Kinetics and New Mechanism of Azoxystrobin Biodegradation by an Ochrobactrum anthropi Strain SH14

 , and
, and 
Abstract
1. Introduction
2. Materials and Methods
2.1. Chemicals and Media
2.2. Isolation and Identification of Azoxystrobin-Degrading Strains
2.3. Growth and Biodegradation Experiments
2.4. Optimization of Biodegradation Conditions
2.5. Biodegradation Kinetics of Azoxystrobin
2.6. Biodegradation Kinetics of Various Strobilurin Fungicides
2.7. Identification of Azoxystrobin Metabolites
2.8. Biodegradation of Azoxystrobin in Soils
2.9. Analytical Methods
3. Results and Discussion
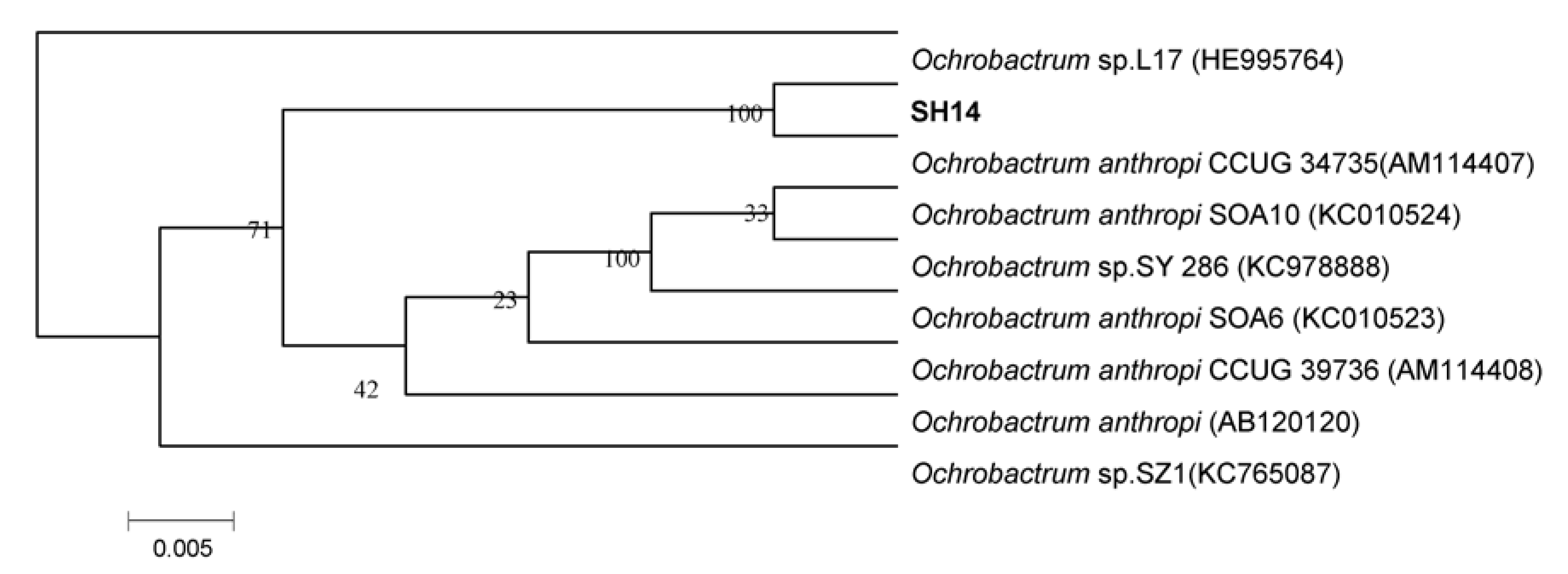
3.1. Isolation and Identification of Azoxystrobin-Degrading Strains
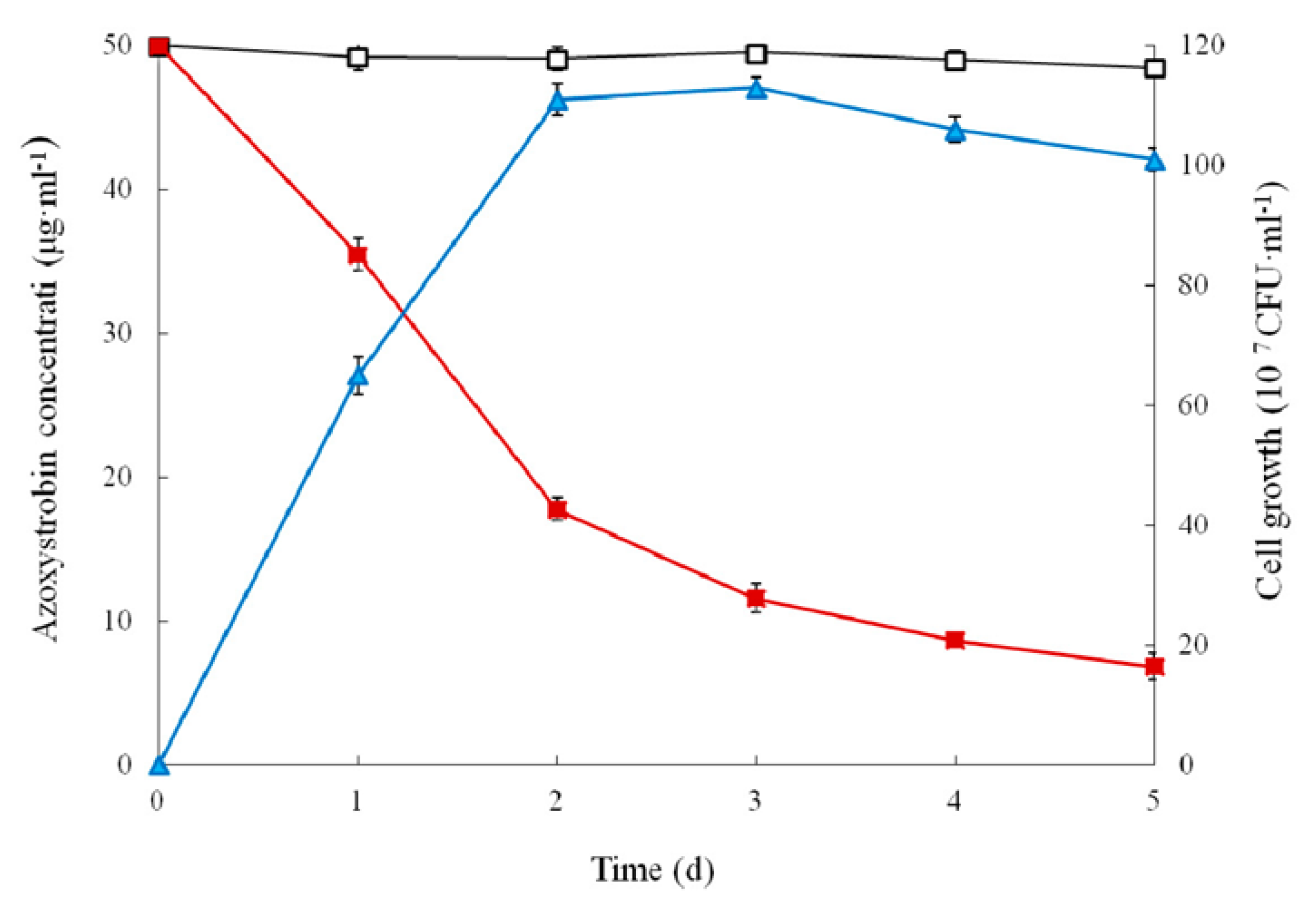
3.2. Growth and Utilization of Azoxystrobin by Strain SH14
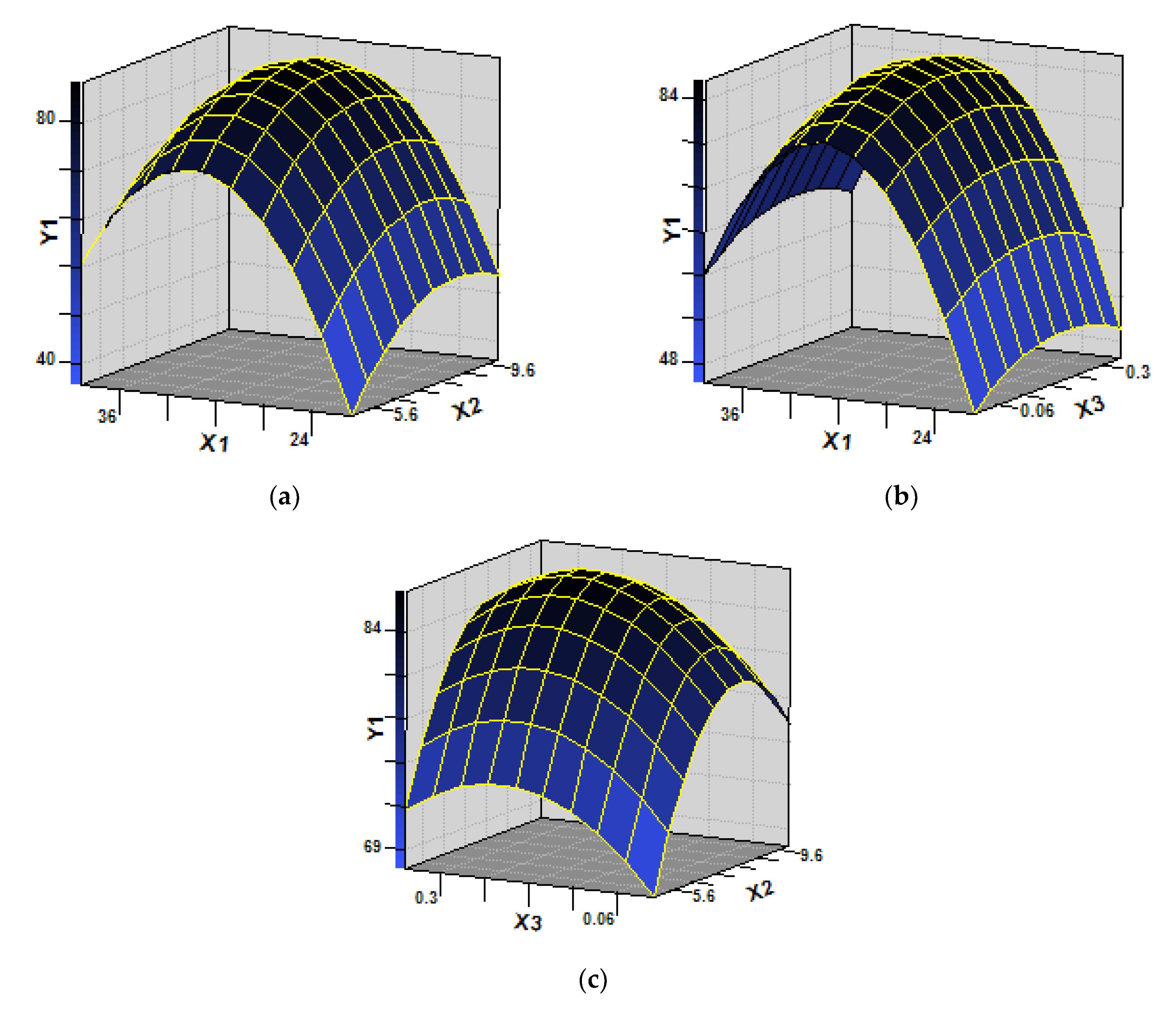
3.3. Optimization of Biodegradation Conditions
− 3.259362X22 − 1.80979X32
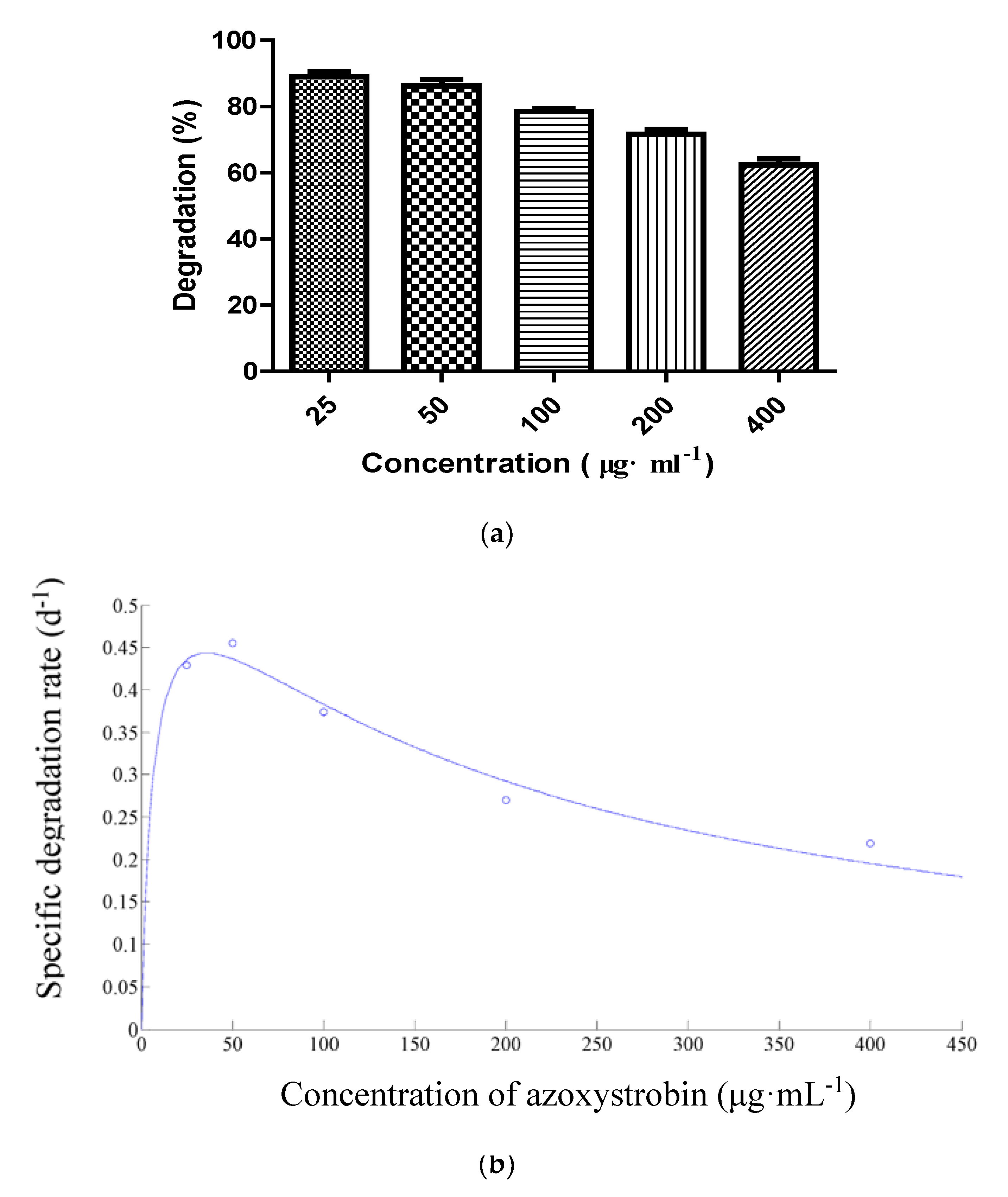
3.4. Biodegradation Kinetics of Azoxystrobin
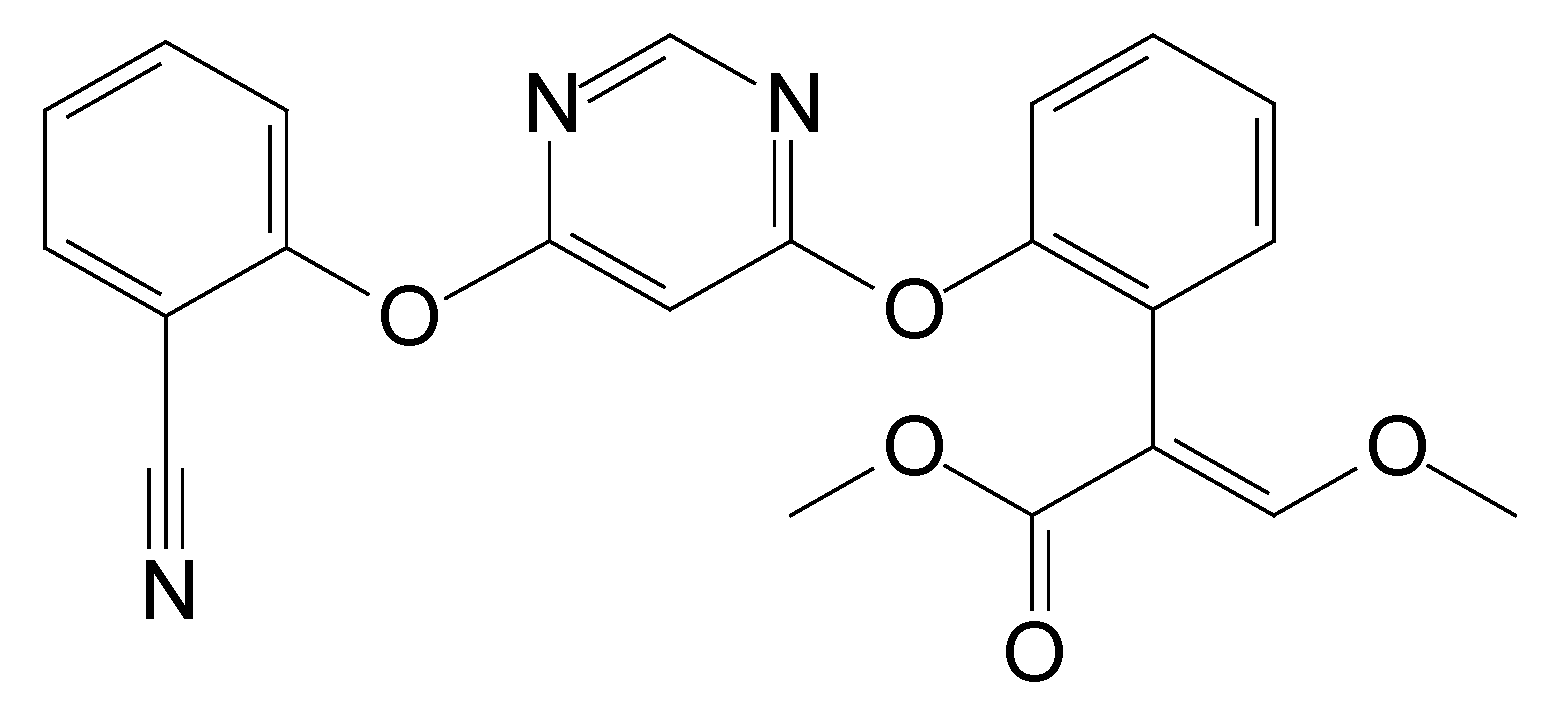
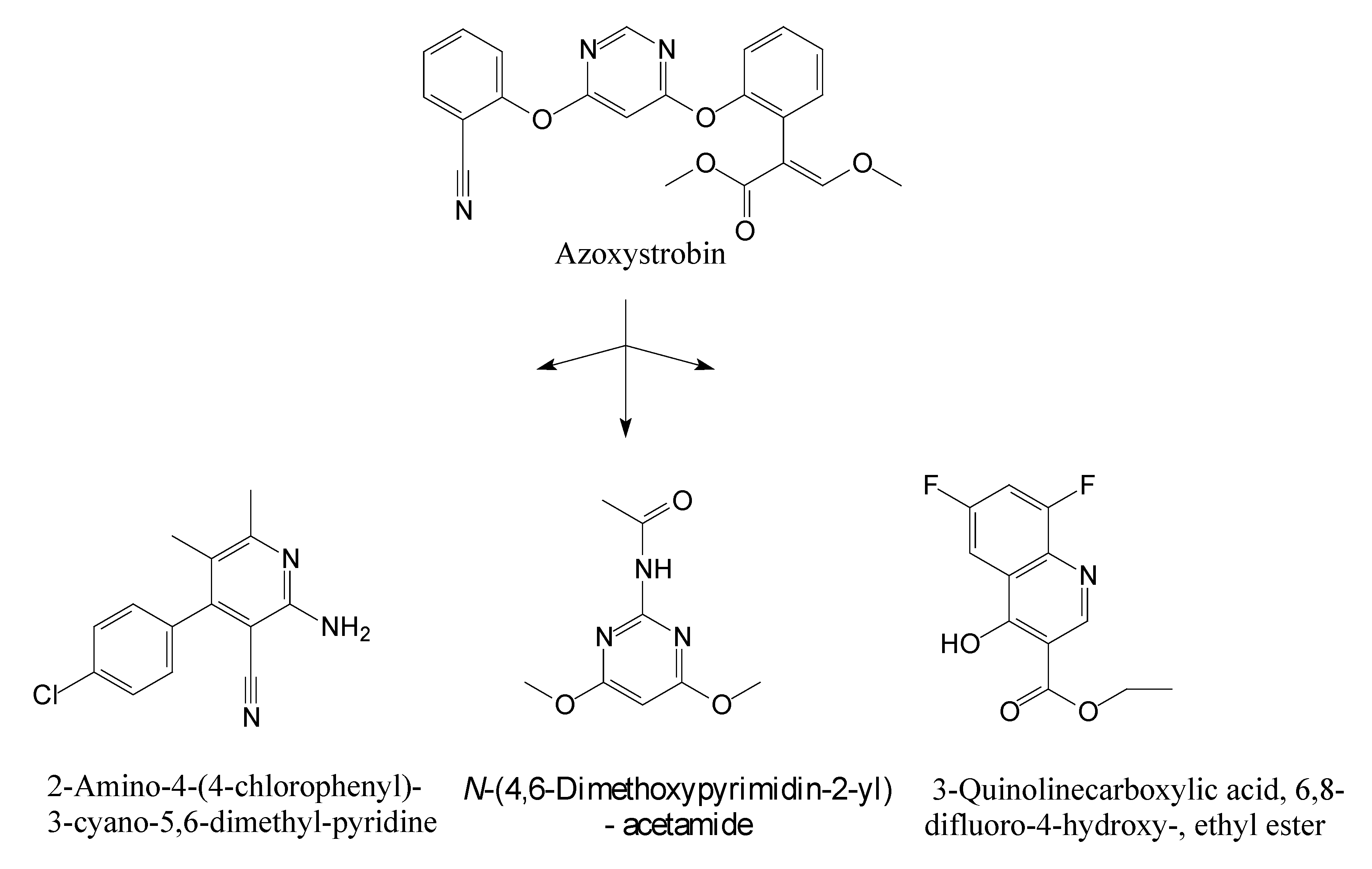
3.5. Identification of Metabolites
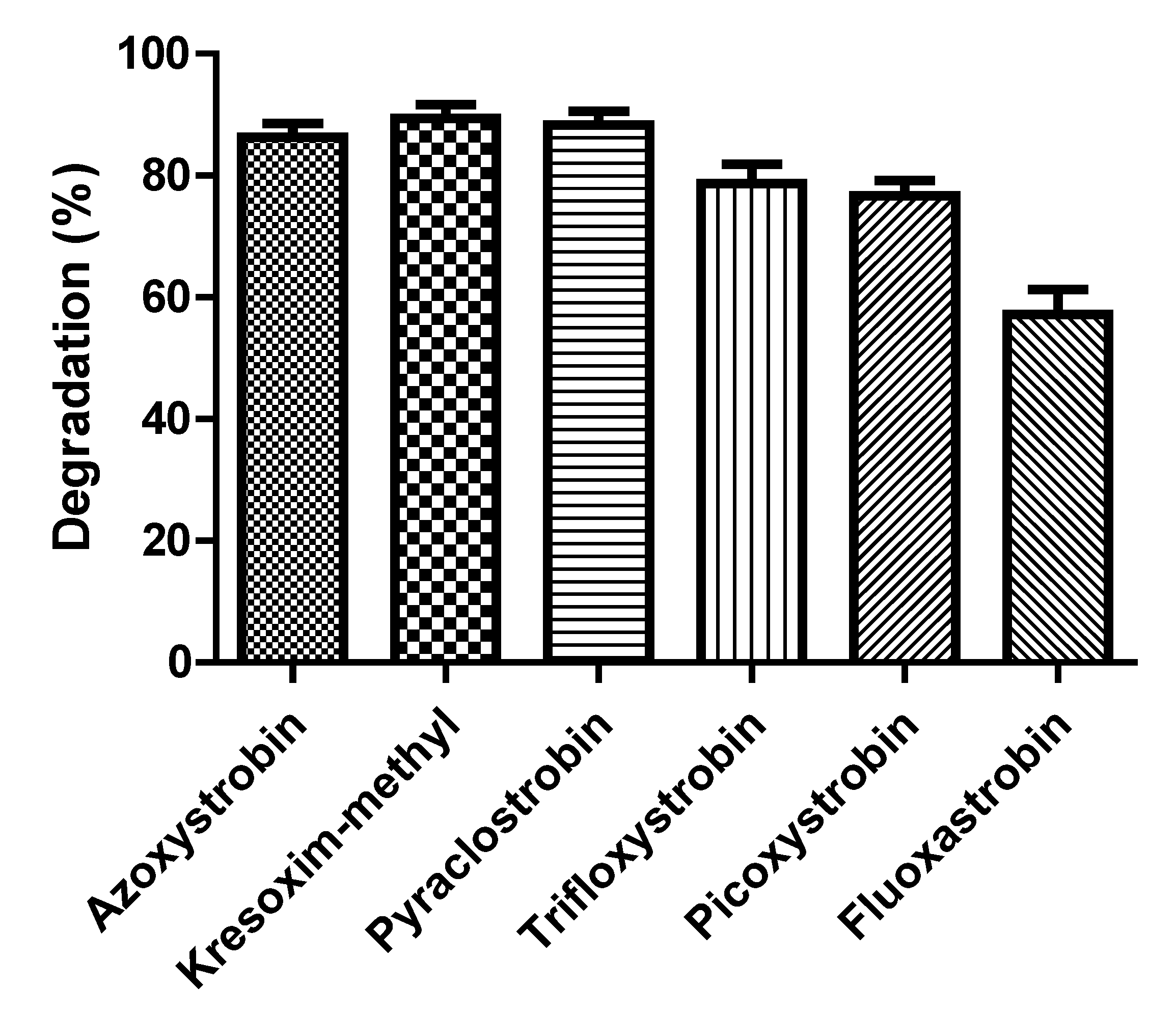
3.6. Biodegradation Kinetics of Various Strobilurin Fungicides
3.7. Biodegradation of Azoxystrobin in Soils
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Feng, Y.; Huang, Y.; Zhan, H.; Bhatt, P.; Chen, S. An overview of strobilurin fungicide degradation: Current status and future perspective. Front. Microbiol. 2020, 11, 389. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.; Yang, X.; Yuan, S.K.; Li, Y.F.; Zhang, A.F.; Yao, J.; Gao, T.C. Effect of azoxystrobin and kresoxim-methyl on rice blast and rice grain yield in China. Ann. Appl. Biol. 2015, 166, 434–443. [Google Scholar] [CrossRef]
- Balba, H. Review of strobilurin fungicide chemicals. J. Environ. Sci. Health B 2007, 42, 441–451. [Google Scholar] [CrossRef] [PubMed]
- Phillips McDougall AgriService. Products Section—2014 Market; Phillips McDougall-Agriservice: Midlothian, UK, 2015. [Google Scholar]
- EFSA. Conclusions on the peer review of the pesticide risk assessment of the active substance azoxystrobin. EFSA J. 2010, 8, 1542–1652. [Google Scholar] [CrossRef]
- Rodrigues, E.T.; Lopes, I.; Pardal, M.A. Occurrence, fate and effects of azoxystrobin in aquatic ecosystems: A review. Environ. Int. 2013, 53, 18–28. [Google Scholar] [CrossRef]
- Dijksterhuis, J.; van Doorn, T.; Samson, R.; Postma, J. Effects of seven fungicides on non-target aquatic fungi. Water Air Soil Pollut. 2011, 222, 421–425. [Google Scholar] [CrossRef]
- Warming, T.P.; Mulderij, G.; Christoffersen, K.S. Clonal variation in physiological responses of Daphnia magna to the strobilurin fungicide azoxystrobin. Environ. Toxicol. Chem. 2009, 28, 374–380. [Google Scholar] [CrossRef]
- Mostafalou, S.; Abdollahi, M. Pesticides and human chronic diseases: Evidences, mechanisms, and perspectives. Toxicol. Appl. Pharm. 2013, 268, 157–177. [Google Scholar] [CrossRef]
- Cayir, A.; Coskun, M.; Coskun, M. Micronuclei, nucleoplasmic bridges, and nuclear buds induced in human lymphocytes by the fungicide signum and its active ingredients (boscalid and pyraclostrobin). Environ. Toxicol. 2014, 29, 723–732. [Google Scholar] [CrossRef]
- Regueiro, J.; Olguin, N.; Simal-Gandara, J.; Suñol, C. Toxicity evaluation of new agricultural fungicides in primary cultured cortical neurons. Environ. Res. 2015, 140, 37–44. [Google Scholar] [CrossRef]
- Pearson, B.L.; Simon, J.M.; Mccoy, E.S.; Salazar, G.; Fragola, G.; Zylka, M.J. Identification of chemicals that mimic transcriptional changes associated with autism, brain aging and neurodegeneration. Nat. Commun. 2016, 7, 11173. [Google Scholar] [CrossRef] [PubMed]
- Bartlett, D.W.; Clough, J.M.; Godwin, J.R.; Hall, A.A.; Hamer, M.; Parr-Dobrzanski, B. The strobilurin fungicides. Pest Manag. Sci. 2002, 58, 649–662. [Google Scholar] [CrossRef] [PubMed]
- Gustafsson, K.; Blidberg, E.; Elfgren, I.K.; Hellström, A.; Kylin, H.; Gorokhova, E. Direct and indirect effects of the fungicide azoxystrobin in outdoor brackish water microcosms. Ecotoxicology 2010, 19, 431–444. [Google Scholar] [CrossRef]
- Jorgensen, L.F.; Kjaer, J.; Olsen, P.; Rosenbom, A.E. Leaching of azoxystrobin and its degradation product R234886 from Danish agricultural field sites. Chemosphere 2012, 88, 554–562. [Google Scholar] [CrossRef] [PubMed]
- Berenzen, N.; Lentzen-Godding, A.; Probst, M.; Schulz, H.; Schulz, R.; Liess, M. A comparison of predicted and measured levels of runoff-related pesticide concentrations in small lowland streams on a landscape level. Chemosphere 2005, 58, 683–691. [Google Scholar] [CrossRef] [PubMed]
- Garanzini, D.S.; Menone, M.L. Azoxystrobin causes oxidative stress and DNA damage in the aquatic macrophyte Myriophyllumquitense. B Environ. Contam. Toxicol. 2015, 94, 146–151. [Google Scholar] [CrossRef] [PubMed]
- Oliveira, F.A.; Reis, L.P.G.; Soto-Blanco, B.; Melo, M.M. Pesticides residues in the prochiloduscostatus (valenciennes, 1850) fish caught in the Sao Francisco River, Brazil. J. Environ. Sci. Health B 2015, 50, 398–405. [Google Scholar] [CrossRef]
- Jia, W.; Mao, L.; Zhang, L.; Zhang, Y.; Jiang, H. Effects of two strobilurins (azoxystrobin and picoxystrobin) on embryonic development and enzyme activities in juveniles and adult fish livers of zebrafish (Danio rerio). Chemosphere 2018, 207, 573–580. [Google Scholar] [CrossRef]
- Harms, H.; Schlosser, D.; Wick, L.Y. Untapped potential: Exploiting fungi in bioremediation of hazardous chemicals. Nat. Rev. Microbiol. 2011, 9, 177–192. [Google Scholar] [CrossRef]
- Chen, S.; Hu, M.; Liu, J.; Zhong, G.; Yang, L.; Rizwan-ul-Haq, M.; Han, H. Biodegradation of beta-cypermethrin and 3-phenoxybenzoic acid by a novel Ochrobactrumlupini DG-S-01. J. Hazard. Mater. 2011, 187, 433–440. [Google Scholar] [CrossRef]
- Xiong, D.; Gao, Z.Z.; Fu, B.; Sun, H.; Tian, S.L.; Xiao, Y.M.; Qin, Z.H. Effect of pyrimorph on soil enzymatic activities and respiration. Eur. J. Soil Biol. 2013, 56, 44–48. [Google Scholar] [CrossRef]
- Chen, S.; Luo, J.J.; Hu, M.Y.; Lai, K.P.; Geng, P.; Huang, H.S. Enhancement of cypermethrin degradation by a coculture of Bacillus cereus ZH-3 and Streptomyces aureus HP-S-01. Bioresour. Technol. 2012, 110, 97–104. [Google Scholar] [CrossRef] [PubMed]
- Cycoń, M.; Mrozik, A.; Piotrowska-Seget, Z. Bioaugmentation as a strategy for the remediation of pesticide-polluted soil: A review. Chemosphere 2017, 172, 52–71. [Google Scholar] [CrossRef] [PubMed]
- Zhan, H.; Wang, H.; Liao, L.; Feng, Y.; Fan, X.; Zhang, L.; Chen, S. Kinetics and novel degradation pathway of permethrin in Acinetobacterbaumannii ZH-14. Front. Microbiol. 2018, 9, 98. [Google Scholar] [CrossRef] [PubMed]
- Arora, P.K.; Srivastava, A.; Garg, S.K.; Singh, V.P. Recent advances in degradation of chloronitrophenols. Bioresour. Technol. 2017, 250, 902–909. [Google Scholar] [CrossRef] [PubMed]
- Huang, Y.; Zhan, H.; Bhatt, P.; Chen, S. Paraquat degradation from contaminated environments: Current achievements and perspectives. Front. Microbiol. 2019, 10, 1754. [Google Scholar] [CrossRef]
- Zhang, W.; Lin, Z.; Pang, S.; Bhatt, P.; Chen, S. Insights into the biodegradation of lindane (γ-hexachlorocyclohexane) using a microbial system. Front. Microbiol. 2020, 11, 522. [Google Scholar] [CrossRef]
- Lopes, F.M.; Batista, K.A.; Batista, G.L.; Mitidieri, S.; Bataus, L.A.; Fernandes, K.F. Biodegradation of epoxyconazole and piraclostrobin fungicides by Klebsiella sp. from soil. World J. Microbiol. Biotechnol. 2010, 26, 1155–1161. [Google Scholar] [CrossRef]
- Clinton, B.; Warden, A.; Haboury, S.; Easton, C.J.; Kotsonis, S.; Taylor, M.C.; Oakeshott, J.G.; Russell, R.J.; Scott, C. Bacterial degradation of strobilurin fungicides: A role for a promiscuous methyl esterase activity of the subtilisin proteases? Biocatal. Biotransform. 2011, 29, 119–129. [Google Scholar] [CrossRef]
- Howell, C.C.; Semple, K.T.; Bending, G.D. Isolation and characterisation of azoxystrobin degrading bacteria from soil. Chemosphere 2014, 95, 370–378. [Google Scholar] [CrossRef]
- Bacmaga, M.; Kucharski, J.; Wyszkowska, J. Microbial and enzymatic activity of soil contaminated with azoxystrobin. Environ. Monit. Asses. 2015, 187, 615–629. [Google Scholar] [CrossRef] [PubMed]
- Chen, S.; Geng, P.; Xiao, Y.; Hu, M. Bioremediation of β-cypermethrin and 3-phenoxybenzaldehyde contaminated soils using Streptomyces aureus HP-S-01. Appl. Microbiol. Biotechnol. 2012, 94, 505–515. [Google Scholar] [CrossRef] [PubMed]
- Xiao, Y.; Chen, S.; Gao, Y.; Hu, W.; Hu, M.; Zhong, G. Isolation of a novel beta-cypermethrin degrading strain Bacillus subtilis BSF01 and its biodegradation pathway. Appl. Microbiol. Biotechnol. 2015, 99, 2849–2859. [Google Scholar] [CrossRef]
- Chen, S.; Deng, Y.; Chang, C.; Lee, J.; Cheng, Y.; Cui, Z.; Zhou, J.; He, F.; Hu, M.; Zhang, L.H. Pathway and kinetics of cyhalothrin biodegradation by Bacillus thuringiensis strain ZS-19. Sci. Rep. 2015, 5, 8784. [Google Scholar] [CrossRef]
- Ye, T.; Zhou, T.; Li, Q.; Xu, X.; Fan, X.; Zhang, L.; Chen, S. Cupriavidussp. HN-2, a novel quorum quenching bacterial isolate, is a potent biocontrolagent against Xanthomonascampestrispv. campestris. Microorganisms 2020, 8, 45. [Google Scholar] [CrossRef]
- Chen, S.; Dong, Y.H.; Chang, C.; Deng, Y.; Zhang, X.F.; Zhong, G.; Song, H.; Hu, M.; Zhang, L.H. Characterization of a novel cyfluthrin-degrading bacterial strain Brevibacteriumaureum and its biochemical degradation pathway. Bioresour. Technol. 2013, 132, 16–23. [Google Scholar] [CrossRef]
- Tamura, K.; Dudley, J.; Nei, M.; Kumar, S. MEGA4: Molecular Evolutionary Genetics Analysis (MEGA) software version 4.0. Mol. Biol. Evol. 2007, 24, 1596–1599. [Google Scholar] [CrossRef]
- Chen, S.; Hu, Q.B.; Hu, M.Y.; Luo, J.J.; Weng, Q.F.; Lai, K.P. Isolation and characterization of a fungus able to degrade pyrethroids and 3-phenoxybenzaldehyde. Bioresour. Technol. 2011, 102, 8110–8116. [Google Scholar] [CrossRef]
- Zhang, C.; Wang, S.; Yan, Y. Isomerization and biodegradation of beta-cypermethrin by Pseudomonas aeruginosa CH7 with biosurfactant production. Bioresour. Technol. 2011, 102, 7139–7146. [Google Scholar] [CrossRef]
- Yang, J.; Feng, Y.; Zhan, H.; Liu, J.; Yang, F.; Zhang, K.; Zhang, L.; Chen, S. Characterization of a pyrethroid-degrading Pseudomonas fulva strain P31 and biochemical degradation pathway of D-phenothrin. Front. Microbiol. 2018, 9, 1003. [Google Scholar] [CrossRef]
- Bhatt, P.; Huang, Y.; Rene, E.R.; Kumar, A.J.; Chen, S. Mechanism of allethrin biodegradation by a newly isolated Sphingomonastrueperi strain CW3 from wastewater sludge. Bioresour. Technol. 2020, 305, 123074. [Google Scholar] [CrossRef]
- Chen, S.; Chang, C.; Deng, Y.; An, S.; Dong, Y.H.; Zhou, J.; Hu, M.; Zhong, G.; Zhang, L.H. Fenpropathrin biodegradation pathway in Bacillus sp. DG-02 and its potential for bioremediation of pyrethroid-contaminated soils. J. Agric. Food Chem. 2014, 62, 2147–2157. [Google Scholar] [CrossRef]
- Cycoń, M.; Zmijowska, A.; Piotrowska-Seget, Z. Enhancement of deltamethrin degradation by soil bioaugmentation with two different strains of Serratiamarcescens. Int. Environ. Sci. Technol. 2014, 11, 1305–1316. [Google Scholar] [CrossRef]
- Tallur, P.N.; Megadi, V.B.; Ninnekar, H.Z. Biodegradation ofcypermethrin by Micrococcus sp. strain CPN 1. Biodegradation 2008, 19, 77–82. [Google Scholar] [CrossRef] [PubMed]
- Chen, S.; Yang, L.; Hu, M.; Liu, J. Biodegradation of fenvalerate and 3-phenoxybenzoic acid by a novel Stenotrophomonas sp. strain ZS-S-01 and its use in bioremediation of contaminated soils. Appl. Microbiol. Biotechnol. 2011, 90, 755–767. [Google Scholar] [CrossRef] [PubMed]
- Cyconń, M.; Wojcik, M.; Piotrowska-Seget, Z. Biodegradationkinetics of the benzimidazole fungicide thiophanate-methyl by bacteriaisolated from loamy sand soil. Biodegradation 2011, 22, 573–583. [Google Scholar] [CrossRef] [PubMed]
- Chen, S.H.; Luo, J.J.; Hu, M.Y.; Geng, P.; Zhang, Y.B. Microbial detoxificationof bifenthrin by a novel yeast and its potential for contaminatedsoils treatment. PLoS ONE 2012, 7, e30862. [Google Scholar]
- Adetutu, E.M.; Ball, A.S.; Osborn, A.M. Azoxystrobin and soil interactions: Degradation and impact on soil bacterial and fungal communities. J. Appl. Microbiol. 2008, 105, 1777–1790. [Google Scholar] [CrossRef]
- Estève, K.; Poupot, C.; Dabert, P.; Mietton-Peuchot, M.; Milisic, V. A saccharomyces cerevisiae-based bioassay for assessing pesticide toxicity. J. Ind. Microbiol. Biotechnol. 2009, 36, 1529–1534. [Google Scholar] [CrossRef]
- Gao, Y.; Chen, S.; Hu, M.; Hu, Q.; Luo, J.; Li, Y. Purification and characterization of a novel chlorpyrifos hydrolase from Cladosporiumcladosporioides Hu-01. PLoS ONE 2012, 7, e38137. [Google Scholar]
- Zhan, H.; Feng, Y.; Fan, X.; Chen, S. Recent advances in glyphosate biodegradation. Appl. Microbiol. Biotechnol. 2018, 102, 5033–5043. [Google Scholar] [CrossRef]
- Bhatt, P.; Bhatt, K.; Huang, Y.; Ziqiu, L.; Chen, S. Esterase is a powerful tool for the biodegradation of pyrethroid insecticides. Chemosphere 2020, 244, 125507. [Google Scholar] [CrossRef]
- Zhang, C.; Jia, L.; Wang, S.H.; Qu, J.; Xu, L.L.; Shi, H.H.; Yan, Y.C. Biodegradation of beta-cypermethrin by two Serratia spp. with different cell surface hydrophobicity. Bioresour. Technol. 2010, 101, 3423–3429. [Google Scholar] [CrossRef]
- Ghevariya, C.M.; Bhatt, J.K.; Dave, B.P. Enhanced chrysene degradation by halotolerant Achromobacterxylosoxidans using response surface methodology. Bioresour. Technol. 2011, 102, 9668–9674. [Google Scholar] [CrossRef]
- Gopal, M.; Jha, S.K.; Niwas, R.; Mukherjee, I.; Shukla, L. Screening of bacteria fordegradation of pesticides. In Proceedings of the National Conference on Soil Contamination and Biodiversity. Industrial Toxicology Research Centre, Lucknow, India, 8–10 February 2002. [Google Scholar]
- Lin, Q.S.; Chen, S.H.; Hu, M.Y.; Rizwan-ul-Haq, M.; Yang, L.; Li, H. Biodegradation of cypermethrin by a newly isolated actinomycetes HU-S-01 from wastewater sludge. Int. J. Environ. Sci. Technol. 2011, 8, 45–56. [Google Scholar] [CrossRef]
- Jilani, S.; Khan, M.A. Biodegradation of cypermethrin by Pseudomonas in a batch activated sludge process. Int. J. Environ. Sci. Technol. 2006, 3, 371–380. [Google Scholar] [CrossRef]
- Luong, J.H. Generalization of Monod kinetics for analyses of growth data with substrate inhibition. Biotechnol. Bioeng. 1987, 29, 242–248. [Google Scholar] [CrossRef]
- Katagi, T. Behaviour of pesticides in water-sediment systems. Rev. Environ. Contam. Toxicol. 2006, 187, 133–251. [Google Scholar]
- Zhang, J.G.; Ruan, L.L.; Zhao, J.H.; Zhu, G.N. An unexpected hydrolysis product from strobilurin fungicide: Azoxystrobin. Chin. J. Struct. Chem. 2011, 30, 1132–1136. [Google Scholar]
- Bauer, A.; Luetjohann, J.; Hanschen, F.S.; Schreiner, M.; Kuballa, J.; Jantzen, E.; Rohn, S. Identification and characterization of pesticide metabolites in Brassicaspecies by liquid chromatography travelling wave ion mobility quadrupoletime-of-flight mass spectrometry (UPLC-TWIMS-QTOF-MS). Food Chem. 2018, 244, 292–303. [Google Scholar] [CrossRef]
- Chen, X.; He, S.; Liang, Z.; Liu, Q.X.; Yan, H.; Hu, J.; Liu, X. Biodegradation of pyraclostrobin by two microbial communities from Hawaiian soils and metabolic mechanism. J. Hazard. Mater. 2018, 354, 225–230. [Google Scholar] [CrossRef]
- Wang, L.; Zhao, J.; Delgado-Moreno, L.; Cheng, J.; Wang, Y.; Zhang, S.; Ye, Q.; Wang, W. Degradation and metabolic profiling for benzene kresoxim-methyl using carbon-14 tracing. Sci. Total Environ. 2018, 637, 638. [Google Scholar] [CrossRef]
- Cullington, J.E.; Walker, A. Rapid biodegradation of diuron and other phenylurea herbicides by a soil bacterium. Soil Biol. Biochem. 1999, 31, 677–686. [Google Scholar] [CrossRef]
- Batisson, I.; Crouzet, O.; Besse-Hoggan, P.; Sancelme, M.; Mangot, J.F.; Mallet, C.; Bohatier, J. Isolation and characterization of mesotrione-degrading Bacillus sp. from soil. Environ. Pollut. 2009, 157, 1195–1201. [Google Scholar] [CrossRef]
- Wang, B.Z.; Ma, Y.; Zhou, W.Y.; Zheng, J.W.; He, J. Biodegradation of syntheticpyrethroids by Ochrobactrumtritici strain pyd-1. World J. Microbiol. Biotechnol. 2011, 27, 2315–2324. [Google Scholar] [CrossRef]
- Birolli, W.G.; Vacondio, B.; Alvarenga, N.; Seleghim, M.H.R.; Porto, A.L.M. Enantioselective biodegradation of the pyrethroid (±)-lambda-cyhalothrin by marine-derived fungi. Chemosphere 2018, 197, 651–660. [Google Scholar] [CrossRef]
- Cycoń, M.; Piotrowska-Seget, Z. Pyrethroid-degrading microorganisms and their potential for the bioremediation of contaminated soils: A review. Front. Microbiol. 2016, 7, 1463. [Google Scholar] [CrossRef]
- Bhatt, P.; Huang, Y.; Zhan, H.; Chen, S. Insight into microbial applications for the biodegradation of pyrethroid insecticides. Front. Microbiol. 2019, 10, 1778. [Google Scholar] [CrossRef]
- Zhang, F.; Wang, L.; Zhou, L.; Wu, D.; Pan, H.; Pan, C. Residue dynamics of pyraclostrobin in peanut and field soil by QuEChERS and LC-MS/MS. Ecotoxicol. Environ. Saf. 2012, 78, 116–122. [Google Scholar] [CrossRef]
- Malhat, F.; Malhat, F.; Kamel, E.; Saber, A.; Hassan, E.; Youssef, A.; Almaz, M.; Hassan, A.; Fayz, A.S. Residues and dissipation of kresoxim methyl in apple under field condition. Food Chem. 2013, 140, 371–374. [Google Scholar] [CrossRef]
- Paramasivam, M.; Selvi, C.; Deepa, M.; Jayaprakash, S.A.; Chandrasekaran, S. Simultaneous determination of tebuconazole, trifloxystrobin, and its metabolite trifloxystrobin acid residues in gherkin under field conditions. J. Sep. Sci. 2015, 38, 958–964. [Google Scholar] [CrossRef] [PubMed]
- Zhan, H.; Huang, Y.; Lin, Z.; Bhatt, P.; Chen, S. New insights into the microbial degradation and catalytic mechanism of synthetic pyrethroids. Environ. Res. 2020, 182, 109138. [Google Scholar] [CrossRef] [PubMed]
- Feng, Y.M.; Fan, X.H.; Zhan, H.; Chen, S.H. Research progress on ecotoxicity and microbial degradation of strobilurin fungicides. Biotechnol. Bull. 2017, 33, 52–58. [Google Scholar]
- Zhang, Q.; Wang, B.; Cao, Z.; Yu, Y. Plasmid-mediated bioaugmentation for the degradation of chlorpyrifos in soil. J. Hazard. Mater. 2012, 221–222, 178–184. [Google Scholar] [CrossRef]
- Lin, Z.; Zhang, W.; Pang, S.; Huang, Y.; Mishra, S.; Bhatt, P.; Chen, S. Current approaches to and future perspectives on methomyl degradation in contaminated soil/water environments. Molecules 2020, 25, 738. [Google Scholar] [CrossRef]
- Pang, S.; Lin, Z.; Zhang, W.; Mishra, S.; Bhatt, P.; Chen, S. Insights into the microbial degradation and biochemical mechanisms of neonicotinoids. Front. Microbiol. 2020, 11, 868. [Google Scholar]
- Liu, J.; Chen, S.; Ding, J.; Xiao, Y.; Han, H.; Zhong, G. Sugarcane bagasse as support for immobilization of Bacillus pumilus HZ-2 and its use in bioremediation of mesotrione-contaminated soils. Appl. Microbiol. Biotechnol. 2015, 99, 10839–10851. [Google Scholar] [CrossRef]
- Bhatt, P.; Huang, Y.; Zhang, W.; Sharma, A.; Chen, S. Enhanced cypermethrin degradation kinetics and metabolic pathway in Bacillus thuringiensis strain SG4. Microorganims 2020, 8, 223. [Google Scholar] [CrossRef]
- Chen, S.; Hu, W.; Xiao, Y.; Deng, Y.; Jia, J.; Hu, M. Degradation of 3-phenoxybenzoic acid by a Bacillus sp. PLoS ONE 2012, 7, e50456. [Google Scholar] [CrossRef]
- Huang, Y.; Lin, Z.; Zhang, W.; Pang, S.; Bhatt, P.; Rene, E.R.; Kumar, A.J.; Chen, S. New insights into the microbial degradation of D-cyphenothrin in contaminated water/soil environments. Microorganisms 2020, 8, 473. [Google Scholar] [CrossRef]
- Zhai, Y.; Li, K.; Song, J.; Shi, Y.; Yan, Y. Molecular cloning, purification and biochemical characterization of a novel pyrethroid-hydrolyzing carboxylesterase gene from Ochrobactrumanthropi YZ-1. J. Hazard. Mater. 2012, 221–222, 206–212. [Google Scholar] [CrossRef] [PubMed]
- Chudasama, K.S.; Thaker, V.S. Genome sequence of Ochrobactrumanthropi strain SUBG007, a plant pathogen and potential xenobiotic compounds degradation bacterium. Genom. Data 2017, 11, 116–117. [Google Scholar] [CrossRef] [PubMed]
- Chen, H.W.; Xu, M.; Ma, X.W.; Tong, Z.H.; Liu, D.F. Isolation and characterization of a chlorate-reducing bacterium Ochrobactrumanthropi XM-1. J. Hazard. Mater. 2019, 380, 120873. [Google Scholar] [CrossRef] [PubMed]
- Aguilera-Arreola, M.G.; Ostria-Hernández, M.L.; Albarrán-Fernández, E.; Juárez-Enriquez, S.R.; Majalca-Martínez, C.; Rico-Verdín, B.; Ruiz, E.A.; Ruiz-Palma, M.D.; Morales-García, M.R.; Contreras-Rodríguez, A. Correct identification of Ochrobactrumanthropi from blood culture using 16rRNA sequencing: A first case report in an immunocompromised patient in Mexico. Front. Med. 2018, 5, 205. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Characteristics | Results | Characteristics | Results |
|---|---|---|---|
| Glucose | + | Mannitol | − |
| Arginine | − | Oxidase | + |
| Arabinose | + | Urea | − |
| Gelatin | − | Maltose | + |
| Mannose | + | N-acetyl-D-glucosamine | + |
| Esculin | − | Tryptophan | − |
| Decanoic acid | + | Glyconate | − |
| Adipic acid | − | Malic acid | + |
| Citric acid | + | Phenylacetic acid | − |
| p-Nitrophenyl β-D-galactopyranoside | − | Potassium nitrate(KNO3) | + |
| Run | X1 | X2 | X3 | Response (Y1) |
|---|---|---|---|---|
| Degradation (%) | ||||
| 1 | −1 | −1 | −1 | 62.3 |
| 2 | −1 | −1 | +1 | 66.1 |
| 3 | −1 | +1 | −1 | 71.5 |
| 4 | −1 | +1 | +1 | 68.4 |
| 5 | +1 | −1 | −1 | 70.7 |
| 6 | +1 | −1 | +1 | 70.2 |
| 7 | +1 | +1 | −1 | 71.3 |
| 8 | +1 | +1 | +1 | 72.5 |
| 9 | −1.68 | 0 | 0 | 54.5 |
| 10 | +1.68 | 0 | 0 | 57.8 |
| 11 | 0 | −1.68 | 0 | 75.6 |
| 12 | 0 | +1.68 | 0 | 81.9 |
| 13 | 0 | 0 | −1.68 | 80.1 |
| 14 | 0 | 0 | +1.68 | 85.6 |
| 15 | 0 | 0 | 0 | 86.1 |
| 16 | 0 | 0 | 0 | 87.0 |
| 17 | 0 | 0 | 0 | 87.2 |
| 18 | 0 | 0 | 0 | 86.5 |
| 19 | 0 | 0 | 0 | 85.3 |
| 20 | 0 | 0 | 0 | 86.0 |
| 21 | 0 | 0 | 0 | 87.2 |
| 22 | 0 | 0 | 0 | 85.7 |
| 23 | 0 | 0 | 0 | 86.9 |
| Compound | Retention Time (min) | m/z | Chemical Structural Formula in NIST Library | Name |
|---|---|---|---|---|
| A | 22.869 | 403 |  | Azoxystrobin |
| B | 9.126 | 197 |  | N-(4,6-Dimethoxypyrimidin-2-yl)-acetamide |
| C | 14.754 | 257.5 |  | 2-Amino-4-(4-chlorophenyl)-3-cyano-5,6-dimethyl-pyridine |
| D | 15.521 | 253 |  | 3-Quinolinecarboxylic acid,6,8-difluoro-4-hydroxy-, ethyl ester |
| Strobilurins | Regression Equation | k (d−1) | t1/2 (d) | R2 |
|---|---|---|---|---|
| Azoxystrobin | Ct = 50.8745 × e−0.4554t | 0.4554 | 1.52 | 0.9846 |
| Kresoxim-methyl | Ct = 50.9434 × e−0.5161t | 0.5161 | 1.34 | 0.9879 |
| Pyraclostrobin | Ct = 50.3763 × e−0.5153t | 0.5153 | 1.35 | 0.9902 |
| Trifloxystrobin | Ct = 51.1746 × e−0.3845t | 0.3845 | 1.80 | 0.9626 |
| Picoxystrobin | Ct = 50.9921 × e−0.3371t | 0.3371 | 2.06 | 0.9823 |
| Fluoxastrobin | Ct = 53.0503 × e−0.1857t | 0.1857 | 3.73 | 0.9419 |
| Soil Treatments | Regression Equation | k (d−1) | t1/2 (d) | R2 |
|---|---|---|---|---|
| SS + azoxystrobin | Ct = 19.9387 × e−0.0064t | 0.0064 | 108.3 | 0.9447 |
| nSS + azoxystrobin | Ct = 19.9796 × e−0.0092t | 0.0092 | 75.3 | 0.9892 |
| SS + azoxystrobin+ SH14 | Ct = 20.7399 × e−0.0550t | 0.0550 | 12.6 | 0.9824 |
| nSS + azoxystrobin+ SH14 | Ct = 20.9485 × e−0.0715t | 0.0715 | 9.7 | 0.9597 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Feng, Y.; Zhang, W.; Pang, S.; Lin, Z.; Zhang, Y.; Huang, Y.; Bhatt, P.; Chen, S. Kinetics and New Mechanism of Azoxystrobin Biodegradation by an Ochrobactrum anthropi Strain SH14. Microorganisms 2020, 8, 625. https://doi.org/10.3390/microorganisms8050625
Feng Y, Zhang W, Pang S, Lin Z, Zhang Y, Huang Y, Bhatt P, Chen S. Kinetics and New Mechanism of Azoxystrobin Biodegradation by an Ochrobactrum anthropi Strain SH14. Microorganisms. 2020; 8(5):625. https://doi.org/10.3390/microorganisms8050625
Chicago/Turabian StyleFeng, Yanmei, Wenping Zhang, Shimei Pang, Ziqiu Lin, Yuming Zhang, Yaohua Huang, Pankaj Bhatt, and Shaohua Chen. 2020. "Kinetics and New Mechanism of Azoxystrobin Biodegradation by an Ochrobactrum anthropi Strain SH14" Microorganisms 8, no. 5: 625. https://doi.org/10.3390/microorganisms8050625
APA StyleFeng, Y., Zhang, W., Pang, S., Lin, Z., Zhang, Y., Huang, Y., Bhatt, P., & Chen, S. (2020). Kinetics and New Mechanism of Azoxystrobin Biodegradation by an Ochrobactrum anthropi Strain SH14. Microorganisms, 8(5), 625. https://doi.org/10.3390/microorganisms8050625