. Cell 1999, 97, 221–231. [Google Scholar] [CrossRef]
- Burridge, K. Focal adhesions: A personal perspective on a half century of progress. FEBS J. 2017, 284, 3355–3361. [Google Scholar] [CrossRef]
- Bade, N.D.; Xu, T.; Kamien, R.D.; Assoian, R.K.; Stebe, K.J. Gaussian Curvature Directs Stress Fiber Orientation and Cell Migration. Biophys. J. 2018, 114, 1467–1476. [Google Scholar] [CrossRef]
- Yang, X.C.; Mei, S.T.; Niu, H.; Li, J.J. Nicotinic acid impairs assembly of leading edge in glioma cells. Oncol. Rep. 2017, 38, 829–836. [Google Scholar] [CrossRef]
- Hung, W.C.; Chen, S.H.; Paul, C.D.; Stroka, K.M.; Lo, Y.C.; Yang, J.T.; Konstantopoulos, K. Distinct signaling mechanisms regulate migration in unconfined versus confined spaces. J. Cell Biol. 2013, 202, 807–824. [Google Scholar] [CrossRef] [PubMed]
- Cho, W.J.; Kim, E.J.; Lee, S.J.; Kim, H.D.; Shin, H.J.; Lim, W.K. Involvement of SPARC in in vitro differentiation of skeletal myoblasts. Biochem. Biophys. Res. Commun. 2000, 271, 630–634. [Google Scholar] [CrossRef] [PubMed]
- Motamed, K.; Blake, D.J.; Angello, J.C.; Allen, B.L.; Rapraeger, A.C.; Hauschka, S.D.; Sage, E.H. Fibroblast growth factor receptor-1 mediates the inhibition of endothelial cell proliferation and the promotion of skeletal myoblast differentiation by SPARC: A role for protein kinase A. J. Cell Biochem. 2003, 90, 408–423. [Google Scholar] [CrossRef] [PubMed]
- Zhang, D.Y.; Xu, J.W.; Yang, P.; Wen, Y.F.; He, H.; Li, J.X.; Liang, J.T.; Zheng, Y.N.; Zhang, Z.J.; Wang, X.W.; et al. Genetic variant of SPARC gene and its association with growth traits in Chinese cattle. Arch. Anim. Breed. 2020, 63, 31–37. [Google Scholar] [CrossRef]
- Wang, N.; Chang, L.L. Maspin suppresses cell invasion and migration in gastric cancer through inhibiting EMT and angiogenesis via ITGB1/FAK pathway. Hum. Cell 2020, 33, 663–675. [Google Scholar] [CrossRef]
- Zheng, X.L.; Lv, W.G. Long noncoding RNA ITGB1 promotes migration and invasion of clear cell renal cell carcinoma by downregulating Mcl-1. Eur. Rev. Med. Pharmacol. Sci. 2019, 23, 1996–2002. [Google Scholar]
- Denes, L.T.; Riley, L.A.; Mijares, J.R.; Arboleda, J.D.; McKee, K.; Esser, K.A.; Wang, E.T. Culturing C2C12 myotubes on micromolded gelatin hydrogels accelerates myotube maturation. Skelet. Muscle 2019, 9, 17. [Google Scholar] [CrossRef]
- Yamamoto, M.; Legendre, N.P.; Biswas, A.A.; Lawton, A.; Yamamoto, S.; Tajbakhsh, S.; Kardon, G.; Goldhamer, D.J. Loss of MyoD and Myf5 in Skeletal Muscle Stem Cells Results in Altered Myogenic Programming and Failed Regeneration. Stem Cell Rep. 2018, 10, 956–969. [Google Scholar] [CrossRef]
- Ishido, M.; Kami, K.; Masuhara, M. In vivo expression patterns of MyoD, p21, and Rb proteins in myonuclei and satellite cells of denervated rat skeletal muscle. Am. J. Physiol. Cell Physiol. 2004, 287, C484–C493. [Google Scholar] [CrossRef]
- Charrier, E.E.; Montel, L.; Asnacios, A.; Delort, F.; Vicart, P.; Gallet, F.; Batonnet-Pichon, S.; Hénon, S. The desmin network is a determinant of the cytoplasmic stiffness of myoblasts. Biol. Cell 2018, 110, 77–90. [Google Scholar] [CrossRef]








© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wang, Y.; Liu, S.; Yan, Y.; Li, S.; Tong, H. SPARCL1 Influences Bovine Skeletal Muscle-Derived Satellite Cell Migration and Differentiation through an ITGB1-Mediated Signaling Pathway. Animals 2020, 10, 1361. https://doi.org/10.3390/ani10081361
Wang Y, Liu S, Yan Y, Li S, Tong H. SPARCL1 Influences Bovine Skeletal Muscle-Derived Satellite Cell Migration and Differentiation through an ITGB1-Mediated Signaling Pathway. Animals. 2020; 10(8):1361. https://doi.org/10.3390/ani10081361
Chicago/Turabian StyleWang, Yuxin, Shuaiyu Liu, Yunqin Yan, Shufeng Li, and Huili Tong. 2020. "SPARCL1 Influences Bovine Skeletal Muscle-Derived Satellite Cell Migration and Differentiation through an ITGB1-Mediated Signaling Pathway" Animals 10, no. 8: 1361. https://doi.org/10.3390/ani10081361
APA StyleWang, Y., Liu, S., Yan, Y., Li, S., & Tong, H. (2020). SPARCL1 Influences Bovine Skeletal Muscle-Derived Satellite Cell Migration and Differentiation through an ITGB1-Mediated Signaling Pathway. Animals, 10(8), 1361. https://doi.org/10.3390/ani10081361