
Effects of Age, Diet CP, NDF, EE, and Starch on the Rumen Bacteria Community and Function in Dairy Cattle

 , and
, and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Ethics Statements
2.2. Animals and Sample Collection
2.3. Sample Analysis
2.3.1. Fermentation Profile and Enzyme Activity
2.3.2. 16S rRNA Sequencing
2.4. Statistics
3. Results
3.1. Rumen Fermentation Profile and Enzyme Activity
3.2. Rumen Bacteria Analysis
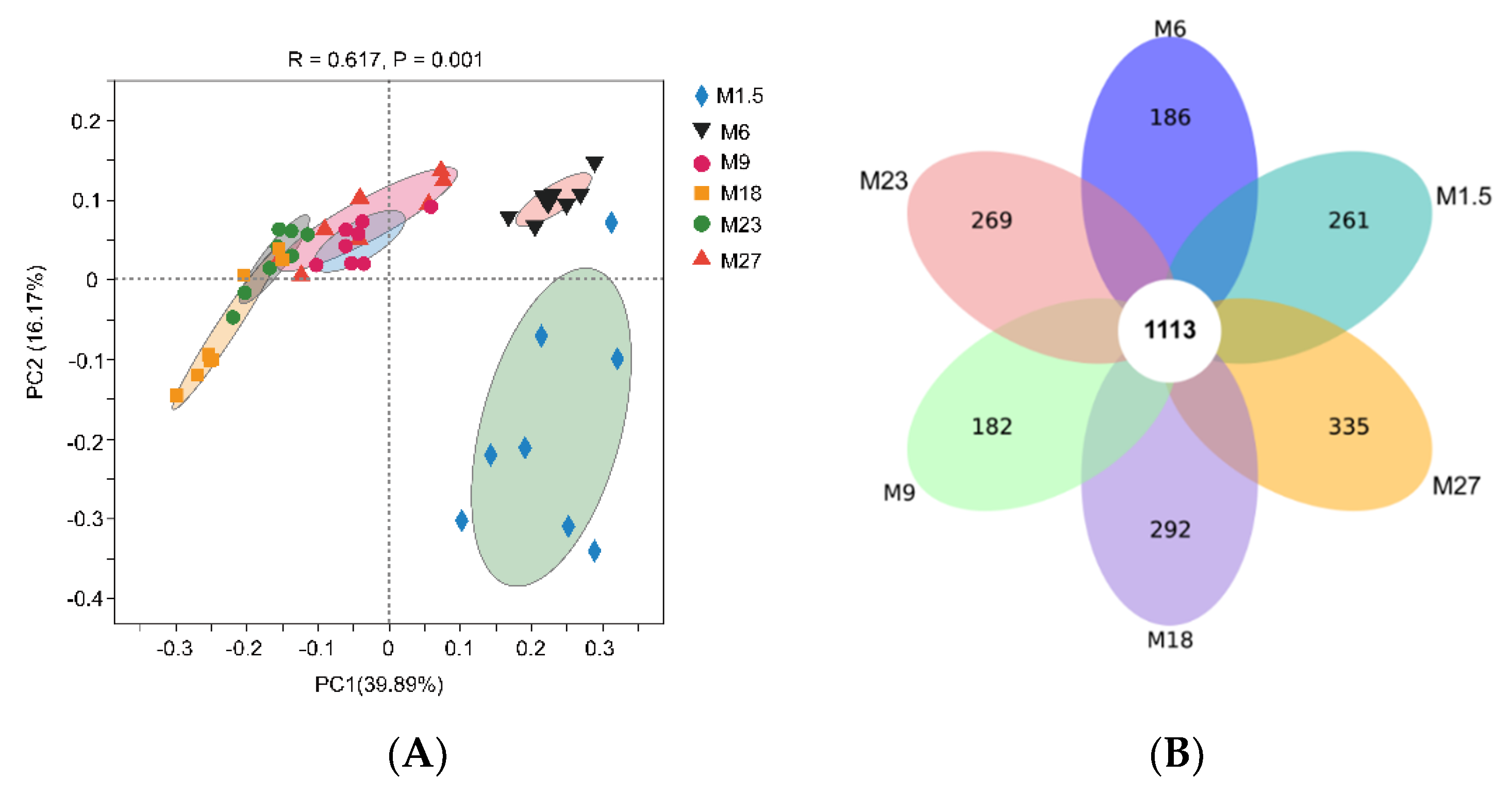
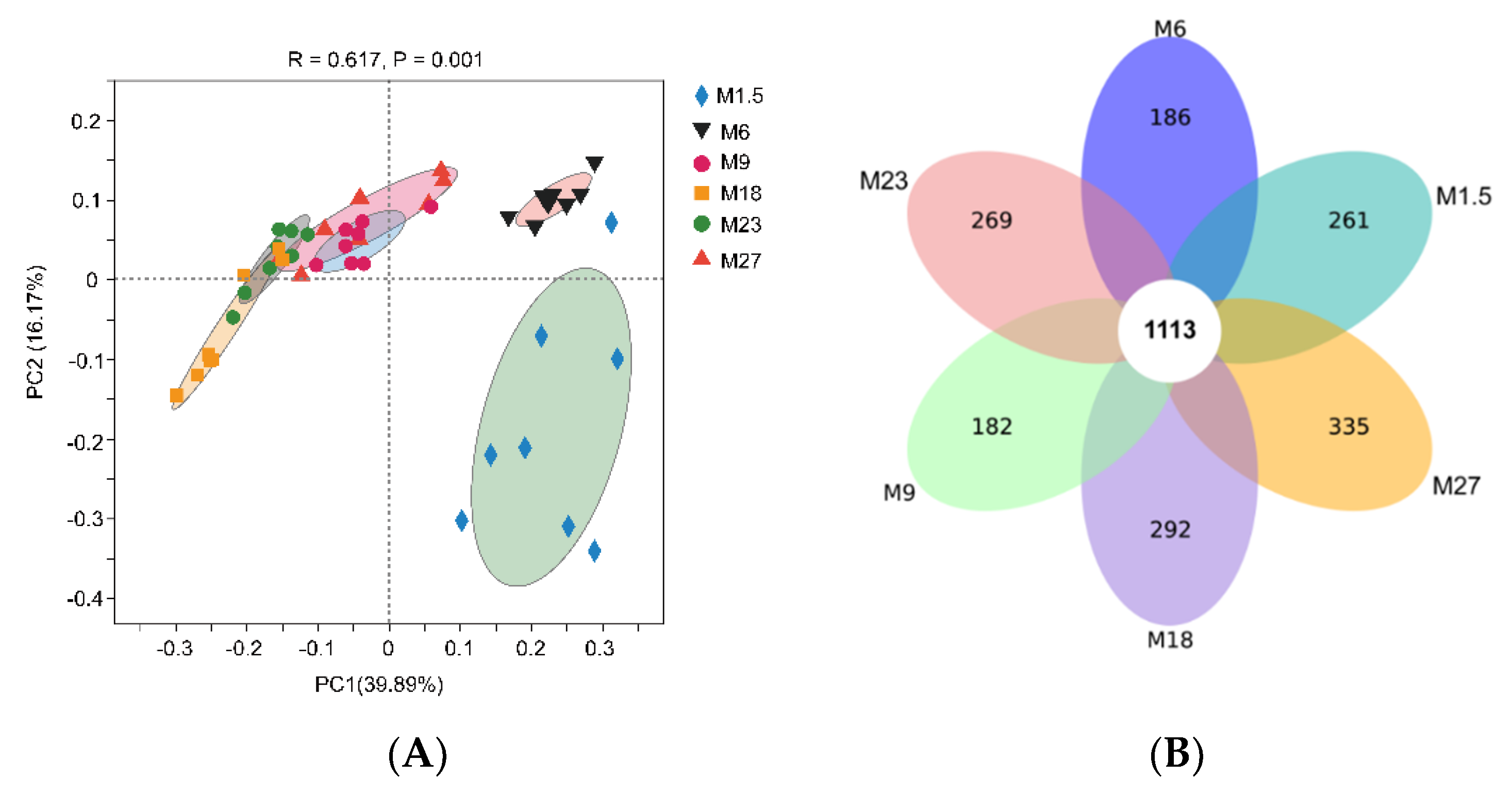
3.2.1. Rumen Bacteria Diversity Analysis
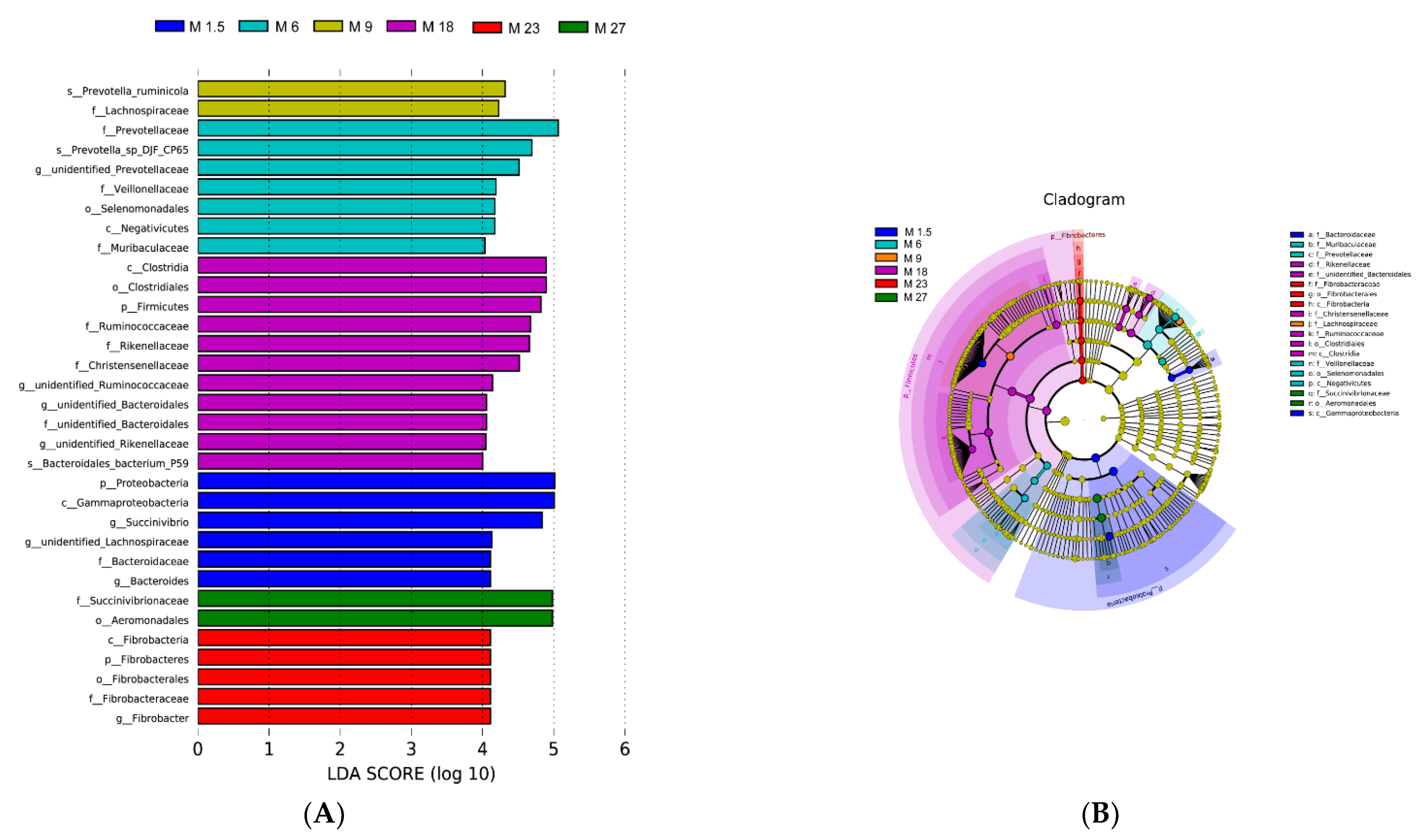
3.2.2. Rumen Bacteria Composition Analysis
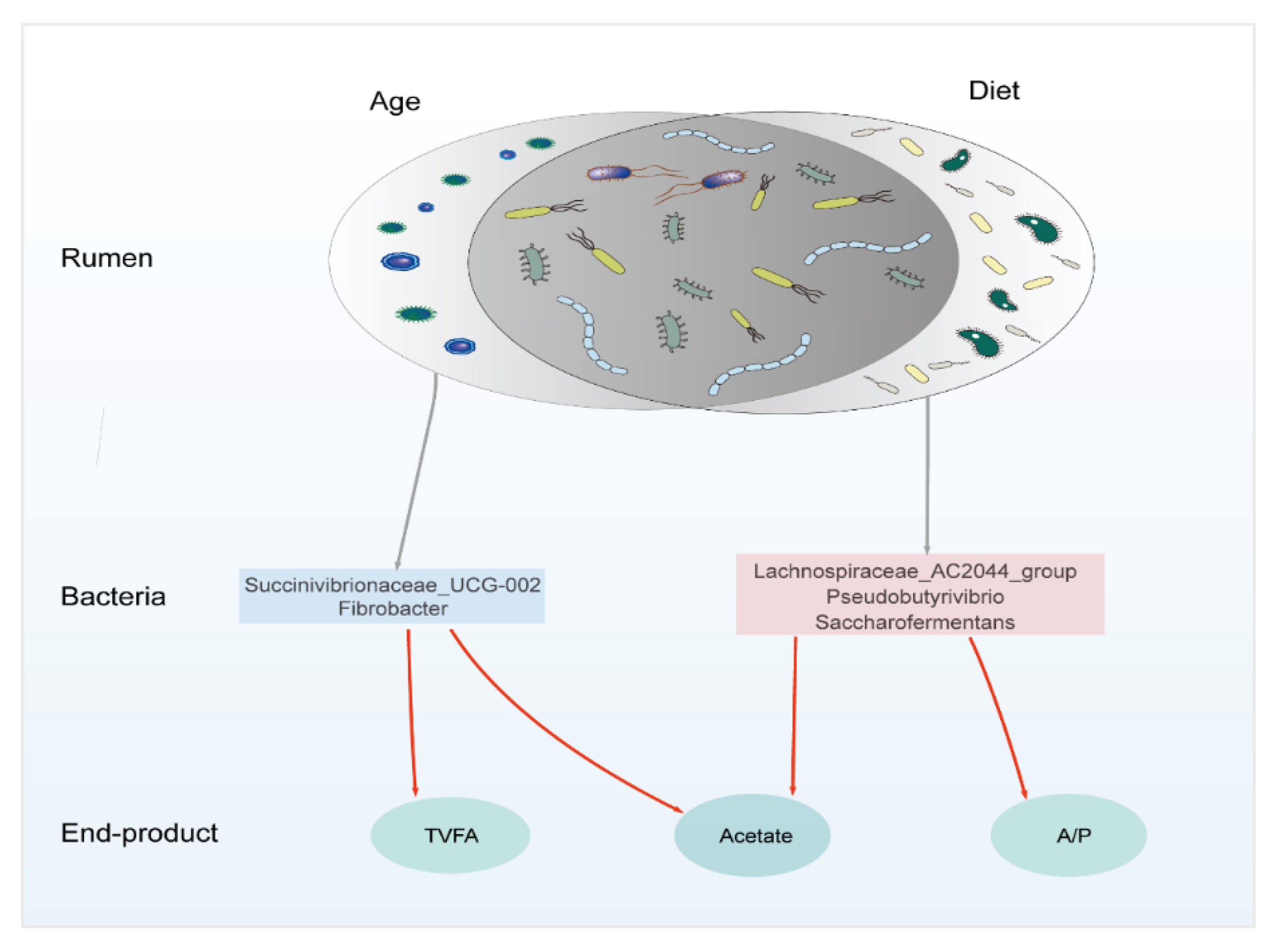
3.3. Driving Factors and the Correlations between Rumen Bacteria and Its Byproducts
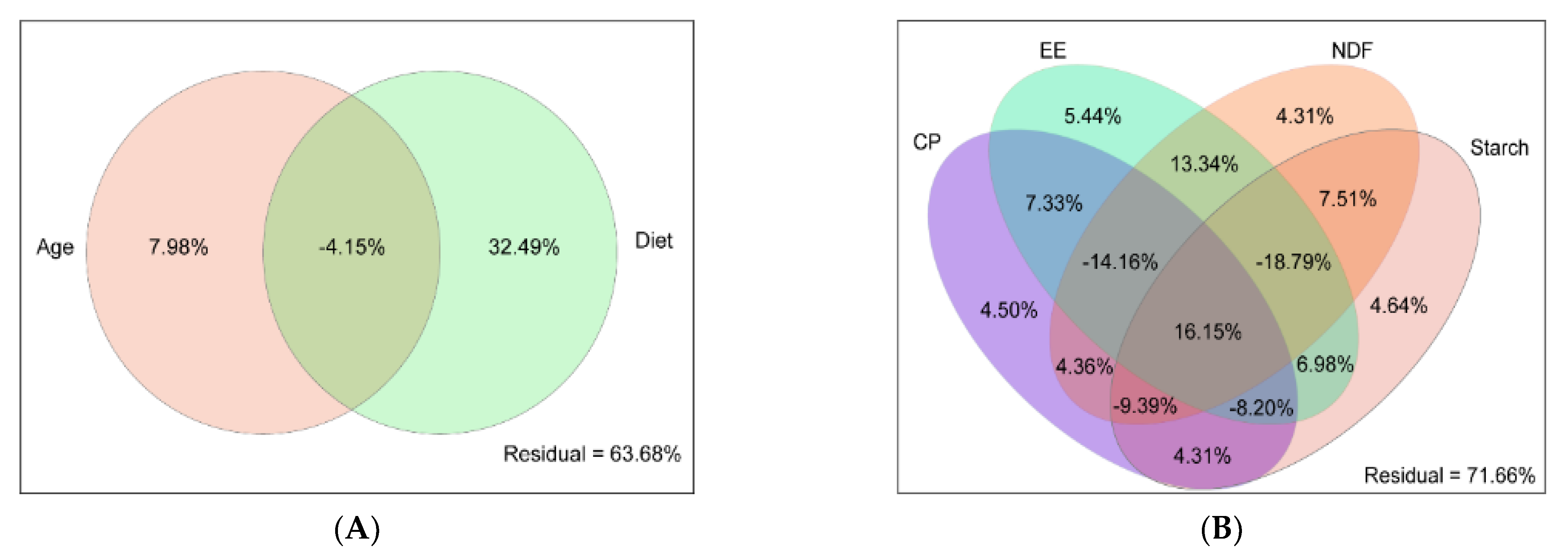
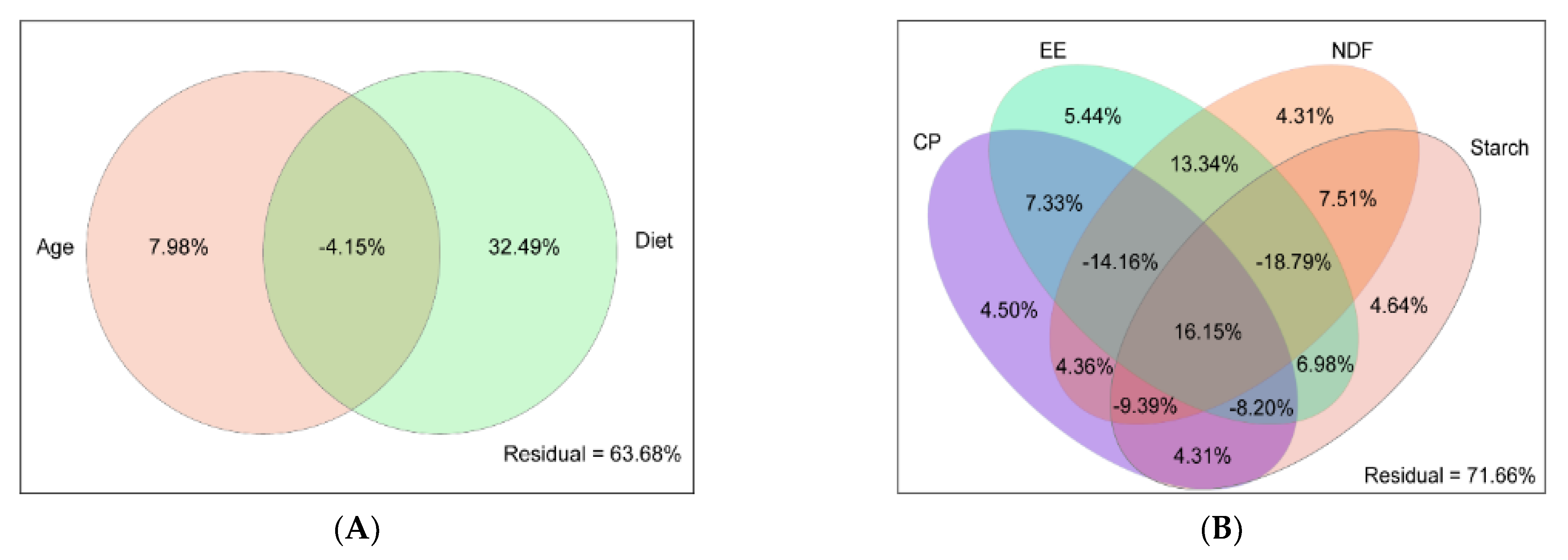
3.3.1. Driving Factors of Rumen Bacteria Variation
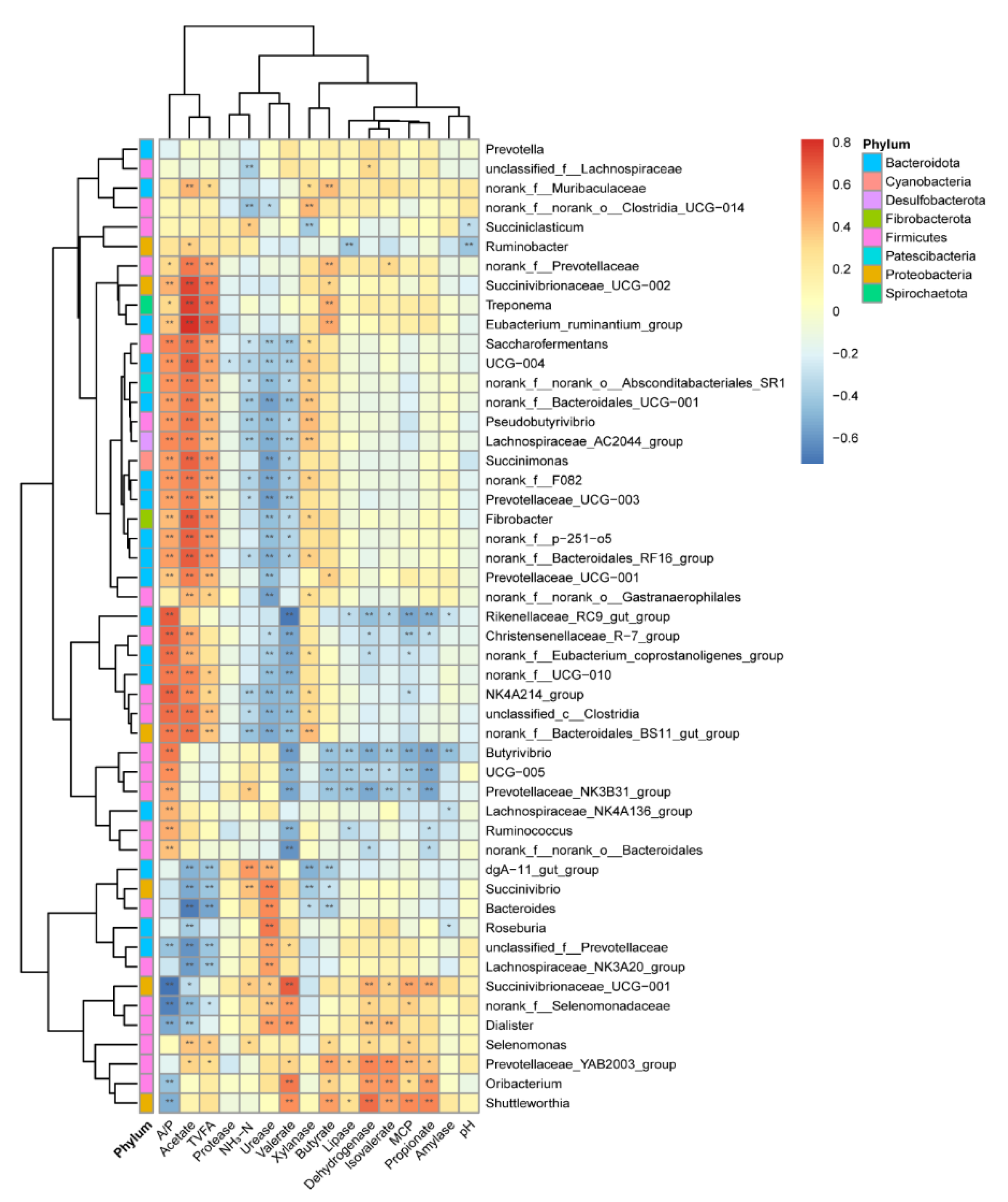
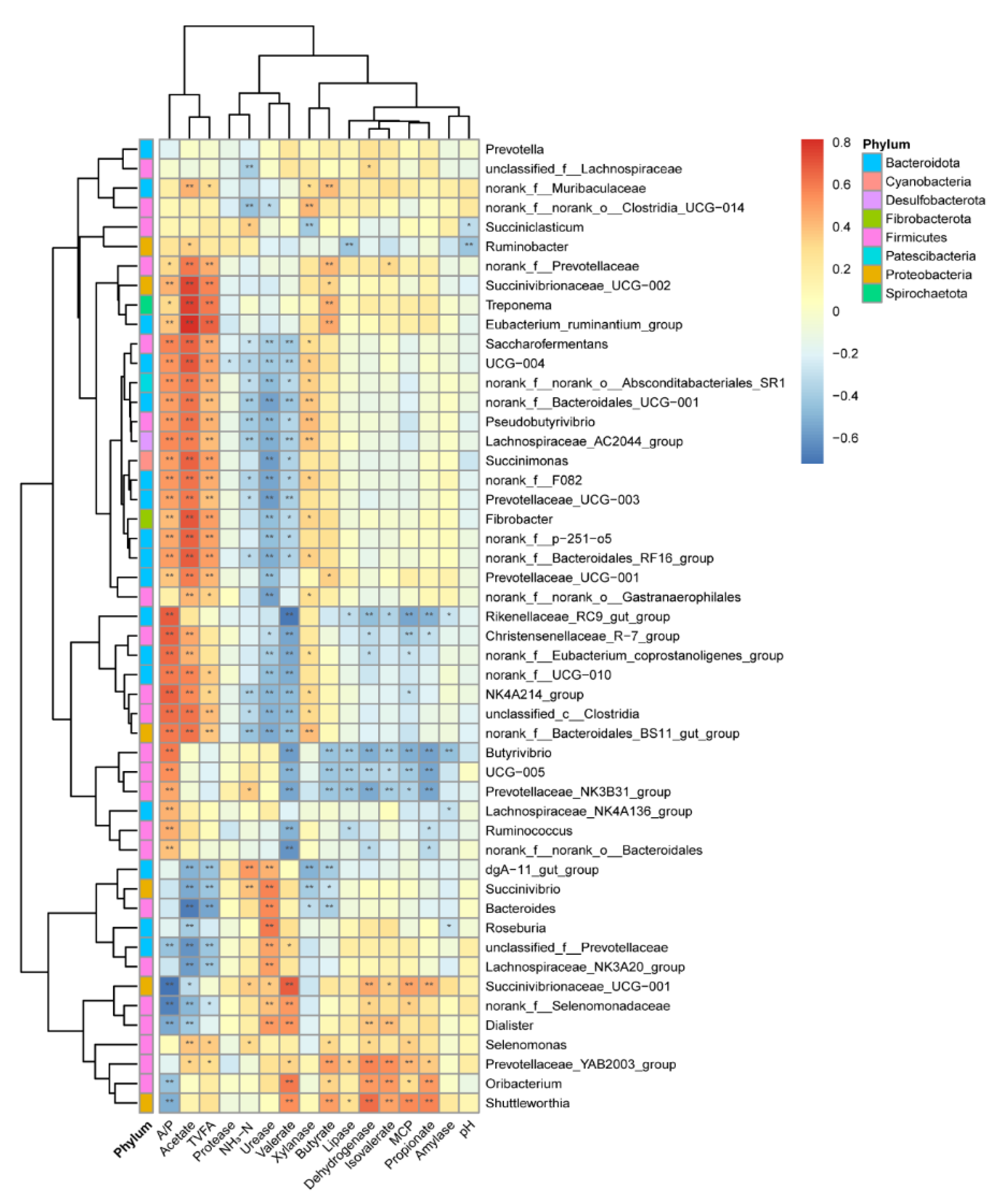
3.3.2. The Correlation between Bacteria and Its Main Byproducts
4. Discussion
4.1. Rumen Fermentation Profile and Enzyme Activity
4.2. Rumen Bacteria Composition
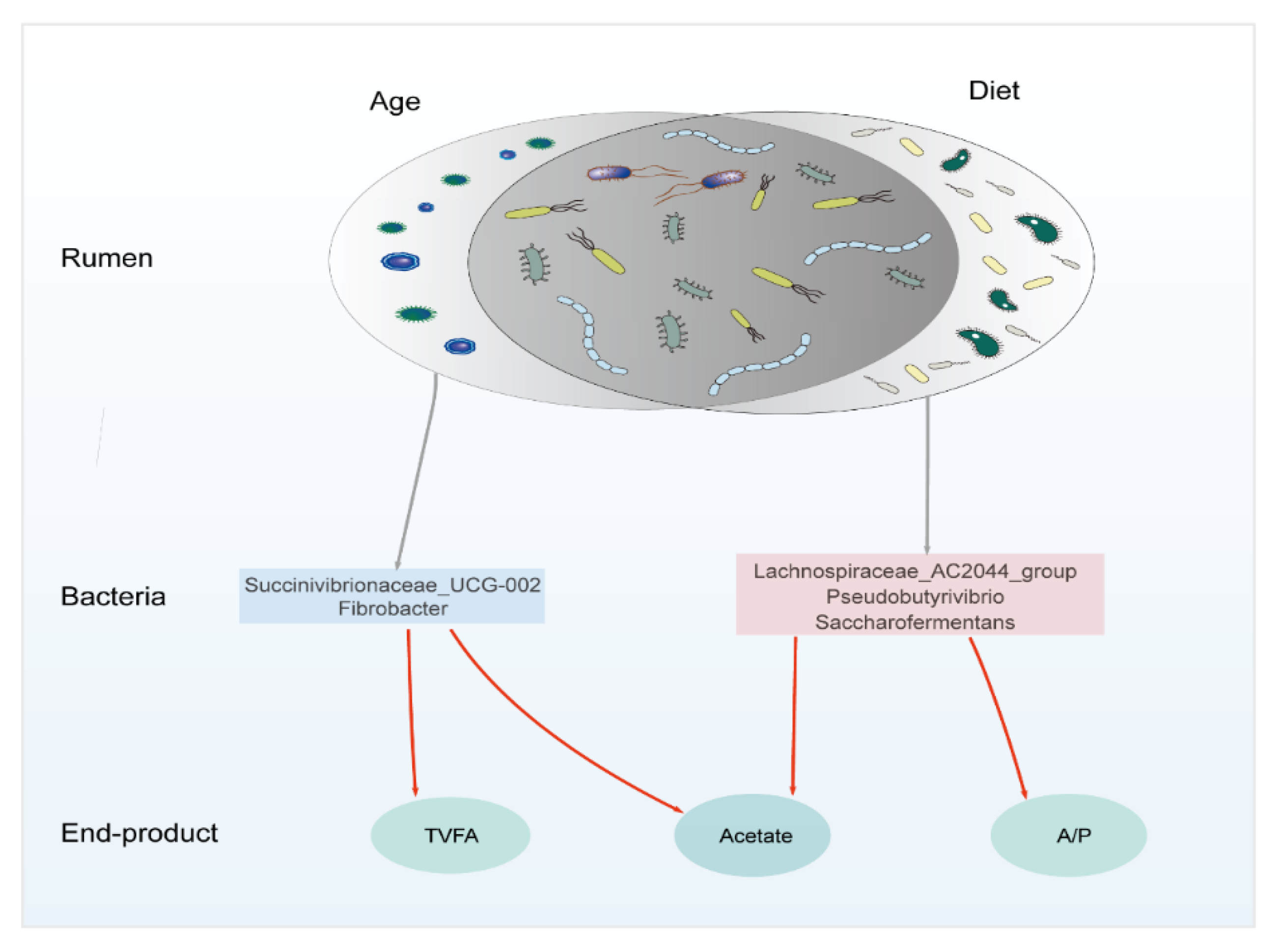
4.3. The Relationship within the Rumen Bacteria, Enzyme, and VFA
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Stewart, R.D.; Auffret, M.D.; Warr, A.; Wiser, A.H.; Press, M.O.; Langford, K.W.; Liachko, I.; Snelling, T.J.; Dewhurst, R.J.; Walker, A.W.; et al. Assembly of 913 microbial genomes from metagenomic sequencing of the cow rumen. Nat Commun. 2018, 9, 870. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Choudhury, P.K.; Salem, A.Z.M.; Jena, R.; Kumar, S.; Singh, R.; Puniya, A.K. Rumen microbiology: An overview. In Rumen Microbiology: From Evolution to Revolution; Puniya, A.K., Singh, R., Kamra, D.N., Eds.; Springer: New Delhi, India, 2015; pp. 3–16. [Google Scholar] [CrossRef]
- Yanez-Ruiz, D.R.; Abecia, L.; Newbold, C.J. Manipulating rumen microbiome and fermentation through interventions during early life: A review. Front. Microbiol. 2015, 6, 1133. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zilber-Rosenberg, I.; Rosenberg, E. Role of microorganisms in the evolution of animals and plants: The hologenome theory of evolution. FEMS Microbiol. Rev. 2008, 32, 723–735. [Google Scholar] [CrossRef]
- Stewart, R.D.; Auffret, M.D.; Warr, A.; Walker, A.W.; Roehe, R.; Watson, M. Compendium of 4,941 rumen metagenome-assembled genomes for rumen microbiome biology and enzyme discovery. Nat. Biotechnol. 2019, 37, 953–961. [Google Scholar] [CrossRef] [Green Version]
- Hess, M.; Sczyrba, A.; Egan, R.; Kim, T.-W.; Chokhawala, H.; Schroth, G.; Luo, S.; Clark, D.S.; Chen, F.; Zhang, T.; et al. Metagenomic discovery of biomass-degrading genes and genomes from cow rumen. Science 2011, 331, 463–467. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Noziere, P.; Steinberg, W.; Silberberg, M.; Morgavi, D.P. Amylase addition increases starch ruminal digestion in first-lactation cows fed high and low starch diets. J. Dairy Sci. 2014, 97, 2319–2328. [Google Scholar] [CrossRef]
- Der Bedrosian, M.C.; Kung, L., Jr. The effect of various doses of an exogenous acid protease on the fermentation and nutritive value of corn silage. J. Dairy Sci. 2019, 102, 10925–10933. [Google Scholar] [CrossRef]
- Fonty, G.; Gouet, P.; Ratefiarivelo, H.; Jouany, J.P. Establishment of Bacteroides succinogenes and measurement of the main digestive parameters in the rumen of gnotoxenic lambs. Can. J. Microbiol. 1988, 34, 938–946. [Google Scholar] [CrossRef]
- Fonty, G.; Gouet, P.; Jouany, J.P.; Senaud, J. Establishment of the microflora and anaerobic fungi in the rumen of lambs. Microbiology 1987, 133, 1835–1843. [Google Scholar] [CrossRef] [Green Version]
- Jami, E.; Israel, A.; Kotser, A.; Mizrahi, I. Exploring the bovine rumen bacterial community from birth to adulthood. ISME J. 2013, 7, 1069–1079. [Google Scholar] [CrossRef] [Green Version]
- Bohra, V.; Dafale, N.A.; Purohit, H.J. Understanding the alteration in rumen microbiome and CAZymes profile with diet and host through comparative metagenomic approach. Arch. Microbiol. 2019, 201, 1385–1397. [Google Scholar] [CrossRef] [PubMed]
- Gruninger, R.J.; Ribeiro, G.O.; Cameron, A.; McAllister, T.A. Invited review: Application of meta-omics to understand the dynamic nature of the rumen microbiome and how it responds to diet in ruminants. Animal 2019, 13, 1843–1854. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kumar, S.; Indugu, N.; Vecchiarelli, B.; Pitta, D.W. Associative patterns among anaerobic fungi, methanogenic archaea, and bacterial communities in response to changes in diet and age in the rumen of dairy cows. Front. Microbiol. 2015, 6, 781. [Google Scholar] [CrossRef] [Green Version]
- Rey, M.; Enjalbert, F.; Combes, S.; Cauquil, L.; Bouchez, O.; Monteils, V. Establishment of ruminal bacterial community in dairy calves from birth to weaning is sequential. J. Appl. Microbiol. 2014, 116, 245–257. [Google Scholar] [CrossRef] [PubMed]
- Anderson, K.L.; Nagaraja, T.G.; Morrill, J.L.; Avery, T.B.; Galitzer, S.J.; Boyer, J.E. Ruminal microbial development in conventionally or early-weaned calves. J. Anim. Sci. 1987, 64, 1215–1226. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gleason, C.B.; White, R.R. Variation in animal performance explained by the rumen microbiome or by diet composition. J. Anim. Sci. 2018, 96, 4658–4673. [Google Scholar] [CrossRef]
- Mu, Y.; Lin, X.; Wang, Z.; Hou, Q.; Wang, Y.; Hu, Z. High-production dairy cattle exhibit different rumen and fecal bacterial community and rumen metabolite profile than low-production cattle. Microbiologyopen 2019, 8, e00673. [Google Scholar] [CrossRef] [PubMed]
- Broderick, G.A.; Kang, J.H. Automated simultaneous determination of ammonia and total amino acids in ruminal fluid and in vitro media 1. J. Dairy Sci. 1980, 63, 64–75. [Google Scholar] [CrossRef]
- Makkar, H.P.; Sharma, O.P.; Dawra, R.K.; Negi, S.S. Simple determination of microbial protein in rumen. J. Dairy Sci. 1982, 65, 2170–2173. [Google Scholar] [CrossRef]
- Cao, Z.J.; Li, S.L.; Xing, J.J.; Ma, M.; Wang, L.L. Effects of maize grain and lucerne particle size on ruminal fermentation, digestibility and performance of cows in midlactation. J. Anim. Physiol. Anim. Nutr. 2008, 92, 157–167. [Google Scholar] [CrossRef]
- Weatherburn, M.W. Phenol-Hypochlorite Reaction for Determination of Ammonia. Anal. Chem. 1967, 36, 971–974. [Google Scholar] [CrossRef]
- Anson, M.L. The estimation of pepsin, trypsin, papain, and cathepsin with hemoglobin. J. Gen. Physiol. 1938, 22, 79–89. [Google Scholar] [CrossRef]
- Bernfield, P. Amylase, α and β Methods. Biochim. Biophys. Acta BBA Enzymol. 1955, 65, 149–158. [Google Scholar]
- Kanwar, S.S.; Kaushal, R.K.; Jawed, A.; Gupta, R.; Chimni, S.S. Methods for inhibition of residual lipase activity in colorimetric assay: A comparative study. Indian J. Biochem. Biophys. 2005, 42, 233. [Google Scholar]
- Lenártová, V.; Holovská, K.; Javorský, P.; Rybosová, E.; Havassy, I. Glutamate dehydrogenase and glutamine synthetase activity of the bacteria of the sheep’s rumen after different nitrogen intake. Physiol. Plant. 1987, 5, 471–476. [Google Scholar]
- Guo, M.; Wu, F.; Hao, G.; Qi, Q.; Li, R.; Li, N.; Wei, L.; Chai, T. Bacillus subtilis improves immunity and disease resistance in rabbits. Front. Immunol. 2017, 8, 354. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Magoc, T.; Salzberg, S.L. FLASH: Fast length adjustment of short reads to improve genome assemblies. Bioinformatics 2011, 27, 2957–2963. [Google Scholar] [CrossRef] [PubMed]
- Bokulich, N.A.; Subramanian, S.; Faith, J.J.; Gevers, D.; Gordon, J.I.; Knight, R.; Mills, D.A.; Caporaso, J.G. Quality-filtering vastly improves diversity estimates from Illumina amplicon sequencing. Nat. Methods 2012, 10, 57–59. [Google Scholar] [CrossRef]
- Caporaso, J.G.; Kuczynski, J.; Stombaugh, J.; Bittinger, K.; Bushman, F.D.; Costello, E.K.; Fierer, N.; Peña, A.G.; Goodrich, J.K.; Gordon, J.I.; et al. QIIME allows analysis of high-throughput community sequencing data. Nat. Methods 2010, 7, 335–336. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Edgar, R.C.; Haas, B.J.; Clemente, J.C.; Quince, C.; Knight, R. UCHIME improves sensitivity and speed of chimera detection. Bioinformatics 2011, 27, 2194–2200. [Google Scholar] [CrossRef] [Green Version]
- Haas, B.J.; Gevers, D.; Earl, A.M.; Feldgarden, M.; Ward, D.V.; Giannoukos, G.; Ciulla, D.; Tabbaa, D.; Highlander, S.K.; Sodergren, E.; et al. Chimeric 16S rRNA sequence formation and detection in Sanger and 454-pyrosequenced PCR amplicons. Genome Res. 2011, 21, 494–504. [Google Scholar] [CrossRef] [Green Version]
- Edgar, R.C. UPARSE: Highly accurate OTU sequences from microbial amplicon reads. Nat. Methods 2013, 10, 996–998. [Google Scholar] [CrossRef]
- Quast, C.; Pruesse, E.; Yilmaz, P.; Gerken, J.; Schweer, T.; Yarza, P.; Peplies, J.; Glockner, F.O. The SILVA ribosomal RNA gene database project: Improved data processing and web-based tools. Nucleic Acids Res. 2013, 41, D590–D596. [Google Scholar] [CrossRef]
- Benjamini, Y.; Hochberg, Y. Controlling the False Discovery Rate: A Practical and Powerful Approach to Multiple Testing. J. R. Stat. Soc. Ser. A Stat. Soc. 1995, 57, 289–300. [Google Scholar] [CrossRef]
- Segata, N.; Izard, J.; Waldron, L.; Gevers, D.; Miropolsky, L.; Garrett, W.S.; Huttenhower, C. Metagenomic biomarker discovery and explanation. Genome Biol. 2011, 12, R60. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, C.J.; Chen, Y.L.; Pan, J.; Wang, Y.M.; Li, M. Spatial and seasonal variation of methanogenic community in a river-bay system in South China. Appl. Microbiol. Biotechnol. 2020, 104, 4593–4603. [Google Scholar] [CrossRef]
- Chopyk, J.; Akrami, K.; Bavly, T.; Shin, J.H.; Schwanemann, L.K.; Ly, M.; Kalia, R.; Xu, Y.; Kelley, S.T.; Malhotra, A.; et al. Temporal variations in bacterial community diversity and composition throughout intensive care unit renovations. Microbiome 2020, 8, 86. [Google Scholar] [CrossRef]
- Shi, W.; Haisan, J.; Inabu, Y.; Sugino, T.; Oba, M. Effects of starch concentration of close-up diets on rumen pH and plasma metabolite responses of dairy cows to grain challenges after calving. J. Dairy Sci. 2020, 103, 11461–11471. [Google Scholar] [CrossRef] [PubMed]
- Shen, H.; Lu, Z.; Xu, Z.; Chen, Z.; Shen, Z.J.M. Associations among dietary non-fiber carbohydrate, ruminal microbiota and epithelium G-protein-coupled receptor, and histone deacetylase regulations in goats. Microbiome 2017, 5, 123. [Google Scholar] [CrossRef] [Green Version]
- Dias, A.L.G.; Freitas, J.A.; Micai, B.; Azevedo, R.A.; Greco, L.F.; Santos, J.E.P. Effect of supplemental yeast culture and dietary starch content on rumen fermentation and digestion in dairy cows. J. Dairy Sci. 2018, 101, 201–221. [Google Scholar] [CrossRef] [PubMed]
- Gasiorek, M.; Stefanska, B.; Pruszynska-Oszmalek, E.; Taciak, M.; Komisarek, J.; Nowak, W. Effect of oat hay provision method on growth performance, rumen fermentation and blood metabolites of dairy calves during preweaning and postweaning periods. Animal 2020, 14, 2054–2062. [Google Scholar] [CrossRef]
- Jiang, F.G.; Lin, X.Y.; Yan, Z.G.; Hu, Z.Y.; Liu, G.M.; Sun, Y.D.; Liu, X.W.; Wang, Z.H. Effect of dietary roughage level on chewing activity, ruminal pH, and saliva secretion in lactating Holstein cows. J. Dairy Sci. 2017, 100, 2660–2671. [Google Scholar] [CrossRef] [Green Version]
- Liu, C.; Meng, Q.; Chen, Y.; Xu, M.; Shen, M.; Gao, R.; Gan, S. Role of Age-Related Shifts in Rumen Bacteria and Methanogens in Methane Production in Cattle. Front. Microbiol. 2017, 8, 1563. [Google Scholar] [CrossRef] [Green Version]
- Patra, A.K.; Aschenbach, J.R. Ureases in the gastrointestinal tracts of ruminant and monogastric animals and their implication in urea-N/ammonia metabolism: A review. J. Adv. Res. 2018, 13, 39–50. [Google Scholar] [CrossRef]
- Jin, D.; Zhao, S.; Zheng, N.; Beckers, Y.; Wang, J. Urea Metabolism and Regulation by Rumen Bacterial Urease in Ruminants—A Review. Ann. Anim. Sci. 2018, 18, 303–318. [Google Scholar] [CrossRef] [Green Version]
- Baldwin, R.L.t.; Connor, E.E. Rumen function and development. Vet. Clin. N. Am. Food Anim. Pract. 2017, 33, 427–439. [Google Scholar] [CrossRef] [PubMed]
- Lu, Z.; Xu, Z.; Shen, Z.; Tian, Y.; Shen, H. Dietary Energy Level Promotes Rumen Microbial Protein Synthesis by Improving the Energy Productivity of the Ruminal Microbiome. Front. Microbiol. 2019, 10, 847. [Google Scholar] [CrossRef] [PubMed]
- Zhu, Z.; Kristensen, L.; Difford, G.F.; Poulsen, M.; Noel, S.J.; Abu Al-Soud, W.; Sorensen, S.J.; Lassen, J.; Lovendahl, P.; Hojberg, O. Changes in rumen bacterial and archaeal communities over the transition period in primiparous Holstein dairy cows. J. Dairy Sci. 2018, 101, 9847–9862. [Google Scholar] [CrossRef]
- Rey, M.; Enjalbert, F.; Monteils, V. Establishment of ruminal enzyme activities and fermentation capacity in dairy calves from birth through weaning. J. Dairy Sci. 2012, 95, 1500–1512. [Google Scholar] [CrossRef]
- White, B.A.; Lamed, R.; Bayer, E.A.; Flint, H.J. Biomass utilization by gut microbiomes. Annu. Rev. Microbiol. 2014, 68, 279–296. [Google Scholar] [CrossRef] [PubMed]
- Koike, S.; Handa, Y.; Goto, H.; Sakai, K.; Miyagawa, E.; Matsui, H.; Ito, S.; Kobayashi, Y. Molecular monitoring and isolation of previously uncultured bacterial strains from the sheep rumen. Appl. Environ. Microbiol. 2010, 76, 1887–1894. [Google Scholar] [CrossRef] [Green Version]
- Burrell, P.C.; O’Sullivan, C.; Song, H.; Clarke, W.P.; Blackall, L.L. Identification, Detection, and Spatial Resolution of Clostridium Populations Responsible for Cellulose Degradation in a Methanogenic Landfill Leachate Bioreactor. Appl. Environ. Microbiol. 2004, 70, 2414–2419. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Huws, S.A.; Edwards, J.E.; Creevey, C.J.; Rees Stevens, P.; Lin, W.; Girdwood, S.E.; Pachebat, J.A.; Kingston-Smith, A.H. Temporal dynamics of the metabolically active rumen bacteria colonizing fresh perennial ryegrass. FEMS Microbiol. Ecol. 2015, 92, fiv137. [Google Scholar] [CrossRef] [Green Version]
- Liu, J.; Zhang, M.; Xue, C.; Zhu, W.; Mao, S. Characterization and comparison of the temporal dynamics of ruminal bacterial microbiota colonizing rice straw and alfalfa hay within ruminants. J. Dairy Sci. 2016, 99, 9668–9681. [Google Scholar] [CrossRef] [Green Version]
- Mao, S.Y.; Huo, W.J.; Zhu, W.Y. Microbiome-metabolome analysis reveals unhealthy alterations in the composition and metabolism of ruminal microbiota with increasing dietary grain in a goat model. Environ. Microbiol. 2016, 18, 525–541. [Google Scholar] [CrossRef] [PubMed]
- Granja-Salcedo, Y.T.; Fernandes, R.M.; de Araujo, R.C.; Kishi, L.T.; Berchielli, T.T.; de Resende, F.D.; Berndt, A.; Siqueira, G.R. Long-term encapsulated nitrate supplementation modulates rumen microbial diversity and rumen fermentation to reduce methane emission in grazing steers. Front. Microbiol. 2019, 10, 614. [Google Scholar] [CrossRef] [PubMed]
- Hailemariam, S.; Zhao, S.; Wang, J. Complete genome sequencing and transcriptome analysis of nitrogen metabolism of succinivibrio dextrinosolvens strain Z6 isolated from dairy cow rumen. Front. Microbiol. 2020, 11, 1826. [Google Scholar] [CrossRef]
- Huws, S.A.; Creevey, C.J.; Oyama, L.B.; Mizrahi, I.; Denman, S.E.; Popova, M.; Munoz-Tamayo, R.; Forano, E.; Waters, S.M.; Hess, M.; et al. Addressing global ruminant agricultural challenges through understanding the rumen microbiome: Past, present, and future. Front. Microbiol. 2018, 9, 2161. [Google Scholar] [CrossRef]
- Neumann, A.P.; McCormick, C.A.; Suen, G. Fibrobacter communities in the gastrointestinal tracts of diverse hindgut-fermenting herbivores are distinct from those of the rumen. Environ. Microbiol. 2017, 19, 3768–3783. [Google Scholar] [CrossRef]
- Palevich, N.; Maclean, P.H.; Kelly, W.J.; Leahy, S.C.; Rakonjac, J.; Attwood, G.T. Complete genome sequence of the polysaccharide-degrading rumen bacterium pseudobutyrivibrio xylanivorans MA3014 reveals an incomplete glycolytic pathway. Genome Biol. Evol. 2020, 12, 1566–1572. [Google Scholar] [CrossRef]
- Tomazetto, G.; Hahnke, S.; Wibberg, D.; Puhler, A.; Klocke, M.; Schluter, A. Proteiniphilum saccharofermentans str. M3/6(T) isolated from a laboratory biogas reactor is versatile in polysaccharide and oligopeptide utilization as deduced from genome-based metabolic reconstructions. Biotechnol. Rep. 2018, 18, e00254. [Google Scholar] [CrossRef] [PubMed]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Items | Groups | SEM | p-Value | |||||
|---|---|---|---|---|---|---|---|---|
| M1.5 | M6 | M9 | M18 | M23 | M27 | |||
| pH | 6.52 ab | 6.69 a | 6.50 ab | 6.53 ab | 6.36 bc | 6.25 c | 0.04 | <0.01 |
| NH3-N (mg/dL) | 32.98 a | 11.23 de | 13.61 cd | 8.97 e | 16.41 c | 20.35 b | 1.22 | <0.01 |
| MCP (µg/mL) | 81.05 b | 116.01 a | 79.14 b | 61.62 c | 119.29 a | 127.07 a | 3.96 | <0.01 |
| Acetate (mmol/mL) | 31.91 c | 47.34 b | 69.16 ab | 75.34 a | 73.58 ab | 64.55 ab | 2.45 | <0.01 |
| Propionate (mmol/mL) | 13.73 d | 24.75 a | 18.84 bcd | 16.21 cd | 20.23 abc | 23.24 ab | 0.89 | <0.01 |
| Butyrate (mmol/mL) | 2.50 d | 7.48 ab | 6.10 bc | 5.04 c | 6.46 abc | 7.76 a | 0.32 | <0.01 |
| TVFA (mmol/mL) | 52.63 c | 80.59 b | 61.48 c | 82.69 c | 101.47 a | 102.33 a | 3.31 | <0.01 |
| A/P | 3.04 b | 1.95 c | 3.70 a | 3.73 a | 3.70 a | 3.01 b | 0.12 | <0.01 |
| Items | Groups | SEM | p-Value | |||||
|---|---|---|---|---|---|---|---|---|
| M1.5 | M6 | M9 | M18 | M23 | M27 | |||
| Dehydrogenase (µg/min/mL) | 0.49 c | 0.87 a | 0.79 ab | 0.64 bc | 0.63 bc | 0.77 ab | 0.03 | <0.01 |
| Urease (µg/min/mL) | 2.63 a | 2.15 abc | 2.31 ab | 1.36 d | 1.68 cd | 1.85 bcd | 0.09 | <0.01 |
| Protease (µg/min/mL) | 14.81 a | 10.12 ab | 9.34 b | 12.27 ab | 10.54 ab | 13.84 ab | 0.69 | <0.01 |
| Xylanase (nmol/min/mL) | 131.68 c | 183.26 bc | 250.43 ab | 311.81 a | 214.75 b | 221.89 b | 12.20 | <0.01 |
| Amylase (mg/min/mL) | 0.57 cd | 1.10 ab | 0.31 d | 0.85 bc | 0.48 bc | 1.47 a | 0.17 | <0.01 |
| Lipase (nmol/min/mL) | 101.49 c | 145.07 a | 117.48 bc | 113.65 bc | 114.30 bc | 127.25 ab | 3.15 | <0.01 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hao, Y.; Gong, Y.; Huang, S.; Ji, S.; Wang, W.; Wang, Y.; Yang, H.; Cao, Z.; Li, S. Effects of Age, Diet CP, NDF, EE, and Starch on the Rumen Bacteria Community and Function in Dairy Cattle. Microorganisms 2021, 9, 1788. https://doi.org/10.3390/microorganisms9081788
Hao Y, Gong Y, Huang S, Ji S, Wang W, Wang Y, Yang H, Cao Z, Li S. Effects of Age, Diet CP, NDF, EE, and Starch on the Rumen Bacteria Community and Function in Dairy Cattle. Microorganisms. 2021; 9(8):1788. https://doi.org/10.3390/microorganisms9081788
Chicago/Turabian StyleHao, Yangyi, Yue Gong, Shuai Huang, Shoukun Ji, Wei Wang, Yajing Wang, Hongjian Yang, Zhijun Cao, and Shengli Li. 2021. "Effects of Age, Diet CP, NDF, EE, and Starch on the Rumen Bacteria Community and Function in Dairy Cattle" Microorganisms 9, no. 8: 1788. https://doi.org/10.3390/microorganisms9081788
APA StyleHao, Y., Gong, Y., Huang, S., Ji, S., Wang, W., Wang, Y., Yang, H., Cao, Z., & Li, S. (2021). Effects of Age, Diet CP, NDF, EE, and Starch on the Rumen Bacteria Community and Function in Dairy Cattle. Microorganisms, 9(8), 1788. https://doi.org/10.3390/microorganisms9081788