Meta-Analysis of Microbial Communities in Hot Springs: Recurrent Taxa and Complex Shaping Factors beyond pH and Temperature

 ,
,  ,
,  ,
, 

Abstract
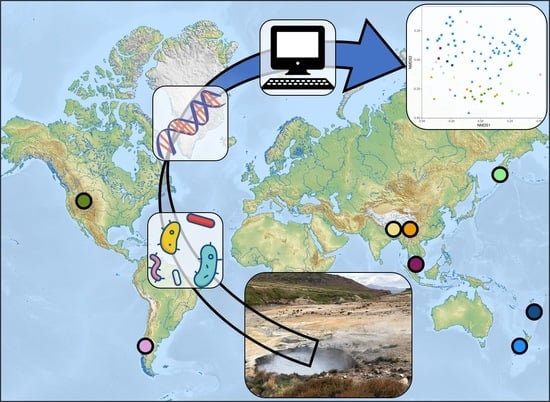
1. Introduction
2. Materials and Methods
2.1. Sample Collection and Physicochemical Determination
2.2. DNA Extraction and Amplicon Sequencing
2.3. Sequences Processing
2.4. Hot Spring Data Collection
2.5. Hot Springs Analysis
3. Results
3.1. Site Description and Physicochemical Characteristics of the Samples
3.2. Sequencing Analysis and Microbial Diversity
3.3. Hot Spring Microbial Structure and Shaping Factors
3.4. Co-Occurrence of Taxa Across Samples
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Hamamura, N.; Macur, R.E.; Korf, S.; Ackerman, G.; Taylor, W.P.; Kozubal, M.; Reysenbach, A.L.; Inskeep, W.P. Linking microbial oxidation of arsenic with detection and phylogenetic analysis of arsenite oxidase genes in diverse geothermal environments. Environ. Microbiol. 2009, 11, 421–431. [Google Scholar] [CrossRef] [PubMed]
- Sorensen, J.W.; Dunivin, T.K.; Tobin, T.C.; Shade, A. Ecological selection for small microbial genomes along a temperate-to-thermal soil gradient. Nat. Microbiol. 2019, 4, 55–61. [Google Scholar] [CrossRef] [PubMed]
- Belilla, J.; Moreira, D.; Jardillier, L.; Reboul, G.; Benzerara, K.; López-García, J.M.; Bertolino, P.; López-Archilla, A.I.; López-García, P. Hyperdiverse archaea near life limits at the polyextreme geothermal Dallol area. Nat. Ecol. Evol. 2019, 3, 1552–1561. [Google Scholar] [CrossRef] [PubMed]
- Skirnisdottir, S.; Hreggvidsson, G.O.; Hjörleifsdottir, S.; Marteinsson, V.T.; Petursdottir, S.K.; Holst, O.; Kristjansson, J.K. Influence of sulfide and temperature on species composition and community structure of hot spring microbial mats. Appl. Environ. Microbiol. 2000, 66, 2835–2841. [Google Scholar] [CrossRef] [PubMed]
- Cole, J.K.; Peacock, J.P.; Dodsworth, J.A.; Williams, A.J.; Thompson, D.B.; Dong, H.; Wu, G.; Hedlund, B.P. Sediment microbial communities in Great Boiling Spring are controlled by temperature and distinct from water communities. ISME J. 2013, 7, 718–729. [Google Scholar] [CrossRef] [PubMed]
- Sharp, C.E.; Brady, A.L.; Sharp, G.H.; Grasby, S.E.; Stott, M.B.; Dunfield, P.F. Humboldt’s spa: Microbial diversity is controlled by temperature in geothermal environments. ISME J. 2014, 8, 1166–1174. [Google Scholar] [CrossRef] [PubMed]
- Pagaling, E.; Grant, W.D.; Cowan, D.A.; Jones, B.E.; Ma, Y.; Ventosa, A.; Heaphy, S. Bacterial and archaeal diversity in two hot spring microbial mats from the geothermal region of Tengchong, China. Extremophiles 2012, 16, 607–618. [Google Scholar] [CrossRef] [PubMed]
- Menzel, P.; Gudbergsdóttir, S.R.; Rike, A.G.; Lin, L.; Zhang, Q.; Contursi, P.; Moracci, M.; Kristjansson, J.K.; Bolduc, B.; Gavrilov, S.; et al. Comparative Metagenomics of Eight Geographically Remote Terrestrial Hot Springs. Microb. Ecol. 2015, 70, 411–424. [Google Scholar] [CrossRef] [PubMed]
- Power, J.F.; Carere, C.R.; Lee, C.K.; Wakerley, G.L.J.; Evans, D.W.; Button, M.; White, D.; Climo, M.D.; Hinze, A.M.; Morgan, X.C.; et al. Microbial biogeography of 925 geothermal springs in New Zealand. Nat. Commun. 2018, 9. [Google Scholar] [CrossRef] [PubMed]
- Uribe-Lorío, L.; Brenes-Guillén, L.; Hernández-Ascencio, W.; Mora-Amador, R.; González, G.; Ramírez-Umaña, C.J.; Díez, B.; Pedrós-Alió, C. The influence of temperature and pH on bacterial community composition of microbial mats in hot springs from Costa Rica. Microbiologyopen 2019, 8, 1–26. [Google Scholar] [CrossRef] [PubMed]
- Mathur, J.; Bizzoco, R.W.; Ellis, D.G.; Lipson, D.A.; Poole, A.W.; Levine, R.; Kelley, S.T. Effects of abiotic factors on the phylogenetic diversity of bacterial communities in acidic thermal springs. Appl. Environ. Microbiol. 2007, 73, 2612–2623. [Google Scholar] [CrossRef] [PubMed]
- Purcell, D.; Sompong, U.; Yim, L.C.; Barraclough, T.G.; Peerapornpisal, Y.; Pointing, S.B. The effects of temperature, pH and sulphide on the community structure of hyperthermophilic streamers in hot springs of northern Thailand. FEMS Microbiol. Ecol. 2007, 60, 456–466. [Google Scholar] [CrossRef] [PubMed]
- Fontaneto, D.; Hortal, J. Microbial biogeography: Is everything small everywhere? In Microbial Ecological Theory: From Individuals to Ecosystems; Ogilvie, L.A., Hirsch, P.R., Eds.; Horizon Scientific Press: Norwich, UK, 2012; pp. 88–98. [Google Scholar]
- Jones, D.S.; Schaperdoth, I.; Macalady, J.L. Biogeography of sulfur-oxidizing Acidithiobacillus populations in extremely acidic cave biofilms. ISME J. 2016, 10, 2879–2891. [Google Scholar] [CrossRef] [PubMed]
- Brock, T.D. Thermophilic Microorganisms and Life at High Temperatures, Springer Series in Microbiology; Springer: Berlin/Heidelberg, Germany, 1978; Volume 43, ISBN 978-1-4612-6286-2. [Google Scholar]
- Huber, R.; Wilharm, T.; Huber, D.; Trincone, A.; Burggraf, S.; König, H.; Reinhard, R.; Rockinger, I.; Fricke, H.; Stetter, K.O. Aquifex pyrophilus gen. nov. sp. nov., Represents a Novel Group of Marine Hyperthermophilic Hydrogen-Oxidizing Bacteria. Syst. Appl. Microbiol. 1992, 15, 340–351. [Google Scholar] [CrossRef]
- Reysenbach, A.-L.; Huber, R.; Stetter, K.O.; Ishii, M.; Kawasumi, T.; Igarashi, Y.; Eder, W.; L’Haridon, S.; Jeanthon, C. Phylum BI. Aquificae phy. nov. In Bergey’s Manual® of Systematic Bacteriology; Springer: Berlin/Heidelberg, Germany, 2001; pp. 359–367. [Google Scholar]
- Ferrera, I.; Longhorn, S.; Banta, A.B.; Liu, Y.; Preston, D.; Reysenbach, A.L. Diversity of 16S rRNA gene, ITS region and aclB gene of the Aquificales. Extremophiles 2007, 11, 57–64. [Google Scholar] [CrossRef] [PubMed]
- Elleuche, S.; Schröder, C.; Sahm, K.; Antranikian, G. Extremozymes-biocatalysts with unique properties from extremophilic microorganisms. Curr. Opin. Biotechnol. 2014, 29, 116–123. [Google Scholar] [CrossRef] [PubMed]
- Raddadi, N.; Cherif, A.; Daffonchio, D.; Neifar, M.; Fava, F. Biotechnological applications of extremophiles, extremozymes and extremolytes. Appl. Microbiol. Biotechnol. 2015, 99, 7907–7913. [Google Scholar] [CrossRef] [PubMed]
- Bertrand, J.C.; Caumette, P.; Lebaron, P.; Matheron, R.; Normand, P.; Sime-Ngando, T. Environmental Microbiology: Fundamentals and Applications; Springer: Berlin/Heidelberg, Germany, 2015; ISBN 9789401791182. [Google Scholar]
- Albarracín, V.H.; Gärtner, W.; Farias, M.E. Forged under the Sun: Life and Art of Extremophiles from Andean Lakes. Photochem. Photobiol. 2016, 92, 14–28. [Google Scholar] [CrossRef] [PubMed]
- Merino, N.; Aronson, H.S.; Bojanova, D.P.; Feyhl-Buska, J.; Wong, M.L.; Zhang, S.; Giovannelli, D. Living at the extremes: Extremophiles and the limits of life in a planetary context. Front. Microbiol. 2019, 10, 780. [Google Scholar] [CrossRef] [PubMed]
- DasSarma, S.; DasSarma, P.; Laye, V.J.; Schwieterman, E.W. Extremophilic models for astrobiology: Haloarchaeal survival strategies and pigments for remote sensing. Extremophiles 2020, 24, 31–41. [Google Scholar] [CrossRef] [PubMed]
- Pesce, A.H. The Domuyo geothermal area, Neuquén, Argentina. Trans. Geotherm. Resour. Counc. 2013, 37, 309–313. [Google Scholar]
- Lavalle, L.; Chiacchiarini, P.; Pogliani, C.; Donati, E. Isolation and characterization of acidophilic bacteria from Patagonia, Argentina. Process Biochem. 2005, 40, 1095–1099. [Google Scholar] [CrossRef]
- Chiacchiarini, P.; Lavalle, L.; Giaveno, A.; Donati, E. First assessment of acidophilic microorganisms from geothermal Copahue-Caviahue system. Hydrometallurgy 2010, 104, 334–341. [Google Scholar] [CrossRef]
- Urbieta, M.S.; González Toril, E.; Aguilera, A.; Giaveno, M.A.; Donati, E. First Prokaryotic Biodiversity Assessment Using Molecular Techniques of an Acidic River in Neuquén, Argentina. Microb. Ecol. 2012, 64, 91–104. [Google Scholar] [CrossRef] [PubMed]
- Urbieta, M.S.; Toril, E.G.; Alejandra Giaveno, M.; Bazán, Á.A.; Donati, E.R. Archaeal and bacterial diversity in five different hydrothermal ponds in the Copahue region in Argentina. Syst. Appl. Microbiol. 2014, 37, 429–441. [Google Scholar] [CrossRef] [PubMed]
- Urbieta, M.; Porati, G.; Segretín, A.; González-Toril, E.; Giaveno, M.; Donati, E. Copahue Geothermal System: A Volcanic Environment with Rich Extreme Prokaryotic Biodiversity. Microorganisms 2015, 3, 344–363. [Google Scholar] [CrossRef] [PubMed]
- Lopez Bedogni, G.; Massello, F.L.; Giaveno, A.; Donati, E.R.; Urbieta, M.S. A deeper look into the biodiversity of the extremely acidic copahue volcano-río agrio system in Neuquén, Argentina. Microorganisms 2020, 8, 58. [Google Scholar] [CrossRef] [PubMed]
- Giaveno, M.A.; Urbieta, M.S.; Ulloa, J.R.; González Toril, E.; Donati, E.R. Physiologic Versatility and Growth Flexibility as the Main Characteristics of a Novel Thermoacidophilic Acidianus Strain Isolated from Copahue Geothermal Area in Argentina. Microb. Ecol. 2013, 65, 336–346. [Google Scholar] [CrossRef] [PubMed]
- Willis, G.; Yaakop, A.S.; Chan, C.S.; Urbieta, M.S.; Chan, K.G.; Ee, R.; Tan-Guan-Sheng, A.; Goh, K.M.; Donati, E.R. Draft genome sequence of the sulfate-reducing bacterium Desulfotomaculum copahuensis strain CINDEFI1 isolated from the geothermal Copahue system, Neuquén, Argentina. Genome Announc. 2016, 4, 1–2. [Google Scholar] [CrossRef]
- Cavello, I.; Urbieta, M.S.; Segretin, A.B.; Giaveno, A.; Cavalitto, S.; Donati, E.R. Assessment of Keratinase and Other Hydrolytic Enzymes in Thermophilic Bacteria Isolated from Geothermal Areas in Patagonia Argentina. Geomicrobiol. J. 2018, 35, 156–165. [Google Scholar] [CrossRef]
- Klindworth, A.; Pruesse, E.; Schweer, T.; Peplies, J.; Quast, C.; Horn, M.; Glöckner, F.O. Evaluation of general 16S ribosomal RNA gene PCR primers for classical and next-generation sequencing-based diversity studies. Nucleic Acids Res. 2013, 41, 1–11. [Google Scholar] [CrossRef] [PubMed]
- R Core Team. R: A Language and Environment for Statistical Computing. R Foundation for Statistical Computing, Austria, 2018. Available online: https://www.R-project.org/ (accessed on 15 October 2019).
- Callahan, B.J.; Mcmurdie, P.J.; Rosen, M.J.; Han, A.W.; A, A.J.; Holmes, S.P. DADA2: High resolution sample inference from Illumina amplicon data. Nat. Methods 2016, 13, 581–583. [Google Scholar] [CrossRef] [PubMed]
- Oksanen, J.; Blanchet, F.G.; Friendly, M.; Kindt, R.; Legrende, P.; McGlinn, D.; Minchin, P.R.; O’Hara, R.B.; Simpson, G.L.; Solymos, P.; et al. vegan: Community Ecology Package. R Package Version 2.5-2. 2019. Available online: https://CRAN.R-project.org/package=vegan (accessed on 15 October 2019).
- McMurdie, P.J.; Holmes, S. Phyloseq: An R Package for Reproducible Interactive Analysis and Graphics of Microbiome Census Data. PLoS ONE 2013. [Google Scholar] [CrossRef] [PubMed]
- Jiang, Z.; Li, P.; Jiang, D.; Dai, X.; Zhang, R.; Wang, Y.; Wang, Y. Microbial community structure and arsenic biogeochemistry in an acid vapor-formed spring in Tengchong geothermal area, China. PLoS ONE 2016, 11, 1–16. [Google Scholar] [CrossRef] [PubMed]
- Colman, D.R.; Feyhl-Buska, J.; Fecteau, K.M.; Xu, H.; Shock, E.L.; Boyd, E.S. Ecological differentiation in planktonic and sediment-associated chemotrophic microbial populations in Yellowstone hot springs. FEMS Microbiol. Ecol. 2016, 92, 1–13. [Google Scholar] [CrossRef] [PubMed]
- Chan, C.S.; Chan, K.G.; Ee, R.; Hong, K.W.; Urbieta, M.S.; Donati, E.R.; Shamsir, M.S.; Goh, K.M. Effects of physiochemical factors on prokaryotic Biodiversity in Malaysian circumneutral hot springs. Front. Microbiol. 2017, 8, 1252. [Google Scholar] [CrossRef] [PubMed]
- Stewart, L.C.; Stucker, V.K.; Stott, M.B.; de Ronde, C.E.J. Marine-influenced microbial communities inhabit terrestrial hot springs on a remote island volcano. Extremophiles 2018, 22, 687–698. [Google Scholar] [CrossRef] [PubMed]
- Najar, I.N.; Sherpa, M.T.; Das, S.; Das, S.; Thakur, N. Microbial ecology of two hot springs of Sikkim: Predominate population and geochemistry. Sci. Total Environ. 2018, 637–638, 730–745. [Google Scholar] [CrossRef] [PubMed]
- Mardanov, A.V.; Gumerov, V.M.; Beletsky, A.V.; Ravin, N.V. Microbial diversity in acidic thermal pools in the Uzon Caldera, Kamchatka. Antonie van Leeuwenhoek. Int. J. Gen. Mol. Microbiol. 2018, 111, 35–43. [Google Scholar] [CrossRef]
- Dufrêne, M.; Legendre, P. Species assemblages and indicator species: The need for a flexible asymmetrical approach. Ecol. Monogr. 1997, 67, 345–366. [Google Scholar] [CrossRef]
- De Cáceres, M.; Legendre, P. Associations between species and groups of sites: Indices and statistical inference. Ecology 2009, 90, 3566–3574. [Google Scholar] [CrossRef] [PubMed]
- Willis, G.; Hedrich, S.; Nancucheo, I.; Johnson, D.B.; Donati, E.R. Microbial Diversity in Acidic Anaerobic Sediments at the Geothermal Caviahue-Copahue System, Argentina. Adv. Mater. Res. 2013, 825, 7–10. [Google Scholar] [CrossRef]
- Lima, M.A.; Urbieta, M.S.; Donati, E. Arsenic-tolerant microbial consortia from sediments of Copahue Geothermal System with potential applications in bioremediation. J. Basic Microbiol. 2019. [Google Scholar] [CrossRef] [PubMed]
- Panarello, H.O. Características isotópicas y termodinámicas de reservorio del campo geotérmico Copahue-Caviahue, provincia del Neuquén. Rev. la Asoc. Geol. Argentina 2002, 57, 328–340. [Google Scholar]
- Gammons, C.H.; Wood, S.A.; Pedrozo, F.; Varekamp, J.C.; Nelson, B.J.; Shope, C.L.; Baffico, G. Hydrogeochemistry and rare earth element behavior in a volcanically acidified watershed in Patagonia, Argentina. Chem. Geol. 2005, 222, 249–267. [Google Scholar] [CrossRef]
- Varekamp, J.C.; Ouimette, A.P.; Herman, S.W.; Flynn, K.S.; Bermudez, A.; Delpino, D. Naturally acid waters from Copahue volcano, Argentina. Appl. Geochem. 2009, 24, 208–220. [Google Scholar] [CrossRef]
- Pedrozo, F.L.; Diaz, M.M.; Temporetti, P.F.; Baffico, G.D.; Beamud, S.G. Características limnológicas de un sistema ácido: Río Agrio-Lago Caviahue, Provincia del Neuquén, Argentina. Ecol. Austral. 2010, 20, 173–184. [Google Scholar]
- Farnfield, H.R.; Marcilla, A.L.; Ward, N.I. Arsenic speciation and trace element analysis of the volcanic río Agrio and the geothermal waters of Copahue, Argentina. Sci. Total Environ. 2012, 433, 371–378. [Google Scholar] [CrossRef] [PubMed]
- Kozich, J.J.; Westcott, S.L.; Baxter, N.T.; Highlander, S.K.; Schloss, P.D. Development of a Dual-Index Sequencing Strategy and Curation Pipeline for Analyzing Amplicon Sequence Data on the MiSeq Illumina Sequencing Platform. Appl. Environ. Microbiol. 2013, 79, 5112–5120. [Google Scholar] [CrossRef] [PubMed]
- Bogdanova, T.I.; Tsaplina, I.A.; Kondrat’eva, T.F.; Duda, V.I.; Suzina, N.E.; Melamud, V.S.; Tourova, T.P.; Karavaiko, G.I. Sulfobacillus thermotolerans sp. nov,. a thermotolerant, chemolithotrophic bacterium. Int. J. Syst. Evol. Microbiol. 2006, 56, 1039–1042. [Google Scholar] [CrossRef] [PubMed]
- Coram, N.J.N.; Rawlings, D.E. Molecular relationship between two groups of the genus Leptospirillum and the finding that &hellip. Appl. Environ. Microbiol. 2002, 68, 838–845. [Google Scholar] [CrossRef] [PubMed]
- Reysenbach, A.L.; Liu, Y.; Lindgren, A.R.; Wagner, I.D.; Sislak, C.D.; Mets, A.; Schouten, S. Mesoaciditoga lauensis gen. nov., sp. nov., a moderately thermoacidophilic member of the order Thermotogales from a deep-sea hydrothermal vent. Int. J. Syst. Evol. Microbiol. 2013, 63, 4724–4729. [Google Scholar] [CrossRef] [PubMed]
- Stackebrandt, E. The Family Lachnospiraceae. In The Prokaryotes; Springer: Berlin/Heidelberg, Germany, 2014; pp. 197–201. ISBN 9783642301209. [Google Scholar]
- Yabuuchi, E.; Kosako, Y. Sphingomonas. In Bergey’s Manual of Systematics of Archaea and Bacteria; John Wiley & Sons, Ltd.: Chichester, UK, 2015; pp. 1–39. ISBN 9781118960608. [Google Scholar]
- Ravot, G.; Magot, M.; Fardeau, M.; Patel, B.K.C.; Thomas, P.; Garcia, J.-L.; Ollivierl, B. Anaerobic, Thiosulfate-Reducing Bacterium From an Oil-Producing Well. Int. J. Syst. Bacteriol. 1999, 49, 1141–1147. [Google Scholar] [CrossRef] [PubMed]
- Serrano, A.E.; Escudero, L.V.; Tebes-Cayo, C.; Acosta, M.; Encalada, O.; Fernández-Moroso, S.; Demergasso, C. First draft genome sequence of a strain from the genus Fusibacter isolated from Salar de Ascotán in Northern Chile. Stand. Genomic Sci. 2017, 12. [Google Scholar] [CrossRef] [PubMed]
- Koeck, D.E.; Ludwig, W.; Wanner, G.; Zverlov, V.V.; Liebl, W.; Schwarz, W.H. Herbinix hemicellulosilytica gen. Nov, sp. nov, a thermophilic cellulose-degrading bacterium isolated from a thermophilic biogas reactor. Int. J. Syst. Evol. Microbiol. 2015, 65, 2365–2371. [Google Scholar] [CrossRef] [PubMed]
- Koeck, D.E.; Hahnke, S.; Zverlov, V.V. Herbinix luporum sp. nov., a thermophilic cellulose-degrading bacterium isolated from a thermophilic biogas reactor. Int. J. Syst. Evol. Microbiol. 2016, 66, 4132–4137. [Google Scholar] [CrossRef] [PubMed]
- Liu, J.; Sui, Y.; Yu, Z.; Shi, Y.; Chu, H.; Jin, J.; Liu, X.; Wang, G. High throughput sequencing analysis of biogeographical distribution of bacterial communities in the black soils of northeast China. Soil Biol. Biochem. 2014, 70, 113–122. [Google Scholar] [CrossRef]
- Shade, A. Diversity is the question, not the answer. ISME J. 2017, 11, 1–6. [Google Scholar] [CrossRef] [PubMed]
- Kristjánsson, J.; Hreggvidsson, G. Ecology and habitats of extremophiles. World J. Microbiol. Biotechnol. 1995, 11, 17–25. [Google Scholar] [CrossRef] [PubMed]
- Pikuta, E.V.; Hoover, R.B.; Tang, J. Microbial extremophiles at the limits of life. Crit. Rev. Microbiol. 2007, 33, 183–209. [Google Scholar] [CrossRef] [PubMed]
- Orellana, R.; Macaya, C.; Bravo, G.; Dorochesi, F.; Cumsille, A.; Valencia, R.; Rojas, C.; Seeger, M. Living at the Frontiers of Life: Extremophiles in Chile and Their Potential for Bioremediation. Front. Microbiol. 2018, 9, 1–25. [Google Scholar] [CrossRef] [PubMed]
- Takacs-Vesbach, C.; Inskeep, W.P.; Jay, Z.J.; Herrgard, M.J.; Rusch, D.B.; Tringe, S.G.; Kozubal, M.A.; Hamamura, N.; Macur, R.E.; Fouke, B.W.; et al. Metagenome sequence analysis of filamentous microbial communities obtained from geochemically distinct geothermal channels reveals specialization of three aquificales lineages. Front. Microbiol. 2013, 4, 1–25. [Google Scholar] [CrossRef] [PubMed]
- Hedlund, B.P.; Reysenbach, A.L.; Huang, L.; Ong, J.C.; Liu, Z.; Dodsworth, J.A.; Ahmed, R.; Williams, A.J.; Briggs, B.R.; Liu, Y.; et al. Isolation of diverse members of the Aquificales from geothermal springs in Tengchong, China. Front. Microbiol. 2015, 6, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Barion, S.; Franchi, M.; Gallori, E.; Di Giulio, M. The first lines of divergence in the Bacteria domain were the hyperthermophilic organisms, the Thermotogales and the Aquificales, and not the mesophilic Planctomycetales. BioSystems 2007, 87, 13–19. [Google Scholar] [CrossRef] [PubMed]
- Wheaton, G.; Counts, J.; Mukherjee, A.; Kruh, J.; Kelly, R. The confluence of heavy metal biooxidation and heavy metal resistance: Implications for bioleaching by extreme thermoacidophiles. Minerals 2015, 5, 397–451. [Google Scholar] [CrossRef]
- Counts, J.; Vitko, N.; Kelly, R. Complete Genome Sequences of Extremely Thermoacidophilic Metal-Mobilizing Type Strain Members of the Archaeal Family Sulfolobaceae, Acidianus brierleyi DSM-1651, Acidianus sulfidivorans DSM-18786, and Metallosphaera hakonensis DSM-7519. Microbiol. Resour. Announc. 2018, 7, e00831-18. [Google Scholar] [CrossRef] [PubMed]
- Florentino, A.P.; Brienza, C.; Stams, A.J.M.; Sánchez-Andrea, I. Desulfurella amilsii sp. nov., a novel acidotolerant sulfur-respiring bacterium isolated from acidic river sediments. Int. J. Syst. Evol. Microbiol. 2016, 66, 1249–1253. [Google Scholar] [CrossRef] [PubMed]
- Brito, E.M.S.; Villegas-Negrete, N.; Sotelo-González, I.A.; Caretta, C.A.; Goñi-Urriza, M.; Gassie, C.; Hakil, F.; Colin, Y.; Duran, R.; Gutiérrez-Corona, F.; et al. Microbial diversity in Los Azufres geothermal field (Michoacán, Mexico) and isolation of representative sulfate and sulfur reducers. Extremophiles 2014, 18, 385–398. [Google Scholar] [CrossRef] [PubMed]
- Bonch-Osmolovskaya, E.A.; Sokolova, T.G.; Kostrikina, N.A.; Zavarzin, G.A. Desulfurella acetivorans gen. nov. and sp. nov. -a new thermophilic sulfur-reducing eubacterium. Arch. Microbiol. 1990, 153, 151–155. [Google Scholar] [CrossRef]
- Albuquerque, L.; Rainey, F.A.; da Costa, M.S. Thermus; in association with Bergey’s Manual Trust; John Wiley & Sons, Inc.: Hoboken, NJ, USA, 2018; ISBN 9781118960608. [Google Scholar]
- Pantazaki, A.A.; Pritsa, A.A.; Kyriakidis, D.A. Biotechnologically relevant enzymes from Thermus thermophilus. Appl. Microbiol. Biotechnol. 2002, 58, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Hedlund, B.P.; McDonald, A.I.; Lam, J.; Dodsworth, J.A.; Brown, J.R.; Hungate, B.A. Potential role of Thermus thermophilus and T. oshimai in high rates of nitrous oxide (N 2O) production in ~80°C hot springs in the US Great Basin. Geobiology 2011, 9, 471–480. [Google Scholar] [CrossRef] [PubMed]
- Zhou, E.M.; Xian, W.D.; Mefferd, C.C.; Thomas, S.C.; Adegboruwa, A.L.; Williams, N.; Murugapiran, S.K.; Dodsworth, J.A.; Ganji, R.; Li, M.M.; et al. Thermus sediminis sp. nov., a thiosulfate-oxidizing and arsenate-reducing organism isolated from Little Hot Creek in the Long Valley Caldera, California. Extremophiles 2018, 22, 983–991. [Google Scholar] [CrossRef] [PubMed]
- Merkel, A.Y.; Pimenov, N.V.; Rusanov, I.I.; Slobodkin, A.I.; Slobodkina, G.B.; Tarnovetckii, I.Y.; Frolov, E.N.; Dubin, A.V.; Perevalova, A.A.; Bonch-Osmolovskaya, E.A. Microbial diversity and autotrophic activity in Kamchatka hot springs. Extremophiles 2017, 21, 307–317. [Google Scholar] [CrossRef] [PubMed]
- Miroshnichenko, M.L.; Lebedinsky, A.V.; Chernyh, N.A.; Tourova, T.P.; Kolganova, T.V.; Spring, S.; Bonch-Osmolovskaya, E.A. Caldimicrobium rimae gen. nov., sp, nov., an extremely thermophilic, facultatively lithoautotrophic, anaerobic bacterium from the Uzon Caldera, Kamchatka. Int. J. Syst. Evol. Microbiol. 2009, 59, 1040–1044. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Kojima, H.; Umezawa, K.; Fukui, M. Caldimicrobium thiodismutans sp. nov., a sulfur-disproportionating bacterium isolated from a hot spring, and emended description of the genus Caldimicrobium. Int. J. Syst. Evol. Microbiol. 2016, 66, 1828–1831. [Google Scholar] [CrossRef] [PubMed]
- Hiras, J.; Wu, Y.W.; Eichorst, S.A.; Simmons, B.A.; Singer, S.W. Refining the phylum Chlorobi by resolving the phylogeny and metabolic potential of the representative of a deeply branching, uncultivated lineage. ISME J. 2016, 10, 833–845. [Google Scholar] [CrossRef] [PubMed]
- Thiel, V.; Costas, A.M.G.; Fortney, N.W.; Martinez, J.N.; Tank, M.; Roden, E.E.; Boyd, E.S.; Ward, D.M.; Hanada, S.; Bryant, D.A. “Candidates thermonerobacter thiotrophicus,” a non-phototrophic member of the Bacteroidetes/Chlorobi with dissimilatory sulfur metabolism in hot spring mat communities. Front. Microbiol. 2019, 10, 1–24. [Google Scholar] [CrossRef]
- Jungbluth, S.P.; Amend, J.P.; Rappé, M.S. Metagenome sequencing and 98 microbial genomes from Juan de Fuca Ridge flank subsurface fluids. Sci. Data 2017, 4, 1–11. [Google Scholar] [CrossRef]
- Nuñez, H.; Covarrubias, P.C.; Moya-Beltrán, A.; Issotta, F.; Atavales, J.; Acuña, L.G.; Johnson, D.B.; Quatrini, R. Detection, identification and typing of Acidithiobacillus species and strains: A review. Res. Microbiol. 2016, 167, 555–567. [Google Scholar] [CrossRef] [PubMed]
- Nuñez, H.; Moya-Beltrán, A.; Covarrubias, P.C.; Issotta, F.; Cárdenas, J.P.; González, M.; Atavales, J.; Acuña, L.G.; Johnson, D.B.; Quatrini, R. Molecular systematics of the Genus Acidithiobacillus: Insights into the phylogenetic structure and diversification of the taxon. Front. Microbiol. 2017, 8. [Google Scholar] [CrossRef] [PubMed]
- Huber, R.; Eder, W.; Heldwein, S.; Wanner, G.; Huber, H.; Rachel, R.; Stetter, K.O. Thermocrinis tuber gen. nov., sp. nov., a pink-filament-forming hyperthermophilic bacterium isolated from Yellowstone National Park. Appl. Environ. Microbiol. 1998, 64, 3576–3583. [Google Scholar] [CrossRef] [PubMed]
- Hetzer, A.; McDonald, I.R.; Morgan, H.W. Venenivibrio stagnispumantis gen. nov., sp. nov., a thermophilic hydrogen-oxidizing bacterium isolated from Champagne Pool, Waiotapu, New Zealand. Int. J. Syst. Evol. Aicrobiol. 2008, 58, 398–403. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
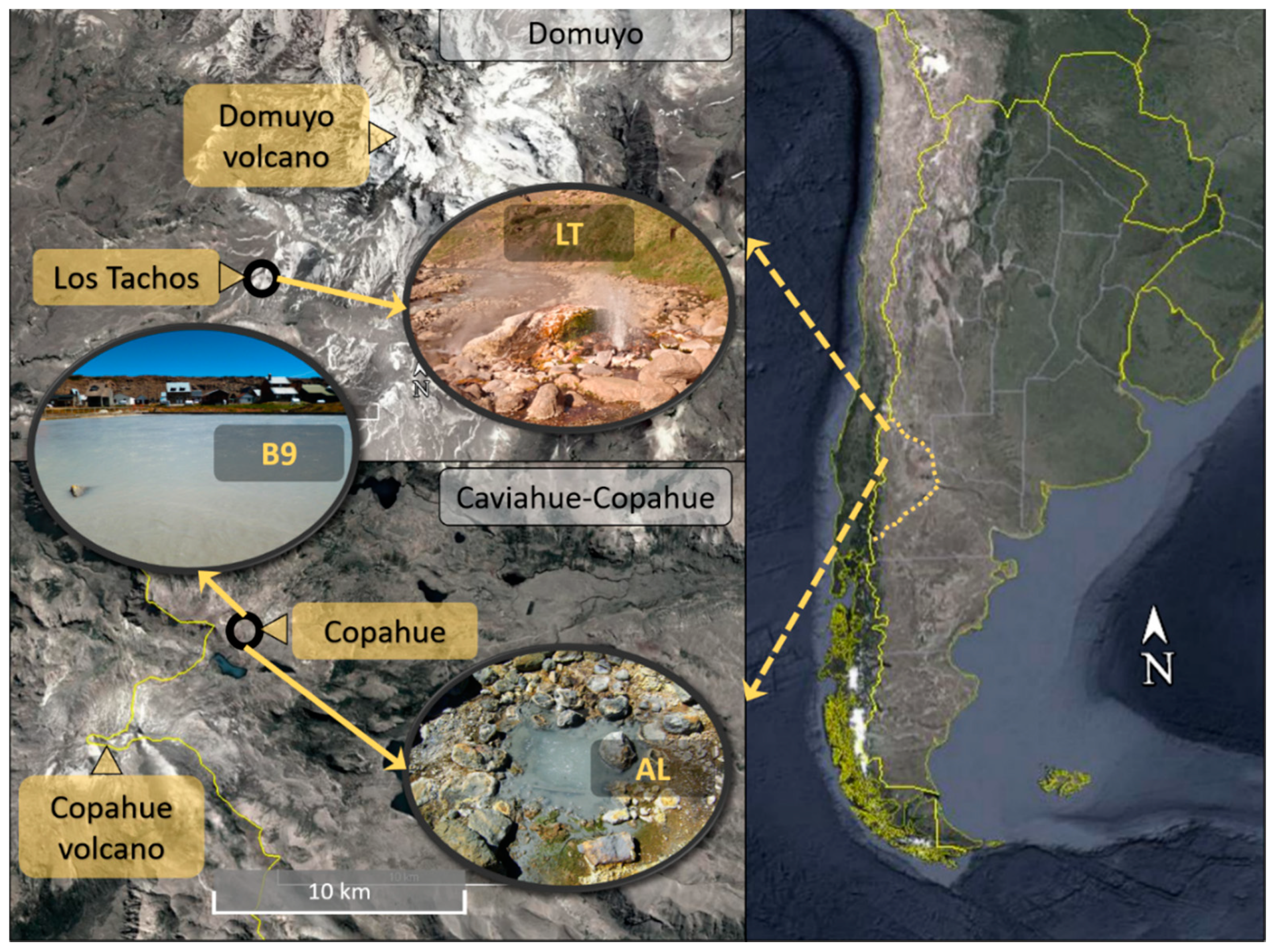
| ID | Sample | pH | T [°C] | Eh [mV] | Oxygen [mg/L] | Conductivity [mS] | Arsenic [mg/L] | Sulfide [mg/L] | Iron [mg/L] |
|---|---|---|---|---|---|---|---|---|---|
| AL | Agua de Limón | 2.36 | 71.1 | −67.6 | 0.3 | 4.66 | 0.05 | 0.5 | 76.13 |
| B9 | Baño 9 | 2.43 | 57.0 | −137.1 | 0.3 | 1.93 | 0.40 | 0.4 | 17.59 |
| LT | Los Tachos | 7.66 | 75.4 | 147.0 | 8.2 | 3.55 | 0.10 | 0.1 | 0.00 |
| Group | Taxa | √(IndVal) | A | B | p-Value |
|---|---|---|---|---|---|
| A | Hydrogenobaculum | 0.87 | 0.99 | 0.77 | 0.0002 |
| AH | Sulfolobaceae Family | 0.72 | 0.73 | 0.70 | 0.0046 |
| AH | Acinetobacter | 0.64 | 0.89 | 0.47 | 0.0032 |
| AT | Desulfurella | 0.72 | 0.86 | 0.60 | 0.0002 |
| AT | BSLdp215 | 0.68 | 0.99 | 0.47 | 0.0004 |
| N | Hydrothermae Phylum | 0.83 | 0.99 | 0.69 | 0.0002 |
| N | Thermus | 0.76 | 0.79 | 0.72 | 0.0004 |
| N | Caldimicrobium | 0.73 | 0.96 | 0.55 | 0.0002 |
| N | OPB56 | 0.72 | 0.94 | 0.55 | 0.0002 |
| N | Hydrogenobacter | 0.70 | 0.90 | 0.55 | 0.0004 |
| H | Anoxybacillus | 0.57 | 1.00 | 0.32 | 0.006 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Massello, F.L.; Chan, C.S.; Chan, K.-G.; Goh, K.M.; Donati, E.; Urbieta, M.S. Meta-Analysis of Microbial Communities in Hot Springs: Recurrent Taxa and Complex Shaping Factors beyond pH and Temperature. Microorganisms 2020, 8, 906. https://doi.org/10.3390/microorganisms8060906
Massello FL, Chan CS, Chan K-G, Goh KM, Donati E, Urbieta MS. Meta-Analysis of Microbial Communities in Hot Springs: Recurrent Taxa and Complex Shaping Factors beyond pH and Temperature. Microorganisms. 2020; 8(6):906. https://doi.org/10.3390/microorganisms8060906
Chicago/Turabian StyleMassello, Francisco L., Chia Sing Chan, Kok-Gan Chan, Kian Mau Goh, Edgardo Donati, and María Sofía Urbieta. 2020. "Meta-Analysis of Microbial Communities in Hot Springs: Recurrent Taxa and Complex Shaping Factors beyond pH and Temperature" Microorganisms 8, no. 6: 906. https://doi.org/10.3390/microorganisms8060906
APA StyleMassello, F. L., Chan, C. S., Chan, K.-G., Goh, K. M., Donati, E., & Urbieta, M. S. (2020). Meta-Analysis of Microbial Communities in Hot Springs: Recurrent Taxa and Complex Shaping Factors beyond pH and Temperature. Microorganisms, 8(6), 906. https://doi.org/10.3390/microorganisms8060906