
Indirect Selection against Antibiotic Resistance via Specialized Plasmid-Dependent Bacteriophages


Abstract
1. Introduction
2. Plasmid-Dependent Bacteriophages
3. How Does Plasmid-Dependent Phages Affect Resistant Bacteria?
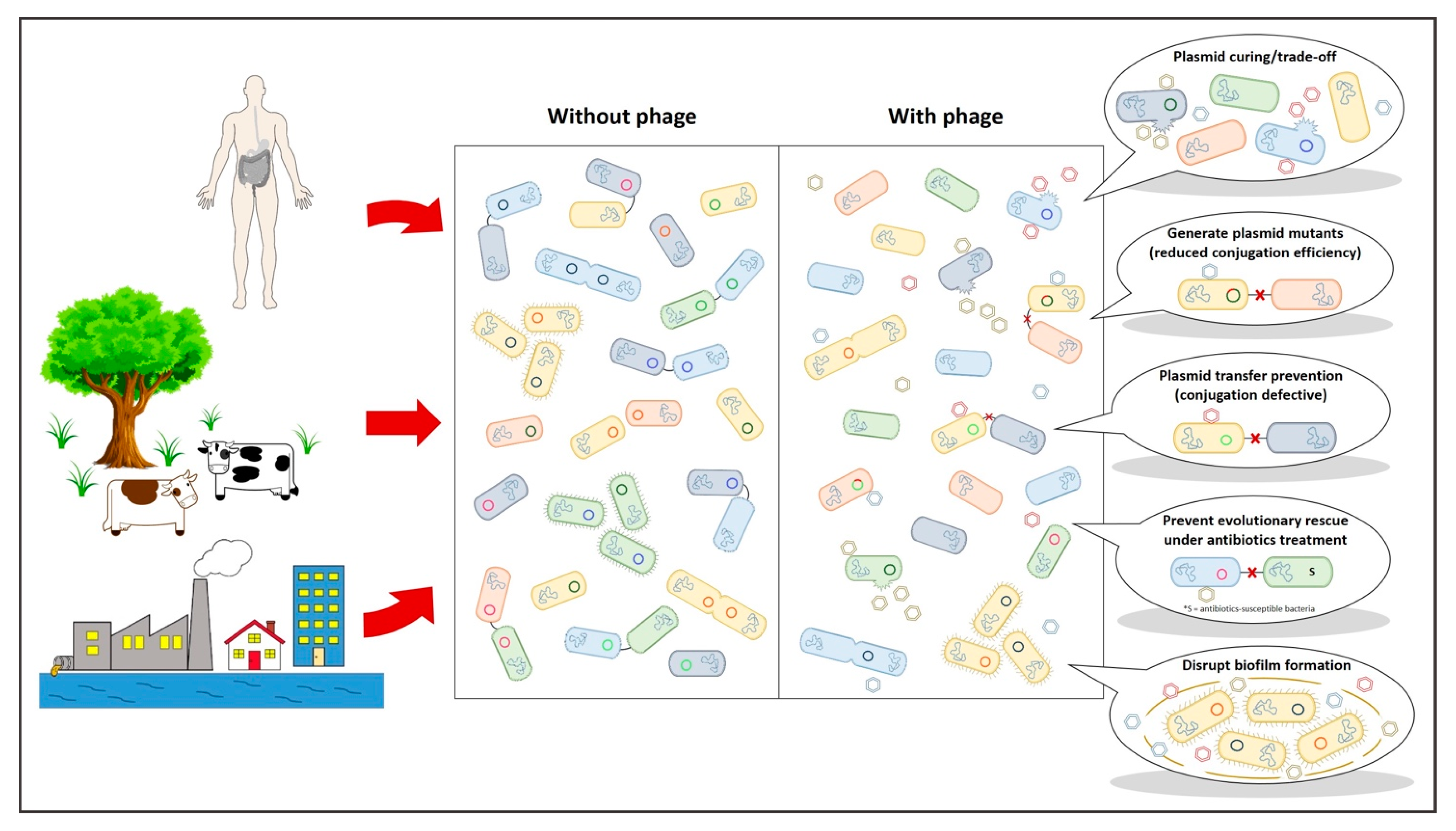
4. Dam the Flow: Why Disrupting Genetic Exchange Can Make a Difference?
5. Concluding Remarks
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Nathan, C. Resisting antimicrobial resistance. Nat. Rev. Microbiol. 2020, 18, 259–260. [Google Scholar] [CrossRef] [PubMed]
- Cairns, J.; Frickel, J.; Jalasvuori, M.; Hiltunen, T.; Becks, L. Genomic evolution of bacterial populations under co-selection by antibiotics and phage. Mol. Ecol. 2017, 26, 1848–1859. [Google Scholar] [CrossRef] [PubMed]
- Cairns, J.; Koskinen, K.; Penttinen, R.; Patinen, T.; Hartikainen, A.; Jokela, R.; Ruusulehto, L.; Viitamäki, S.; Mattila, S.; Hiltunen, T.; et al. Black Queen evolution and trophic interactions determine plasmid survival after the disruption of conjugation network. mSystems 2018, 3, e00104-18. [Google Scholar] [CrossRef]
- Chong, Y.; Shimoda, S.; Shimono, N. Current epidemiology, genetic evolution and clinical impact of extended-spectrum β-lactamase-producing Escherichia coli and Klebsiella pneumoniae. Infect. Genet. Evol. 2018, 61, 185–188. [Google Scholar] [CrossRef] [PubMed]
- Ruotsalainen, P.; Given, C.; Penttinen, R.; Jalasvuori, M. Beta-Lactam Sensitive Bacteria Can Acquire ESBL-Resistance via Conjugation after Long-Term Exposure to Lethal Antibiotic Concentration. Antibiotics 2020, 9, 296. [Google Scholar]
- Tooke, C.L.; Hinchliffe, P.; Bragginton, E.C.; Colenso, C.K.; Hirvonen, V.H.; Takebayashi, Y.; Spencer, J. β-Lactamases and β-Lactamase Inhibitors in the 21st Century. J. Mol. Biol. 2019, 431, 3472–3500. [Google Scholar] [CrossRef]
- Carattoli, A. Resistance Plasmid Families in Enterobacteriaceae. Antimicrob. Agents Chemother. 2009, 53, 2227–2238. [Google Scholar] [CrossRef]
- Jalasvuori, M.; Koonin, E. Classification of Prokaryotic Genetic Replicators: Between Selfishness and Altruism. Ann. N. Y. Acad. Sci. 2015, 1341, 96–105. [Google Scholar]
- Dahlberg, C.; Chao, L. Amelioration of the cost of conjugative plasmid carriage in Eschericha coli K12. Genetics 2003, 165, 1641–1649. [Google Scholar]
- Jalasvuori, M. Vehicles, replicators, and intercellular movement of genetic information: Evolutionary dissection of a bacterial cell. Int. J. Evol. Biol. 2012, 2012, 874153. [Google Scholar] [CrossRef]
- Lopatkin, A.J.; Meredith, H.R.; Srimani, J.K.; Pfeiffer, C.; Durrett, R.; You, L. Persistence and reversal of plasmid-mediated antibiotic resistance. Nat. Commun. 2017, 8, 1689. [Google Scholar] [CrossRef] [PubMed]
- Buckner, M.M.C.; Ciusa, M.L.; Piddock, L.J.V. Strategies to combat antimicrobial resistance: Anti-plasmid and plasmid curing. FEMS Microbiol. Rev. 2018, 42, 781–804. [Google Scholar] [CrossRef] [PubMed]
- Getino, M.; de la Cruz, F. Natural and Artificial Strategies to Control the Conjugative Transmission of Plasmids. Microbiol. Spectr. 2018, 6, 1. [Google Scholar]
- Bikard, D.; Euler, C.W.; Jiang, W.; Nussenzweig, P.M.; Goldberg, G.W.; Duportet, X.; Fischetti, V.A.; Marraffini, L.A. Exploiting CRISPR-cas nucleases to produce sequence-specific antimicrobials. Nat. Biotechnol. 2014, 32, 1146–1150. [Google Scholar] [CrossRef] [PubMed]
- Citorik, R.J.; Mimee, M.; Lu, T.K. Sequence-specific antimicrobials using efficiently delivered RNA-guided nucleases. Nat. Biotechnol. 2014, 32, 1141–1145. [Google Scholar] [CrossRef] [PubMed]
- Ruotsalainen, P.; Penttinen, R.; Mattila, S.; Jalasvuori, M. Midbiotics: Conjugative plasmids for genetic engineering of natural gut flora. Gut Microbes 2019, 10, 643–653. [Google Scholar] [CrossRef]
- Ojala, V.; Mattila, S.; Hoikkala, V.; Bamford, J.K.H.; Hiltunen, T.; Jalasvuori, M. Scoping the effectiveness and evolutionary obstacles in utilizing plasmid-dependent phages to fight antibiotic resistance. Future Microbiol. 2016, 11, 999–1009. [Google Scholar]
- Benson, S.D.; Bamford, J.K.H.; Bamford, D.H.; Burnett, R.M. Viral evolution revealed by bacteriophage PRD1 and human adenovirus coat protein structures. Cell 1999, 98, 825–833. [Google Scholar] [CrossRef]
- Krupovic, M.; Bamford, D.H. Virus evolution: How far does the double beta-barrel viral lineage extend? Nat. Rev. Microbiol. 2008, 6, 941–948. [Google Scholar] [CrossRef]
- Olsen, R.H.; Siak, J.S.; Gray, R.H. Characteristics of PRD1, a plasmid-dependent broad host range DNA bacteriophage. J. Virol. 1974, 14, 689–699. [Google Scholar] [CrossRef]
- Abrescia, N.; Cockburn, J.; Grimes, J.; Sutton, G.C.; Diprose, J.M.; Butcher, S.J.; Fuller, S.D.; Martín, C.S.; Burnett, R.M.; Stuart, D.I.; et al. Insights into assembly from structural analysis of bacteriophage PRD1. Nature 2004, 432, 68–74. [Google Scholar] [CrossRef] [PubMed]
- Kotilainen, M.M.; Grahn, A.M.; Bamford, J.K.; Bamford, D.H. Binding of an Escherichia coli double-stranded DNA virus PRD1 to a receptor coded by an IncP-type plasmid. J. Bacteriol. 1993, 175, 3089–3095. [Google Scholar] [CrossRef] [PubMed]
- Rydman, P.S.; Bamford, D.H. Bacteriophage PRD1 DNA entry uses a viral membrane-associated transglycosylase activity. Mol. Microbiol. 2000, 37, 356–363. [Google Scholar] [CrossRef]
- Rydman, P.S.; Bamford, D.H. The lytic enzyme of bacteriophage PRD1 is associated with the viral membrane. J. Bacteriol. 2002, 184, 104–110. [Google Scholar] [CrossRef] [PubMed]
- Butcher, S.J.; Manole, V.; Karhu, N.J. Lipid-containing viruses: Bacteriophage PRD1 assembly. Adv. Exp. Med. Biol. 2012, 726, 365–377. [Google Scholar] [CrossRef] [PubMed]
- Žiedaitė, G.; Daugelavičius, R.; Bamford, J.K.H.; Bamford, D.H. The Holin Protein of Bacteriophage PRD1 Forms a Pore for Small-Molecule and Endolysin Translocation. J. Bacteriol. 2005, 187, 5397–5405. [Google Scholar] [CrossRef] [PubMed]
- Strömsten, N.J.; Bamford, D.H.; Bamford, J.K. In vitro DNA packaging of PRD1: A common mechanism for internal-membrane viruses. J. Mol. Biol. 2005, 348, 617–629. [Google Scholar] [CrossRef] [PubMed]
- Žiedaitė, G.; Kivelä, H.M.; Bamford, J.K.; Bamford, D.H. Purified membrane-containing procapsids of bacteriophage PRD1 package the viral genome. J. Mol. Biol. 2009, 386, 637–647. [Google Scholar] [CrossRef] [PubMed]
- Loeb, T. Isolation of a bacteriophage specific for the F plus and Hfr mating types of Escherichia coli K-12. Science 1960, 131, 932–933. [Google Scholar] [CrossRef]
- Caro, L.G.; Schnos, M. The attachment of the male-specific bacteriophage F1 to sensitive strains of Escherichia coli. Proc. Natl. Acad. Sci. USA 1966, 56, 126–132. [Google Scholar]
- Marvin, D.A.; Symmons, M.F.; Straus, S.K. Structure and assembly of filamentous bacteriophages. Prog. Biophys. Mol. Biol. 2014, 114, 80–122. [Google Scholar] [CrossRef] [PubMed]
- Rasched, I.; Oberer, E. Ff coliphages: Structural and functional relationships. Microbiol. Rev. 1986, 50, 401–427. [Google Scholar] [CrossRef] [PubMed]
- Jacobson, A. Role of F pili in the penetration of bacteriophage fl. J. Virol. 1972, 10, 835–843. [Google Scholar] [CrossRef]
- Russel, M.; Whirlow, H.; Sun, T.P.; Webster, R.E. Low-frequency infection of F- bacteria by transducing particles of filamentous bacteriophages. J. Bacteriol. 1988, 170, 5312–5316. [Google Scholar] [CrossRef]
- Sun, T.P.; Webster, R.E. fii, a bacterial locus required for filamentous phage infection and its relation to colicin-tolerant tolA and tolB. J. Bacteriol. 1986, 165, 107–115. [Google Scholar] [CrossRef] [PubMed]
- Smilowitz, H. Bacteriophage fl infection: Fate of theparental major coat protein. J. Virol. 1974, 13, 94–99. [Google Scholar] [CrossRef] [PubMed]
- Trenkner, E.; Bonhoeffer, F.; Gierer, A. The fate of the protein components of bacteriophage fd during infection. Biochem. Biophys. Res. Commun. 1967, 28, 932–939. [Google Scholar] [CrossRef]
- Hertveldt, K.; Beliën, T.; Volckaert, G. General M13 phage display: M13 phage display in identification and characterization of protein-protein interactions. Methods Mol. Biol. 2009, 502, 321–339. [Google Scholar]
- Ledsgaard, L.; Kilstrup, M.; Karatt-Vellatt, A.; McCafferty, J.; Laustsen, A.H. Basics of Antibody Phage Display Technology. Toxins 2018, 10, 236. [Google Scholar] [CrossRef]
- Friedman, S.D.; Genthner, F.J.; Gentry, J.; Sobsey, M.D.; Vinjé, J. Gene mapping and phylogenetic analysis of the complete genome from 30 single-stranded RNA male-specific coliphages (family Leviviridae). J. Virol. 2009, 83, 11233–11243. [Google Scholar] [CrossRef]
- Frost, L.S. Conjugative Pili and Pilus-Specific Phages. In Bacterial Conjugation; Clewell, D.B., Ed.; Springer: Boston, MA, USA, 1993. [Google Scholar] [CrossRef]
- Spiegelman, S.; Haruna, I.; Holland, I.B.; Beaudreau, G.; Mills, D. The Synthesis of a Self-propagating and Infectious Nucleic Acid with a Purified Enzyme. Proc. Natl. Acad. Sci. USA 1965, 54, 919–927. [Google Scholar] [CrossRef] [PubMed]
- Mills, D.R.; Peterson, R.L.; Spiegelman, S. An extracellular Darwinian experiment with a self-duplicating nucleic acid molecule. Proc. Natl. Acad. Sci. 1967, 58, 217–224. [Google Scholar] [CrossRef] [PubMed]
- Rozwandowicz, M.; Brouwer, M.S.M.; Fischer, J.; A Wagenaar, J.; Gonzalez-Zorn, B.; Guerra, B.; Mevius, D.J.; Hordijk, J. Plasmids carrying antimicrobial resistance genes in Enterobacteriaceae. J. Antimicrob. Chemother. 2018, 73, 1121–1137. [Google Scholar] [CrossRef] [PubMed]
- Hartard, C.; Rivet, R.; Banas, S.; Gantzer, C. Occurrence of and sequence variation among F-specific RNA bacteriophage subgroups in feces and wastewater of urban and animal origins. Appl. Environ. Microbiol. 2015, 81, 6505–6515. [Google Scholar] [CrossRef]
- Calci, K.R.; Burkhardt, W., III; Watkins, W.D.; Rippey, S.R. Occurrence of Male-Specific Bacteriophage in Feral and Domestic Animal Wastes, Human Feces, and Human-Associated Wastewaters. Appl. Environ. Microbiol. 1998, 64, 5027–5029. [Google Scholar]
- Nuttall, D.; Maker, D.; Colleran, E. A method for the direct isolation of IncH plasmid-dependent bacteriophages. Lett. Appl. Microbiol. 1987, 5, 37–40. [Google Scholar] [CrossRef]
- Bradley, D.E.; Sirgel, F.A.; Coetzee, J.N.; Hedges, R.W.; Coetzee, W.F. Phages C-2 and J: IncC and IncJ plasmid-dependent phages, respectively. J. Gen. Microbiol. 1982, 128, 2485–2498. [Google Scholar] [CrossRef]
- Saren, A.M.; Ravantti, J.J.; Benson, S.D.; Burnett, R.M.; Paulin, L.; Bamford, D.H.; Bamford, J.K. A snapshot of viral evolution from genome analysis of the tectiviridae family. J. Mol. Biol. 2005, 350, 427–440. [Google Scholar] [CrossRef]
- Rumnieks, J.; Tars, K. Diversity of pili-specific bacteriophages: Genome sequence of IncM plasmid-dependent RNA phage M. BMC Microbiol. 2012, 12, 277. [Google Scholar] [CrossRef]
- Pumpens, P. Single-Stranded RNA Phages: From Molecular Biology to Nanotechnology, 1st ed.; CRC Press: Boca Raton, FL, USA, 2020. [Google Scholar] [CrossRef]
- Davis, J.; Strauss, J.H.; Sinnsheimer, R.L. Bacteriophage MS2, another RNA phage. Science 1961, 134, 1427. [Google Scholar]
- Watanabe, I.; Miyake, T.; Sakurai, T.; Shiba, T.; Ohno, T. Group characteristics of RNA phages. Proc. Jpn. Acad. 1967, 43, 210–213. [Google Scholar] [CrossRef]
- Kannoly, S.; Shao, Y.; Wang, I.-N. Rethinking the Evolution of Single-Stranded RNA (ssRNA) Bacteriophages Based on Genomic Sequences and Characterizations of Two R-Plasmid-Dependent ssRNA Phages, C-1 and Hgal1. J. Bacteriol. 2012, 194, 5073–5079. [Google Scholar] [CrossRef] [PubMed]
- Sirgel, F.; Coetzee, W. Characteristics and classification of IncC group plasmid-dependent phage C-1. Intervirology 1983, 20, 155–158. [Google Scholar] [CrossRef] [PubMed]
- Sirgel, F.A.; Coetzee, J.N.; Hedges, R.W.; Lecatsas, G. Phage C-1: An IncC group; plasmid-specific phage. J. Gen. Microbiol. 1981, 122, 155–160. [Google Scholar] [CrossRef]
- Coetzee, J.N.; Bradley, D.E.; Lecatsas, G.; Du Toit, L.; Hedges, R.W. Bacteriophage D: An IncD group plasmid-specific phage. J. Gen. Microbiol. 1985, 131, 3375–3383. [Google Scholar] [CrossRef]
- Coetzee, J.N.; Bradley, D.E.; Hedges, R.W. Phages I alpha and I2-2: IncI plasmid-dependent bacteriophages. J. Gen. Microbiol. 1982, 128, 2797–2804. [Google Scholar] [CrossRef]
- Coetzee, J.N.; Bradley, D.E.; Hedges, R.W.; Fleming, J.; Lecatsas, G. Bacteriophage M: An incompatibility group M plasmid-specific phage. J. Gen. Microbiol. 1983, 129, 2271–2276. [Google Scholar] [CrossRef]
- Coetzee, J.N.; Bradley, D.; Fleming, J.; Toit, L.D.; Hughes, V.; Hedges, R. Phage pilHα: A Phage Which Adsorbs to IncHI and IncHII Plasmid-coded Pili. Microbiology 1985, 131, 1115–1121. [Google Scholar]
- Maher, D.; Sherburne, R.; Taylor, D.E. Bacteriophages for incompatibility group H plasmids: Morphological and growth characteristics. Plasmid 1991, 26, 141–146. [Google Scholar] [CrossRef]
- Ruokoranta, T.M.; Grahn, A.M.; Ravantti, J.J.; Poranen, M.M.; Bamford, D.H. Complete genome sequence of the broad host range single-stranded RNA phage PRR1 places it in the Levivirus genus with characteristics shared with Alloleviviruses. J. Virol. 2006, 80, 9326–9330. [Google Scholar] [CrossRef]
- Bradley, D.E. Adsorption of bacteriophages specific for Pseudomonas aeruginosa R factors RP1 and R1822. Biochem. Biophys. Res. Commun. 1974, 57, 893–900. [Google Scholar] [CrossRef]
- Olsen, R.H.; Thomas, D.D. Characteristics and purification of PRR1, an RNA phage specific for the broad host range Pseudomonas R1822 drug resistance plasmid. J. Virol. 1973, 12, 1560–1567. [Google Scholar] [PubMed]
- Coetzee, J.N.; Bradley, D.E.; Hedges, R.W.; Hughes, Y.M.; McConnell, M.M.; Du Toit, L.; Tweehuysen, M. Bacteriophages Fiach, SR, SF: Phages which adsorb to pili encoded by plasmids of the S-complex. J. Gen. Microbiol. 1986, 132, 2907–2917. [Google Scholar] [PubMed]
- Bradley, D.E.; Coetzee, J.N.; Bothma, T.; Hedges, R.W. Phage t: A group T plasmid-dependent bacteriophage. J. Gen. Microbiol. 1981, 126, 397–403. [Google Scholar] [CrossRef] [PubMed]
- Daugelavicius, R.; Bamford, J.K.; Grahn, A.M.; Lanka, E.; Bamford, D.H. The IncP plasmid encoded cell envelope-associated DNA transfer complex increases cell permeability. J. Bacteriol. 1997, 179, 5195–5202. [Google Scholar] [CrossRef] [PubMed]
- Meynell, G.; Lawn, A. Filamentous Phages specific for the I Sex Factor. Nature 1968, 217, 1184–1186. [Google Scholar] [CrossRef]
- Bradley, D.E.; Coetzee, J.N.; Hedges, R.W. IncI2 plasmids specify sensitivity to filamentous bacteriophage IKe. J. Bacteriol. 1983, 154, 505–507. [Google Scholar] [CrossRef]
- Bradley, D.E. Morphology of pili determined by the N incompatibility group plasmid N3, and interaction with bacteriophages PR4 and IKe. Plasmid 1979, 2, 632–636. [Google Scholar] [CrossRef]
- Bradley, D.E.; Coetzee, J.N.; Bothma, T.; Hedges, R.W. Phage X: A plasmid-dependent, broad host range, filamentous bacterial virus. J. Gen. Microbiol. 1981, 126, 389–396. [Google Scholar] [CrossRef] [PubMed]
- Stanisich, V.A. The properties and host range of malespecific bacteriophage of Ps. aeruginosa. J. Gen. Microbiol. 1974, 84, 332–342. [Google Scholar] [CrossRef][Green Version]
- Coetzee, J.N.; Bradley, D.E.; Hedges, R.W.; Tweehuizen, M.; Du Toit, L. Phage tf-1: A filamentous bacteriophage specific for bacteria harbouring the IncT plasmid pIN25. J. Gen. Microbiol. 1987, 133, 953–960. [Google Scholar] [CrossRef] [PubMed]
- Jalasvuori, M.; Friman, V.-P.; Nieminen, A.; Bamford, J.K.H.; Buckling, A. Bacteriophage selection against a plasmid-encoded sex-apparatus leads to the loss of antibiotic resistance plasmids. Biol. Lett. 2011, 23, 902–905. [Google Scholar] [CrossRef] [PubMed]
- Colom, J.; Batista, D.; Baig, A.; Tang, Y.; Liu, S.; Yuan, F.; Belkhiri, A.; Marcelino, L.; Barbosa, F.; Rubio, M.; et al. Sex pilus specific bacteriophage to drive bacterial population towards antibiotic sensitivity. Sci. Rep. 2019, 9, 12616. [Google Scholar] [CrossRef] [PubMed]
- Ojala, V.; Laitalainen, J.; Jalasvuori, M. Fight evolution with evolution: Plasmid-dependent phages with a wide host range prevent the spread of antibiotic resistances. Evol. Appl. 2013, 6, 925–932. [Google Scholar] [CrossRef] [PubMed]
- Jalasvuori, M.; Penttinen, R. What can evolutionary rescue tell us about the emergence of new resistant bacteria? Future Microbiol. 2017, 12, 731–733. [Google Scholar] [CrossRef]
- May, T.; Tsuruta, K.; Okabe, S. Exposure of conjugative plasmid carrying Escherichia coli biofilms to male-specific bacteriophages. ISME J. 2011, 5, 771–775. [Google Scholar] [CrossRef]
- Mikonranta, L.; Buckling, A.; Jalasvuori, M.; Raymond, B. Targeting antibiotic resistant bacteria with phage reduces bacterial density in an insect host. Biol. Lett. 2019, 15, 20180895. [Google Scholar] [CrossRef]
- Sun, D.; Jeannot, K.; Xiao, Y.; Knapp, C.W. Editorial: Horizontal Gene Transfer Mediated Bacterial Antibiotic Resistance. Front. Microbiol. 2019, 10, 1933. [Google Scholar] [CrossRef]
- Jalasvuori, M. Silent rain: Does the atmosphere-mediated connectivity between microbiomes influence bacterial evolutionary rates? FEMS Microb. Ecol. 2020, 96, fiaa096. [Google Scholar]
- San Millan, A.; MacLean, R.C. Fitness Costs of Plasmids: A Limit to Plasmid Transmission. Microbiol. Spec. 2017, 5, 1–12. [Google Scholar]
- Bottery, M.J.; Wood, A.J.; Brockhurst, M.A. Selective conditions for a multidrug resistance plasmid depend on the sociality of antibiotic resistance. Antimicrob. Agents Chemother. 2016, 60, 2524–2527. [Google Scholar] [CrossRef] [PubMed]
- Cairns, J.; Jalasvuori, M.; Ojala, V.; Brockhurst, M.; Hiltunen, T. Conjugation is necessary for a bacterial plasmid to survive under protozoan predation. Biol. Lett. 2016, 12, 20150953. [Google Scholar] [CrossRef] [PubMed]
- Schmidt, C. Phage therapy’s latest makeover. Nat. Biotechnol. 2019, 37, 581–586. [Google Scholar] [CrossRef] [PubMed]



{kind=link}
{kind=link}
| Family Genus | Bacteriophage | Morphology | Size | Genome | Genome Length (bp) | Example Host a | Incompatibility | Phage Attachment Site | References |
|---|---|---|---|---|---|---|---|---|---|
| Leviviridae | icosahedral capsid | 20–28 nm | ssRNA | 3400–4200 | [51] | ||||
| Levivirus | |||||||||
| MS2 | Escherichia coli | IncF | Sides of pili | [52] | |||||
| Levivirus | |||||||||
| Qβ | Escherichia coli | IncF | Sides of pili | [53] | |||||
| Unclassified | |||||||||
| C-1 | Salmonella typhimurium | IncC | Sides of pili | [54,55,56] | |||||
| D | Escherichia coli | IncD | Sides of the end of pilus | [57] | |||||
| Hgal1 | Escherichia coli | IncH | Sides of pili | [47,54] | |||||
| Iɑ | Escherichia coli | IncI1 | Sides of pili | [58] | |||||
| M | Escherichia coli | IncM | Sides of pili | [50,59] | |||||
| pilHɑ | Escherichia coli | IncHI, IncHII | Sides of pili | [60,61] | |||||
| PRR1 | Pseudomonas | IncP | Sides and base of pili | [62,63,64] | |||||
| SR | Escherichia coli | IncS, IncFV | Sides of pili | [65] | |||||
| t | Escherichia coli | IncT | Sides of pili | [66] | |||||
| Tectiviridae | icosahedral capsid, membrane-containing | 63 nm | dsDNA | ||||||
| PRD1 | 15000 | Escherichia coli | IncPɑ (IncN, IncW) | Mpf (mating pair formation) complex | [22,67] | ||||
| Inoviridae | filamentous | 6–8 nm × 700–2000 nm | ssDNA | 6000–12000 | |||||
| C-2 | Salmonella typhimurium | IncC | Sides of pili | [48] | |||||
| M13 | Escherichia coli | IncF | Tip of pili | [29,30] | |||||
| If1 | Escherichia coli | IncI | Tip of pili | [68] | |||||
| Ike | Escherichia coli | IncN, IncI2 | Tip of pili | [69,70] | |||||
| X | Escherichia coli | IncX (IncI2, IncM, IncN, IncP-1, IncW) | Tip of pili | [71] | |||||
| Pf3 | Pseudomonas aeruginosa | IncP-1 | Sides of pili | [63,72] | |||||
| SF | Escherichia coli | IncS, IncFI-V, IncD | Tip of pili | [65] | |||||
| tf-1 | Escherichia coli | IncT | Point of pili | [73] | |||||
| Unclassified | |||||||||
| J | 40 nm, 18 nm non-contractile tail | Escherichia coli | IncJ, IncC, IncD | Sides of pili | [48] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Penttinen, R.; Given, C.; Jalasvuori, M. Indirect Selection against Antibiotic Resistance via Specialized Plasmid-Dependent Bacteriophages. Microorganisms 2021, 9, 280. https://doi.org/10.3390/microorganisms9020280
Penttinen R, Given C, Jalasvuori M. Indirect Selection against Antibiotic Resistance via Specialized Plasmid-Dependent Bacteriophages. Microorganisms. 2021; 9(2):280. https://doi.org/10.3390/microorganisms9020280
Chicago/Turabian StylePenttinen, Reetta, Cindy Given, and Matti Jalasvuori. 2021. "Indirect Selection against Antibiotic Resistance via Specialized Plasmid-Dependent Bacteriophages" Microorganisms 9, no. 2: 280. https://doi.org/10.3390/microorganisms9020280
APA StylePenttinen, R., Given, C., & Jalasvuori, M. (2021). Indirect Selection against Antibiotic Resistance via Specialized Plasmid-Dependent Bacteriophages. Microorganisms, 9(2), 280. https://doi.org/10.3390/microorganisms9020280