Quantifying the Geometric Shell Shape between Populations of True Limpets Lottia Mesoleuca (Mollusca: Lottidae) in Colombia



Simple Summary
Abstract
1. Introduction
2. Materials and Methods
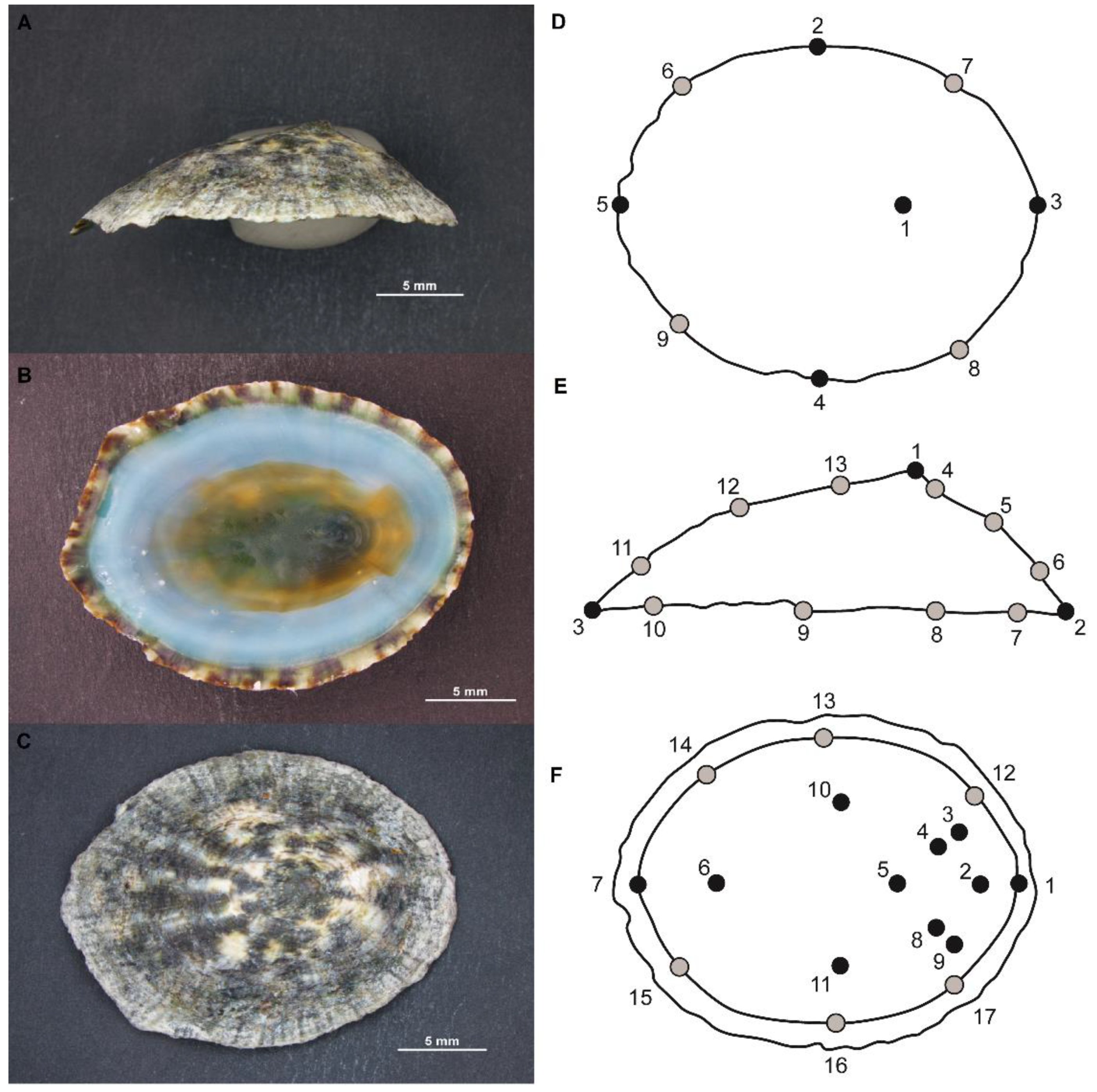
2.1. Sampling and Acquisition of Data
2.2. Morphometrics and Multivariate Analyses
3. Results
3.1. Morphological Component
Lottia Mesoleuca (Menke, 1851)
3.2. Morphological Variation
4. Discussion
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Benítez, H.A.; Püschel, T.A. Modelando la varianza de la forma: Morfometría geométrica aplicaciones en biología evolutiva. Int. J. Morphol. 2014, 32, 998–1008. [Google Scholar] [CrossRef]
- Adams, D.C.; Rohlf, F.J.; Slice, D.E. Geometric morphometrics: Ten years of progress following the ‘revolution’. Ital. J. Zool. 2004, 71, 5–16. [Google Scholar] [CrossRef]
- Adams, D.C.; Rohlf, F.J.; Slice, D.E. A field comes of age: Geometric morphometrics in the 21st century. Hystrix-Ital. J. Mammal. 2013, 24, 7–14. [Google Scholar] [CrossRef]
- Rohlf, F.J.; Marcus, L.F. A revolution in morphometrics. Trends Ecol. Evol. 1993, 8, 129–132. [Google Scholar] [CrossRef]
- Bookstein, F.L. Landmark methods for forms without landmarks: Morphometrics of group differences in outline shape. Med. Image Anal. 1997, 1, 225–243. [Google Scholar] [CrossRef]
- Bookstein, F.L. Size and shape spaces for landmark data in two dimensions. Stat. Sci. 1986, 1, 181–222. [Google Scholar] [CrossRef]
- Oxnard, C.; O’Higgins, P. Biology clearly needs morphometrics. Does morphometrics need biology? Biol. Theory 2009, 4, 84–97. [Google Scholar] [CrossRef]
- Gray, J.S. Marine biodiversity: Patterns, threats and conservation needs. Biodivers. Conserv. 1997, 6, 153–175. [Google Scholar] [CrossRef]
- Radulovici, A.E.; Archambault, P.; Dufresne, F. DNA barcodes for marine biodiversity: Moving fast forward? Diversity 2010, 2, 450–472. [Google Scholar] [CrossRef]
- Díaz, J.; Cantera, J.; Puyana, M. Estado actual del conocimiento en sistemática de moluscos marinos recientes de Colombia. Bol. Ecotrópica 1998, 33, 15–37. [Google Scholar]
- Ardilla, N.E.; Navas, G.R.C. Libro Rojo de Invertebrados Marinos de Colombia; Universidad Nacional de Colombia: Bogotá, Colombia, 2002. [Google Scholar]
- Branch, G.M. The biology of limpets: Physical factors, energy flow, and ecological interactions. Oceanogr. Mar. Biol. 1981, 19, 235–379. [Google Scholar]
- Branch, G. Limpets: Their role in littoral and sublittoral community dynamics. In the Ecology of Rocky Coast; Moore, P.G., Seeds, R., Eds.; Hodder and Stoughton: London, UK, 1985; pp. 97–116. [Google Scholar]
- Nakano, T.; Ozawa, T. Worldwide phylogeography of limpets of the order Patellogastropoda: Molecular, morphological and palaeontological evidence. J. Molluscan Stud. 2007, 73, 79–99. [Google Scholar] [CrossRef]
- Nakano, T.; Yazaki, I.; Kurokawa, M.; Yamaguchi, K.; Kuwasawa, K. The origin of the endemic patellogastropod limpets of the Ogasawara Islands in the northwestern Pacific. J. Molluscan Stud. 2009, 75, 87–90. [Google Scholar] [CrossRef]
- Goldstien, S.J.; Gemmell, N.J.; Schiel, D.R. Colonisation and connectivity by intertidal limpets among New Zealand, Chatham and Sub-Antarctic Islands. I. Genetic connections. Mar. Ecol. Prog. Ser. 2009, 388, 111–119. [Google Scholar] [CrossRef]
- Suter, H. Acmaeidae of New Zealand. Proc. Malacol. Soc. Lond. 1907, 7, 324–326. [Google Scholar]
- Oliver, W.R.B. Australasian Patelloididae; WAG Skinner: Wellington, New Zealand, 1926. [Google Scholar]
- Powell, A. The patellid limpets of the world (Patellidae). Indo-Pasific Mollusca 1973, 3, 75–206. [Google Scholar]
- Ponder, W.; Creese, R. A revision of the Australian species of Notoacmea, Collisella and Patelloida (Mollusca: Gastropoda: Acmaeidae). J. Malacol. Soc. Aust. 1980, 4, 167–208. [Google Scholar] [CrossRef]
- Sasaki, K. Johnius (johnieops) philippinus, a new sciaenid from the philippines, with a synopsis of species included in the subgenusjohnieops. Ichthyol. Res. 1999, 46, 271–279. [Google Scholar] [CrossRef]
- Walker, A. Introduction to the ecology of the Antarctic limpet Patinigera polaris (Hombron and Jacquinot) as Signy Islands, South Orkney Islands. Br. Antarct. Surv. Bull. 1972, 28, 49–69. [Google Scholar]
- Nolan, C.P. Size, shape and shell morphology in the Antarctic limpet Nacella concinna at Signy Island, South Orkney Islands. J. Molluscan Stud. 1991, 57, 225–238. [Google Scholar] [CrossRef]
- Weihe, E.; Abele, D. Differences in the physiological response of inter-and subtidal Antarctic limpets Nacella concinna to aerial exposure. Aquat. Biol. 2008, 4, 155–166. [Google Scholar] [CrossRef]
- Coleman, R.A.; Browne, M.; Theobalds, T. Aggregation as a defense: Limpet tenacity changes in response to simulated predator attack. Ecology 2004, 85, 1153–1159. [Google Scholar] [CrossRef]
- Iwasaki, K. Analyses of limpet defense and predator offense in the field. Mar. Biol. 1993, 116, 277–289. [Google Scholar] [CrossRef]
- Silva, A.; Hawkins, S.; Boaventura, D.; Thompson, R. Predation by small mobile aquatic predators regulates populations of the intertidal limpet Patella vulgata (L.). J. Exp. Mar. Biol. Ecol. 2008, 367, 259–265. [Google Scholar] [CrossRef]
- Albarrán-Mélzer, N.C.; Ruiz, L.J.R.; Benítez, H.A.; Lagos, M.E. Can temperature shift morphological changes of invasive species? A morphometric approach on the shells of two tropical freshwater snail species. Hydrobiologia 2020, 847, 151–160. [Google Scholar] [CrossRef]
- Gouveia, N.; Oliveira, C.R.; Martins, C.P.; Maranho, L.A.; Pereira, C.D.S.; Manoela, R.; Harayashiki, C.A.; Almeida, S.M.; Castro, I.B. Can shell alterations in limpets be used as alternative biomarkers of coastal contamination? Chemosphere 2019, 224, 9–19. [Google Scholar] [CrossRef] [PubMed]
- Tamburi, N.E.; Seuffert, M.E.; Martín, P.R. Temperature-induced plasticity in morphology and relative shell weight in the invasive apple snail Pomacea canaliculata. J. Therm. Biol. 2018, 74, 331–336. [Google Scholar] [CrossRef] [PubMed]
- Cabral, J.P.; Jorge, R.N. Compressibility and shell failure in the European Atlantic Patella limpets. Mar. Biol. 2007, 150, 585–597. [Google Scholar] [CrossRef]
- Lindberg, D.R.; Pearse, J.S. Experimental manipulation of shell color and morphology of the limpets Lottia asmi (Middendorff) and Lottia digitalis (Rathke)(Mollusca: Patellogastropoda). J. Exp. Marine Biol. Ecol. 1990, 140, 173–185. [Google Scholar] [CrossRef]
- Zúniga-Reinoso, Á.; Benítez, H.A. The overrated use of the morphological cryptic species concept: An example with Nyctelia darkbeetles (Coleoptera: Tenebrionidae) using geometric morphometrics. Zool. Anz. A J. Comp. Zool. 2015, 255, 47–53. [Google Scholar] [CrossRef]
- Djurakic, M.; Milankov, V. The utility of plastron shape for uncovering cryptic diversity in Hermann’s tortoise. J. Zool. 2020, 310, 145–157. [Google Scholar] [CrossRef]
- Gunz, P.; Mitteroecker, P. Semilandmarks: A method for quantifying curves and surfaces. Hystrix-Ital. J. Mammal. 2013, 24, 103–109. [Google Scholar] [CrossRef]
- Mitteroecker, P.; Gunz, P. Advances in geometric morphometrics. Evol. Biol. 2009, 36, 235–247. [Google Scholar] [CrossRef]
- Keen, A.M. Sea shells of tropical West America; Stanford University Press: Standford, CA, USA, 1971. [Google Scholar]
- Rohlf, F.J. TPSdig, v. 2.17, State University at Stony Brook: New York, NY, USA, 2013.
- Rohlf, F.J.; Slice, D. Extensions of the Procustes methods for the optimal superimposition of landmarks. Syst. Zool. 1990, 39, 40–59. [Google Scholar] [CrossRef]
- Dryden, I.L.; Mardia, K.V. Statistical Shape Analysis; Wiley: Chichester, UK, 1998; Volume 4. [Google Scholar]
- R Core Team R: A language and environment for statistical computing. Available online: https://www.r-project.org/ (accessed on 11 April 2020).
- Adams, D.C.; Otárola-Castillo, E. Geomorph: An R package for the collection and analysis of geometric morphometric shape data. Methods Ecol. Evol. 2013, 4. [Google Scholar] [CrossRef]
- Jolliffe, I.T. Principal Component Analysis, 2nd ed.; Springer: New York, NY, USA, 2002. [Google Scholar]
- Benítez, H.A.; Parra, L.E.; Sepulveda, E.; Sanzana, M.J. Geometric perspectives of sexual dimorphism in the wing shape of lepidoptera: The case of Synneuria sp (Lepidoptera: Geometridae). J. Entomol. Res. Soc. 2011, 13, 53–60. [Google Scholar]
- Lemic, D.; Benítez, H.A.; Bažok, R. Intercontinental effect on sexual shape dimorphism and allometric relationships in the beetle pest Diabrotica virgifera virgifera LeConte (Coleoptera: Chrysomelidae). Zool. Anz. A J. Comp. Zool. 2014, 253, 203–206. [Google Scholar] [CrossRef]
- Klingenberg, C.P. MorphoJ: An integrated software package for geometric morphometrics. Mol. Ecol. Resour. 2011, 11, 353–357. [Google Scholar] [CrossRef]
- Püschel, T.A.; Espejo, J.; Sanzana, M.-J.; Benitez, H.A. Analysing the floral elements of the lost tree of Easter Island: A morphometric comparison between the remaining ex-situ lines of the endemic extinct species Sophora toromiro. PLoS ONE 2014, 9. [Google Scholar] [CrossRef]
- Morey, L.C.; Blashfield, R.K.; Skinner, H.A. A comparison of cluster analysis techniques withing a sequential validation framework. Multivar. Behav. Res. 1983, 18, 309–329. [Google Scholar] [CrossRef]
- Orton, J. Some limiting factors in the environment of the common limpet, P. vulgata. Nature 1933, 131, 693–694. [Google Scholar] [CrossRef]
- Wanninger, A.; Ruthensteiner, B.; Lobenwein, S.; Salvenmoser, W.; Dictus, W.J.; Haszprunar, G. Development of the musculature in the limpet Patella (Mollusca, Patellogastropoda). Dev. Genes Evol. 1999, 209, 226–238. [Google Scholar] [CrossRef] [PubMed]
- Cabral, J.P. Shape and growth in European Atlantic Patella limpets (Gastropoda, Mollusca). Ecological implications for survival. Web Ecol. 2007, 7, 11–21. [Google Scholar] [CrossRef]
- Adams, D.C.; Rohlf, F.J. Ecological character displacement in Plethodon: Biomechanical differences found from a geometric morphometric study. Proc. Natl. Acad. Sci. USA 2000, 97, 4106–4111. [Google Scholar] [CrossRef]
- Lemic, D.; Benítez, H.A.; Püschel, T.A.; Gašparić, H.V.; Šatvar, M.; Bažok, R. Ecological morphology of the sugar beet weevil Croatian populations: Evaluating the role of environmental conditions on body shape. Zool. Anz. A J. Comp. Zool. 2016, 260, 25–32. [Google Scholar] [CrossRef]
- Mikac, K.M.; Lemic, D.; Benítez, H.A.; Bažok, R. Changes in corn rootworm wing morphology are related to resistance development. J. Pest Sci. 2019, 92, 443–451. [Google Scholar] [CrossRef]
- Mikac, K.M.; Lemic, D.; Bažok, R.; Benítez, H.A. Wing shape changes: A morphological view of the Diabrotica virgifera virgifera European invasion. Biol. Invasions 2016, 18, 3401–3407. [Google Scholar] [CrossRef]
- Benítez, H.A.; Vargas, H.A.; Püschel, T.A. Left–right asymmetry and morphological consequences of a host shift in the oligophagous Neotropical moth Macaria mirthae (Lepidoptera: Geometridae). J. Insect Conserv. 2015, 19, 589–598. [Google Scholar] [CrossRef]
- Klingenberg, C.P. Evolution and development of shape: Integrating quantitative approaches. Nat. Rev. Genet. 2010, 11, 623–635. [Google Scholar] [CrossRef]
- McQuaid, C. Biology of the gastropod family Littorinidae; I. Evolutionary aspects. Oceanogr. Mar. Biol. Ann. Rev. 1996, 34, 233–262. [Google Scholar]
- Parsonage, S.; Hughes, J. Natural selection and the distribution of shell colour morphs in three species of Littoraria (Gastropoda: Littorinidae) in Moreton Bay, Queensland. Biol. J. Linn. Soc. 2002, 75, 219–232. [Google Scholar] [CrossRef]
- Cole, T.J. Inheritance of juvenile shell colour of the oyster drill Urosalpinx cinerea. Nature 1975, 257, 794–795. [Google Scholar] [CrossRef] [PubMed]
- Underwood, A.; Creese, R. Observations on the biology of the trochid gastropod Austrocochlea constricta (Lamarck)(Prosobranchia). II. The effects of available food on shell-banding pattern. J. Exp. Mar. Biol. Ecol. 1976, 23, 229–240. [Google Scholar] [CrossRef]
- Reimchen, T. Substratum heterogeneity, crypsis, and colour polymorphism in an intertidal snail (Littorina mariae). Can. J. Zool. 1979, 57, 1070–1085. [Google Scholar] [CrossRef]
- Atkinson, W.; Warwick, T. The role of selection in the colour polymorphism of Littorina rudis Maton and Littorina arcana Hannaford-Ellis (Prosobranchia: Littorinidae). Biol. J. Linn. Soc. 1983, 20, 137–151. [Google Scholar] [CrossRef]
- Vermeij, G.J.; Carlson, S.J. The muricid gastropod subfamily Rapaninae: Phylogeny and ecological history. Paleobiology 2000, 26, 19–46. [Google Scholar] [CrossRef]
- Collin, R. The utility of morphological characters in gastropod phylogenetics: An example from the Calyptraeidae. Biol. J. Linn. Soc. 2003, 78, 541–593. [Google Scholar] [CrossRef]
- Teske, P.R.; Barker, N.P.; McQuaid, C.D. Lack of genetic differentiation among four sympatric southeast African intertidal limpets (Siphonariidae): Phenotypic plasticity in a single species? J. Molluscan Stud. 2007, 73, 223–228. [Google Scholar] [CrossRef]
- Demes, K.W.; Graham, M.H.; Suskiewicz, T.S. Phenotypic plasticity reconciles incongruous molecular and morphological taxonomies: The giant kelp, Macrocystis (Laminariales, Phaeophyceae), is a monospecific genus. J. Phycol. 2009, 45, 1266–1269. [Google Scholar] [CrossRef]
- Reisser, C.M.; Wood, A.R.; Bell, J.J.; Gardner, J.P. Connectivity, small islands and large distances: The Cellana strigilis limpet complex in the Southern Ocean. Mol. Ecol. 2011, 20, 3399–3413. [Google Scholar] [CrossRef]
- Appleton, R.D.; Palmer, A.R. Water-borne stimuli released by predatory crabs and damaged prey induce more predator-resistant shells in a marine gastropod. Proc. Natl. Acad. Sci. USA 1988, 85, 4387–4391. [Google Scholar] [CrossRef] [PubMed]
- DeWitt, T.J.; Robinson, B.W.; Wilson, D.S. Functional diversity among predators of a freshwater snail imposes an adaptive trade-off for shell morphology. Evol. Ecol. Res. 2000, 2, 129–148. [Google Scholar]
- Lakowitz, T.; Brönmark, C.; Nyström, P. Tuning in to multiple predators: Conflicting demands for shell morphology in a freshwater snail. Freshw. Biol. 2008, 53, 2184–2191. [Google Scholar] [CrossRef]
- Hoverman, J.T.; Relyea, R.A. The long-term impacts of predators on prey: Inducible defenses, population dynamics, and indirect effects. Oikos 2012, 121, 1219–1230. [Google Scholar] [CrossRef]
- Hoverman, J.T.; Relyea, R.A. Survival trade-offs associated with inducible defences in snails: The roles of multiple predators and developmental plasticity. Funct. Ecol. 2009, 23, 1179–1188. [Google Scholar] [CrossRef]
- Anistratenko, V. Switching of early ontogeny type, its mechanism, and role in evolution of mollusca. Biol. Bull. 2010, 37, 130–137. [Google Scholar] [CrossRef]
- Kolbin, K.; Kulikova, V. Reproduction and larval development of the limpet Lottia persona (Rathke, 1833)(Gastropoda: Lottiidae). Russ. J. Mar. Biol. 2011, 37, 239–242. [Google Scholar] [CrossRef]
- Frýda, J. Phylogeny of Palaeozoic gastropods inferred from their ontogeny. In Earth and Life; Springer: Berlin/Heidelberg, Germany, 2012; pp. 395–435. [Google Scholar]
- Telesca, L.; Michalek, K.; Sanders, T.; Peck, L.S.; Thyrring, J.; Harper, E.M. Blue mussel shell shape plasticity and natural environments: A quantitative approach. Sci. Rep. 2018, 8, 2865. [Google Scholar] [CrossRef]
- Avaca, M.S.; Narvarte, M.; Martín, P.; Van Der Molen, S. Shell shape variation in the Nassariid Buccinanops globulosus in northern Patagonia. Helgol. Mar. Res. 2013, 67, 567–577. [Google Scholar] [CrossRef]
- Hall-Spencer, J.M.; Rodolfo-Metalpa, R.; Martin, S.; Ransome, E.; Fine, M.; Turner, S.M.; Rowley, S.J.; Tedesco, D.; Buia, M.-C. Volcanic carbon dioxide vents show ecosystem effects of ocean acidification. Nature 2008, 454, 96–99. [Google Scholar] [CrossRef]
- Menge, B.A.; Sutherland, J.P. Species diversity gradients: Synthesis of the roles of predation, competition, and temporal heterogeneity. Am. Nat. 1976, 110, 351–369. [Google Scholar] [CrossRef]
- Kaliontzopoulou, A.; Carretero, M.A.; Llorente, G.A. Intraspecific ecomorphological variation: Linear and geometric morphometrics reveal habitat-related patterns within Podarcis bocagei wall lizards. J. Evol. Biol. 2010, 23, 1234–1244. [Google Scholar] [CrossRef] [PubMed]
- Sherratt, E.; Anstis, M.; Keogh, J.S. Ecomorphological diversity of Australian tadpoles. Ecol. Evol. 2018, 8, 12929–12939. [Google Scholar] [CrossRef] [PubMed]
- Benítez, H.A.; Püschel, T.; Lemic, D.; Čačija, M.; Kozina, A.; Bažok, R. Ecomorphological variation of the wireworm cephalic capsule: Studying the interaction of environment and geometric shape. PLoS ONE 2014, 9. [Google Scholar] [CrossRef] [PubMed]
- De Esteban-Trivigno, S. Ecomorphology of extinct Xenarthrans: Analysis of the mandible using geometric morphometrics methods. Ameghiniana 2011, 48, 381–398. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Population | Isla Gorgona (Proc. p Value) | Isla Palma (Proc. p Value) | Los Negritos (Proc. p Value) |
|---|---|---|---|
| Lateral | |||
| Isla Palma | 0.0324 (<0.0001) | ||
| Los Negritos | 0.0199 (0.5609) | 0.0232 (0.1318) | |
| Morrochiquito | 0.0364 (<0.0001) | 0.0178 (0.0340) | 0.0263 (0.0929) |
| Dorsal | |||
| Isla Palma | 0.0412 (<0.0001) | ||
| Los Negritos | 0.0689 (0.0001) | 0.0407 (0.0001) | |
| Morrochiquito | 0.0539 (<0.0001) | 0.0309 (<0.0001) | 0.0226 (0.2673) |
| Ventral | |||
| Isla Palma | 0.0149 (0.0006) | ||
| Los Negritos | 0.0228 (0.0851) | 0.0165 (0.1102) | |
| Morrochiquito | 0.0226 (0.0004) | 0.0128 (0.0087) | 0.0252 (0.0027) |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Echeverry, A.M.; Londoño-Cruz, E.; Benítez, H.A. Quantifying the Geometric Shell Shape between Populations of True Limpets Lottia Mesoleuca (Mollusca: Lottidae) in Colombia. Animals 2020, 10, 675. https://doi.org/10.3390/ani10040675
Echeverry AM, Londoño-Cruz E, Benítez HA. Quantifying the Geometric Shell Shape between Populations of True Limpets Lottia Mesoleuca (Mollusca: Lottidae) in Colombia. Animals. 2020; 10(4):675. https://doi.org/10.3390/ani10040675
Chicago/Turabian StyleEcheverry, Ana María, Edgardo Londoño-Cruz, and Hugo A. Benítez. 2020. "Quantifying the Geometric Shell Shape between Populations of True Limpets Lottia Mesoleuca (Mollusca: Lottidae) in Colombia" Animals 10, no. 4: 675. https://doi.org/10.3390/ani10040675
APA StyleEcheverry, A. M., Londoño-Cruz, E., & Benítez, H. A. (2020). Quantifying the Geometric Shell Shape between Populations of True Limpets Lottia Mesoleuca (Mollusca: Lottidae) in Colombia. Animals, 10(4), 675. https://doi.org/10.3390/ani10040675