The Role of Oxidoreductase-Like Protein Olp1 in Sexual Reproduction and Virulence of Cryptococcus neoformans



Abstract
1. Introduction
2. Materials and Methods
2.1. Ethics Statement
2.2. Strains, Media, and Growth Conditions
2.3. OLP1 Gene Expression Pattern Analysis
2.4. Detection of OLP1 Gene Expression Using qRT-PCR
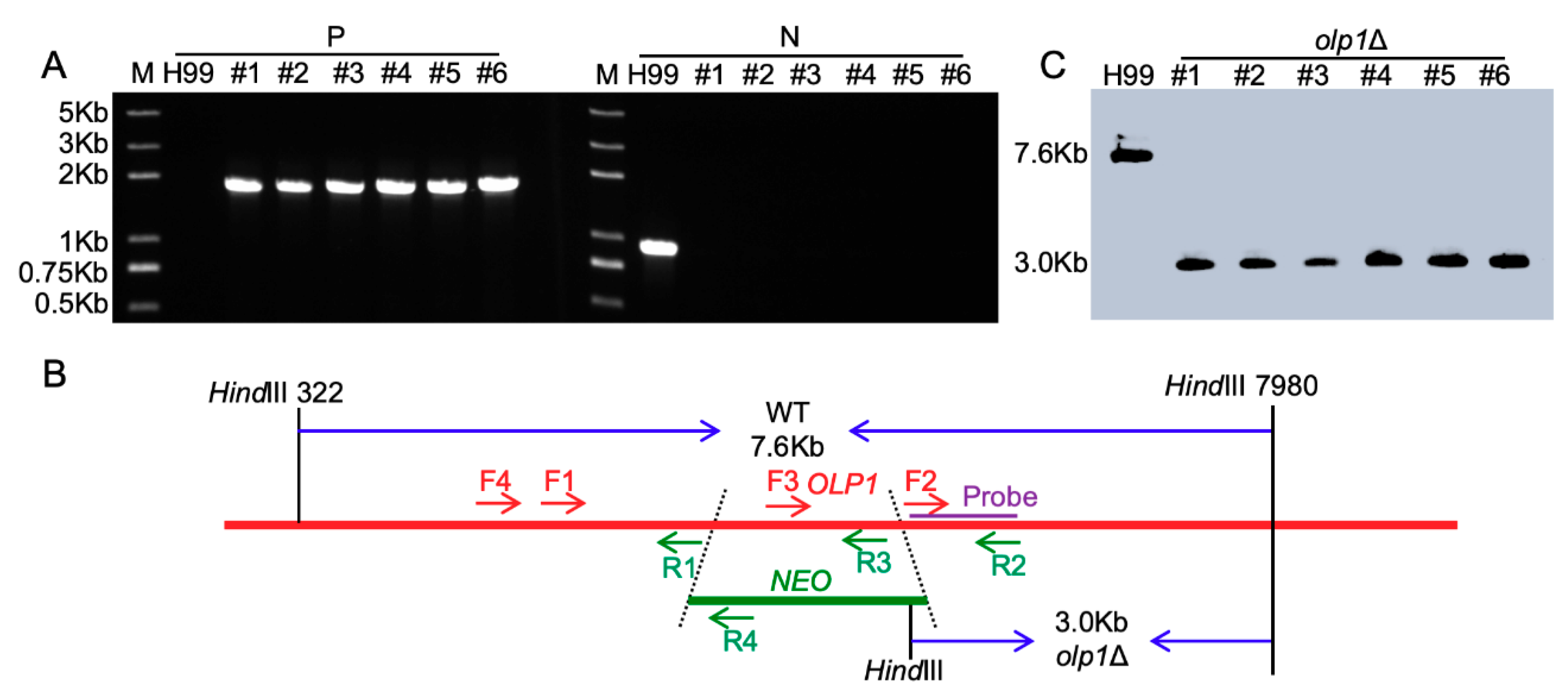
2.5. Generation of OLP1 Gene Knockout, Complementation, and Overexpression Strains
2.6. Generation of Nop1-mCherry Strains
2.7. Assays for Melanin, Capsule Production, and Mating
2.8. Virulence Studies
2.9. Histopathology and Fungal Burdens in Infected Organs
3. Results
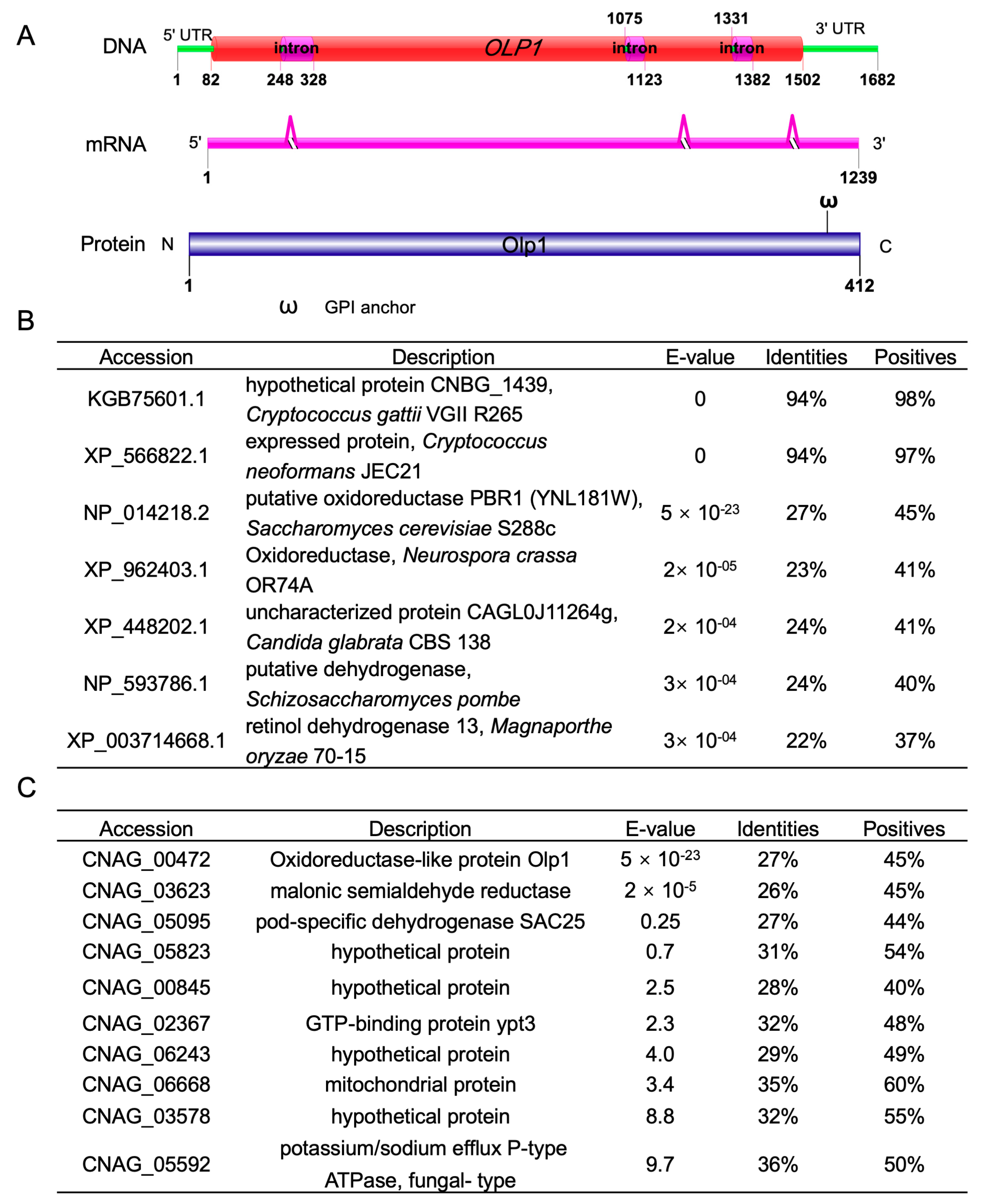
3.1. Identification of the Oxidoreductases-Like Protein Olp1 in C. neoformans
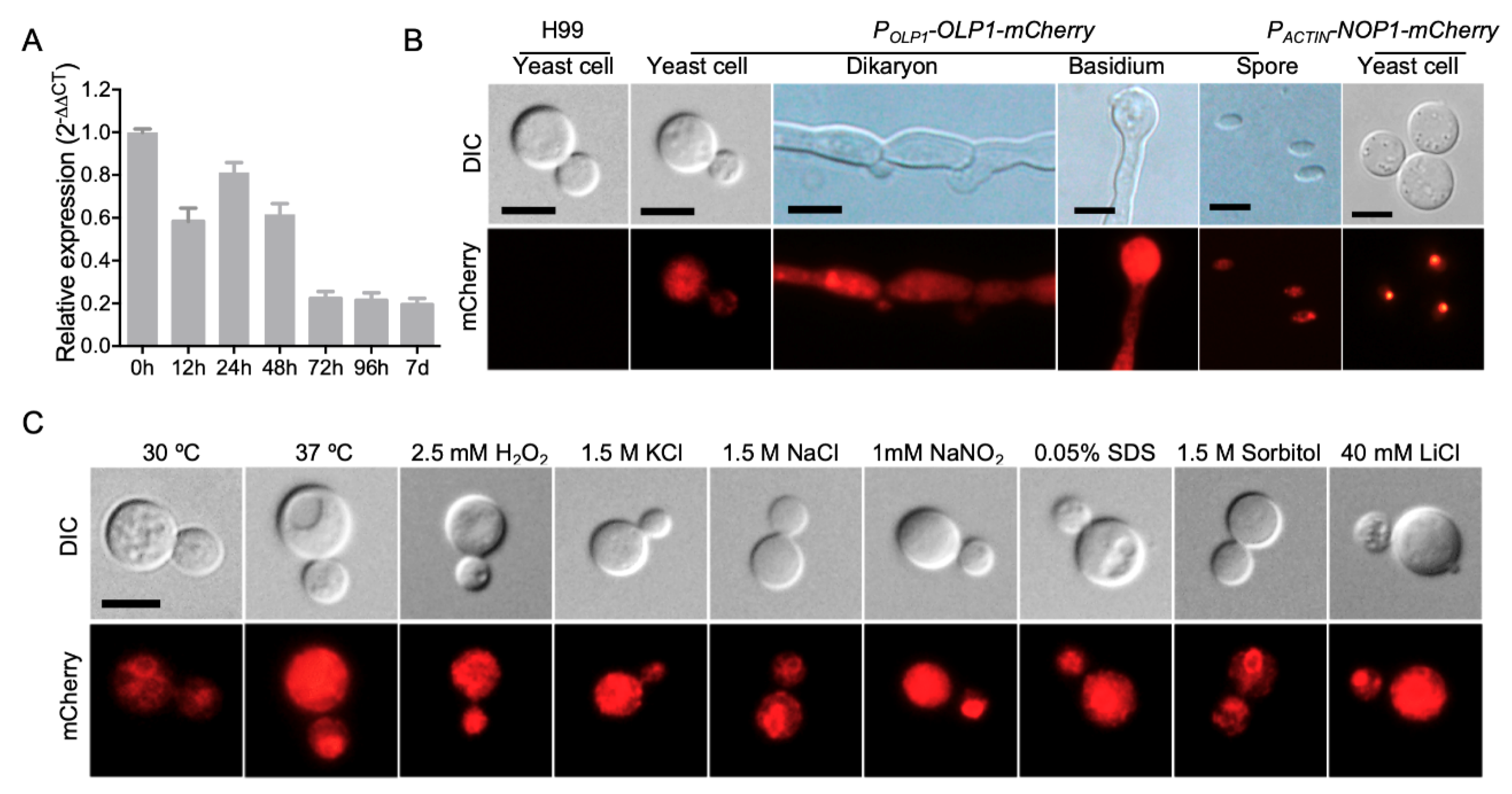
3.2. Expression of the OLP1 Gene in C. neoformans
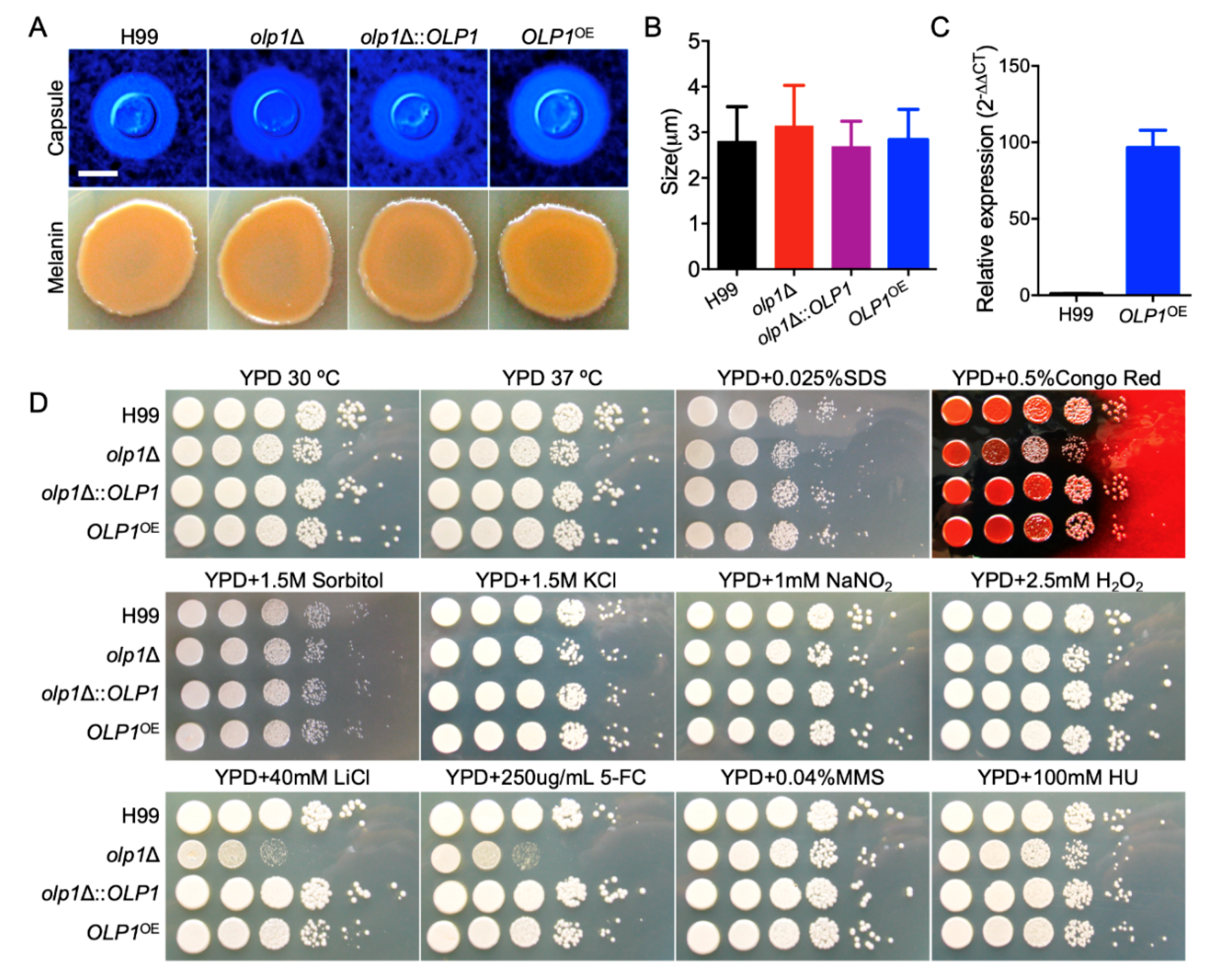
3.3. Olp1 Plays Roles in Stress Responses
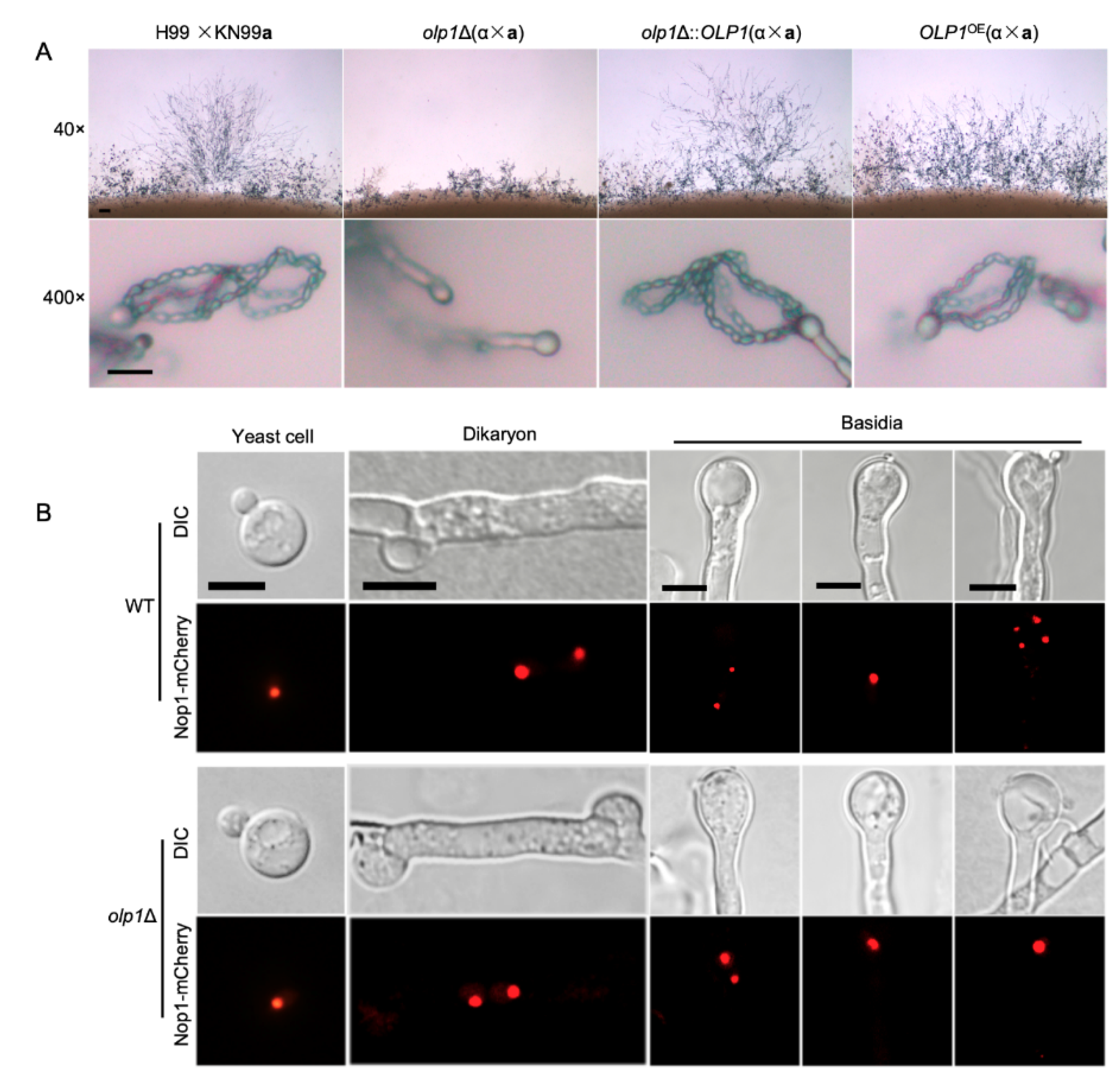
3.4. Olp1 is Crucial for Sexual Sporulation
3.5. Olp1 Is Involved in Meiosis and Nuclear Division
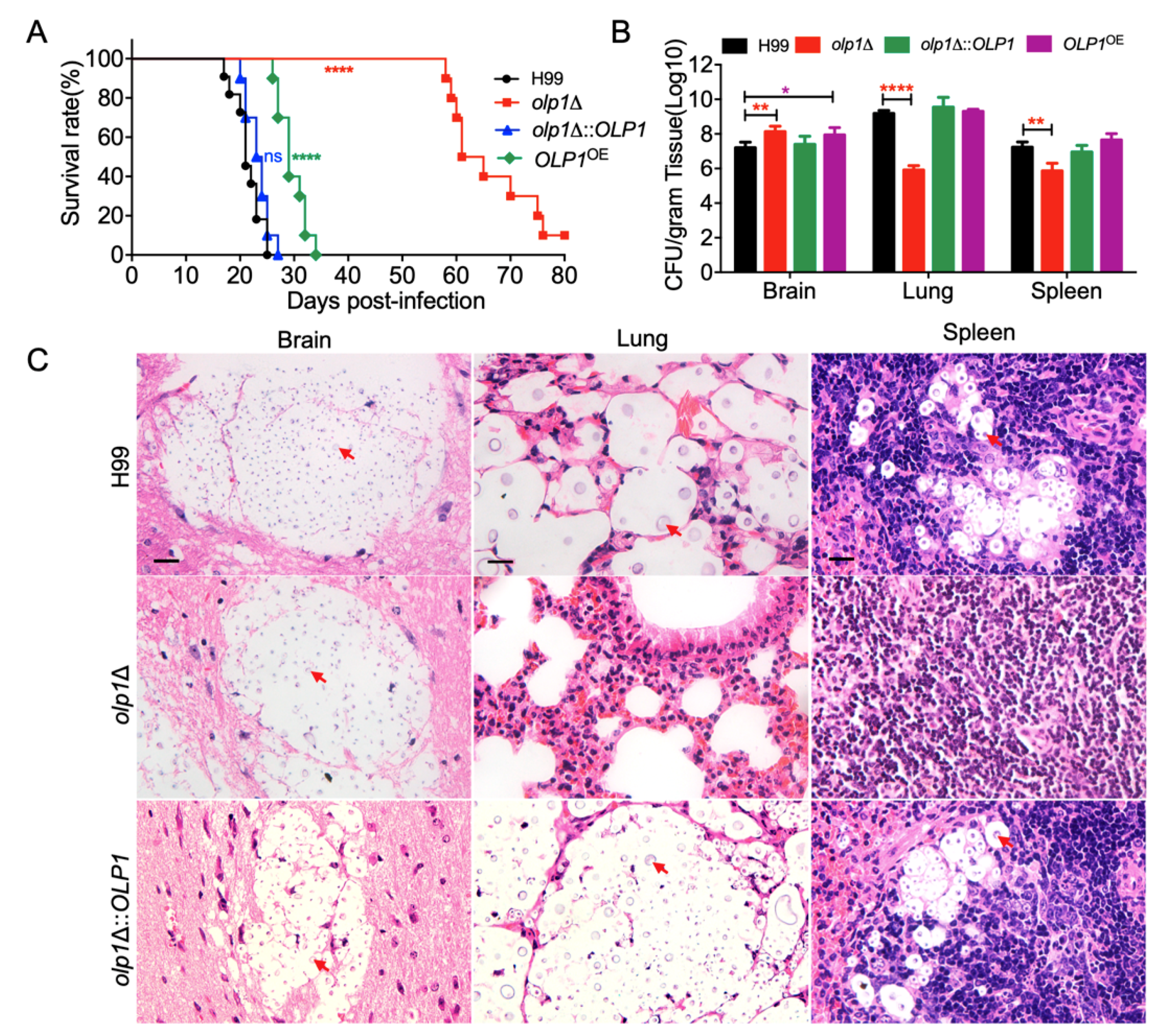
3.6. Olp1 Is Required for Fungal Virulence
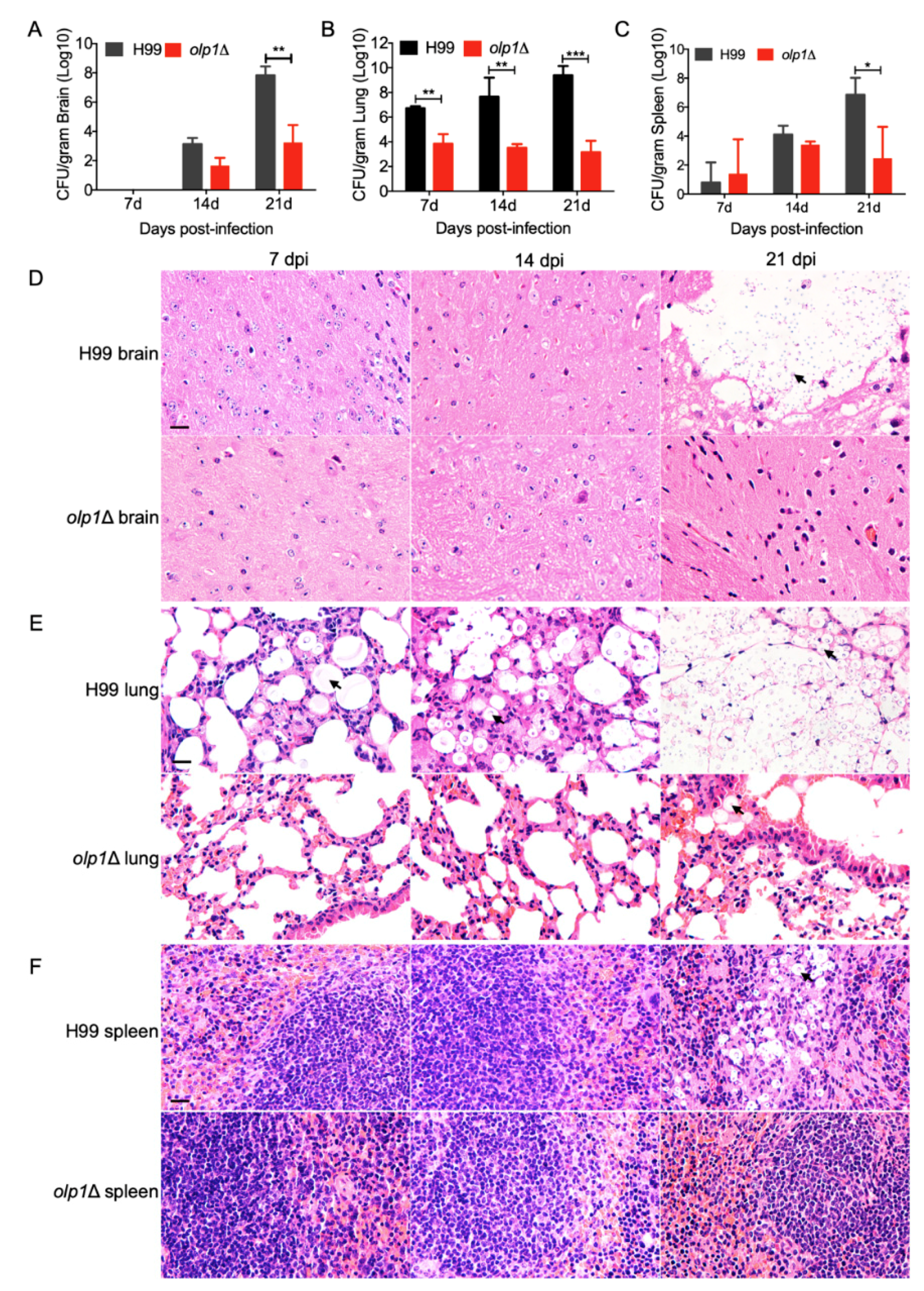
3.7. Olp1 Is Vital for Fungal Infection Progression
4. Discussion
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- May, R.C.; Stone, N.R.; Wiesner, D.L.; Bicanic, T.; Nielsen, K. Cryptococcus: From environmental saprophyte to global pathogen. Nat. Rev. Microbiol. 2016, 14, 106–117. [Google Scholar] [CrossRef]
- Park, B.J.; Wannemuehler, K.A.; Marston, B.J.; Govender, N.; Pappas, P.G.; Chiller, T.M. Estimation of the current global burden of cryptococcal meningitis among persons living with HIV/AIDS. AIDS 2009, 23, 525–530. [Google Scholar] [CrossRef]
- Rajasingham, R.; Smith, R.M.; Park, B.J.; Jarvis, J.N.; Govender, N.P.; Chiller, T.M.; Denning, D.W.; Loyse, A.; Boulware, D.R. Global burden of disease of HIV-associated cryptococcal meningitis: An updated analysis. Lancet Infect. Dis. 2017, 17, 873–881. [Google Scholar] [CrossRef]
- Xu, J.; Vilgalys, R.; Mitchell, T.G. Multiple gene genealogies reveal recent dispersion and hybridization in the human pathogenic fungus Cryptococcus neoformans. Mol. Ecol. 2000, 9, 1471–1481. [Google Scholar] [CrossRef]
- Kwon-Chung, K.J.; Boekhout, T.; Fell, J.W.; Diaz, M. (1557) Proposal to Conserve the Name Cryptococcus gattii against C. hondurianus and C. bacillisporus (Basidiomycota, Hymenomycetes, Tremellomycetidae). Taxon 2002, 51, 804–806. [Google Scholar] [CrossRef]
- Hagen, F.; Khayhan, K.; Theelen, B.; Kolecka, A.; Polacheck, I.; Sionov, E.; Falk, R.; Parnmen, S.; Lumbsch, H.T.; Boekhout, T. Recognition of seven species in the Cryptococcus gattii/Cryptococcus neoformans species complex. Fungal Genet. Biol. 2015, 78, 16–48. [Google Scholar] [CrossRef]
- Kwon-Chung, K.J.; Bennett, J.E.; Wickes, B.L.; Meyer, W.; Cuomo, C.A.; Wollenburg, K.R.; Bicanic, T.A.; Castaneda, E.; Chang, Y.C.; Chen, J.H.; et al. The Case for Adopting the “Species Complex” Nomenclature for the Etiologic Agents of Cryptococcosis. mSphere 2017, 2. [Google Scholar] [CrossRef]
- Zhao, Y.; Lin, J.; Fan, Y.; Lin, X. Life Cycle of Cryptococcus neoformans. Annu. Rev. Microbiol. 2019, 73, 17–42. [Google Scholar] [CrossRef]
- Lin, X.; Heitman, J. The biology of the Cryptococcus neoformans species complex. Annu. Rev. Microbiol. 2006, 60, 69–105. [Google Scholar] [CrossRef]
- Lin, X.; Hull, C.M.; Heitman, J. Sexual reproduction between partners of the same mating type in Cryptococcus neoformans. Nature 2005, 434, 1017–1021. [Google Scholar] [CrossRef] [PubMed]
- Kozel, T.R. Virulence factors of Cryptococcus neoformans. Trends Microbiol. 1995, 3, 295–299. [Google Scholar] [CrossRef]
- Kronstad, J.; Jung, W.H.; Hu, G. Beyond the big three: Systematic analysis of virulence factors in Cryptococcus neoformans. Cell Host Microbe 2008, 4, 308–310. [Google Scholar] [CrossRef] [PubMed]
- Cox, G.M.; Mukherjee, J.; Cole, G.T.; Casadevall, A.; Perfect, J.R. Urease as a virulence factor in experimental cryptococcosis. Infect. Immun. 2000, 68, 443–448. [Google Scholar] [CrossRef]
- Cox, G.M.; McDade, H.C.; Chen, S.C.; Tucker, S.C.; Gottfredsson, M.; Wright, L.C.; Sorrell, T.C.; Leidich, S.D.; Casadevall, A.; Ghannoum, M.A.; et al. Extracellular phospholipase activity is a virulence factor for Cryptococcus neoformans. Mol. Microbiol. 2001, 39, 166–175. [Google Scholar] [CrossRef]
- Chaturvedi, V.; Wong, B.; Newman, S.L. Oxidative killing of Cryptococcus neoformans by human neutrophils. Evidence that fungal mannitol protects by scavenging reactive oxygen intermediates. J. Immunol. 1996, 156, 3836–3840. [Google Scholar]
- Chaturvedi, V.; Flynn, T.; Niehaus, W.G.; Wong, B. Stress tolerance and pathogenic potential of a mannitol mutant of Cryptococcus neoformans. Microbiology 1996, 142 Pt 4, 937–943. [Google Scholar] [CrossRef]
- Qiu, Y.; Davis, M.J.; Dayrit, J.K.; Hadd, Z.; Meister, D.L.; Osterholzer, J.J.; Williamson, P.R.; Olszewski, M.A. Immune modulation mediated by cryptococcal laccase promotes pulmonary growth and brain dissemination of virulent Cryptococcus neoformans in mice. PLoS ONE 2012, 7, e47853. [Google Scholar] [CrossRef] [PubMed]
- Jong, A.; Wu, C.H.; Gonzales-Gomez, I.; Kwon-Chung, K.J.; Chang, Y.C.; Tseng, H.K.; Cho, W.L.; Huang, S.H. Hyaluronic acid receptor CD44 deficiency is associated with decreased Cryptococcus neoformans brain infection. J. Biol. Chem. 2012, 287, 15298–15306. [Google Scholar] [CrossRef]
- Odom, A.; Muir, S.; Lim, E.; Toffaletti, D.L.; Perfect, J.; Heitman, J. Calcineurin is required for virulence of Cryptococcus neoformans. EMBO J. 1997, 16, 2576–2589. [Google Scholar] [CrossRef]
- Vu, K.; Tham, R.; Uhrig, J.P.; Thompson, G.R., 3rd; Na Pombejra, S.; Jamklang, M.; Bautos, J.M.; Gelli, A. Invasion of the central nervous system by Cryptococcus neoformans requires a secreted fungal metalloprotease. MBio 2014, 5, e01101-14. [Google Scholar] [CrossRef] [PubMed]
- Na Pombejra, S.; Salemi, M.; Phinney, B.S.; Gelli, A. The Metalloprotease, Mpr1, Engages AnnexinA2 to Promote the Transcytosis of Fungal Cells across the Blood-Brain Barrier. Front. Cell. Infect. Microbiol. 2017, 7, 296. [Google Scholar] [CrossRef]
- May, S.W.; Padgette, S.R. Oxidoreductase Enzymes in Biotechnology: Current Status and Future Potential. Bio/Technology 1983, 1, 677. [Google Scholar] [CrossRef]
- King, A.; Selak, M.A.; Gottlieb, E. Succinate dehydrogenase and fumarate hydratase: Linking mitochondrial dysfunction and cancer. Oncogene 2006, 25, 4675–4682. [Google Scholar] [CrossRef]
- Gottlieb, E.; Tomlinson, I.P. Mitochondrial tumour suppressors: A genetic and biochemical update. Nat. Rev. Cancer 2005, 5, 857–866. [Google Scholar] [CrossRef]
- Battelli, M.G.; Polito, L.; Bortolotti, M.; Bolognesi, A. Xanthine oxidoreductase in cancer: More than a differentiation marker. Cancer Med. 2016, 5, 546–557. [Google Scholar] [CrossRef] [PubMed]
- Shen, J.; Barrios, R.J.; Jaiswal, A.K. Inactivation of the quinone oxidoreductases NQO1 and NQO2 strongly elevates the incidence and multiplicity of chemically induced skin tumors. Cancer Res. 2010, 70, 1006–1014. [Google Scholar] [CrossRef]
- Grunblatt, E.; Riederer, P. Aldehyde dehydrogenase (ALDH) in Alzheimer’s and Parkinson’s disease. J. Neural Transm. (Vienna) 2016, 123, 83–90. [Google Scholar] [CrossRef]
- Marchitti, S.A.; Brocker, C.; Stagos, D.; Vasiliou, V. Non-P450 aldehyde oxidizing enzymes: The aldehyde dehydrogenase superfamily. Expert Opin. Drug Metab. Toxicol. 2008, 4, 697–720. [Google Scholar] [CrossRef]
- Suzuki, H.; DeLano, F.A.; Parks, D.A.; Jamshidi, N.; Granger, D.N.; Ishii, H.; Suematsu, M.; Zweifach, B.W.; Schmid-Schonbein, G.W. Xanthine oxidase activity associated with arterial blood pressure in spontaneously hypertensive rats. Proc. Natl. Acad. Sci. USA 1998, 95, 4754–4759. [Google Scholar] [CrossRef]
- Berry, C.E.; Hare, J.M. Xanthine oxidoreductase and cardiovascular disease: Molecular mechanisms and pathophysiological implications. J. Physiol. 2004, 555, 589–606. [Google Scholar] [CrossRef]
- Zhang, Z.; Wang, J.; Chai, R.; Qiu, H.; Jiang, H.; Mao, X.; Wang, Y.; Liu, F.; Sun, G. An S-(hydroxymethyl)glutathione dehydrogenase is involved in conidiation and full virulence in the rice blast fungus Magnaporthe oryzae. PLoS ONE 2015, 10, e0120627. [Google Scholar] [CrossRef]
- Pedrini, N.; Ortiz-Urquiza, A.; Huarte-Bonnet, C.; Fan, Y.; Juarez, M.P.; Keyhani, N.O. Tenebrionid secretions and a fungal benzoquinone oxidoreductase form competing components of an arms race between a host and pathogen. Proc. Natl. Acad. Sci. USA 2015, 112, E3651–E3660. [Google Scholar] [CrossRef]
- Fang, W.; Fa, Z.; Liao, W. Epidemiology of Cryptococcus and cryptococcosis in China. Fungal Genet. Biol. 2015, 78, 7–15. [Google Scholar] [CrossRef]
- Grahl, N.; Puttikamonkul, S.; Macdonald, J.M.; Gamcsik, M.P.; Ngo, L.Y.; Hohl, T.M.; Cramer, R.A. In Vivo hypoxia and a fungal alcohol dehydrogenase influence the pathogenesis of invasive pulmonary aspergillosis. PLoS Pathog. 2011, 7, e1002145. [Google Scholar] [CrossRef]
- Akhter, S.; McDade, H.C.; Gorlach, J.M.; Heinrich, G.; Cox, G.M.; Perfect, J.R. Role of alternative oxidase gene in pathogenesis of Cryptococcus neoformans. Infect. Immun. 2003, 71, 5794–5802. [Google Scholar] [CrossRef]
- Giles, S.S.; Perfect, J.R.; Cox, G.M. Cytochrome c peroxidase contributes to the antioxidant defense of Cryptococcus neoformans. Fungal Genet. Biol. 2005, 42, 20–29. [Google Scholar] [CrossRef]
- Suvarna, K.; Bartiss, A.; Wong, B. Mannitol-1-phosphate dehydrogenase from Cryptococcus neoformans is a zinc-containing long-chain alcohol/polyol dehydrogenase. Microbiology 2000, 146 Pt 10, 2705–2713. [Google Scholar] [CrossRef]
- Missall, T.A.; Pusateri, M.E.; Lodge, J.K. Thiol peroxidase is critical for virulence and resistance to nitric oxide and peroxide in the fungal pathogen, Cryptococcus neoformans. Mol. Microbiol. 2004, 51, 1447–1458. [Google Scholar] [CrossRef]
- Missall, T.A.; Lodge, J.K. Thioredoxin reductase is essential for viability in the fungal pathogen Cryptococcus neoformans. Eukaryot Cell 2005, 4, 487–489. [Google Scholar] [CrossRef]
- Almeida, F.; Wolf, J.M.; Casadevall, A. Virulence-Associated Enzymes of Cryptococcus neoformans. Eukaryot Cell 2015, 14, 1173–1185. [Google Scholar] [CrossRef]
- Xue, C.Y.; Tada, Y.; Dong, X.N.; Heitman, J. The human fungal pathogen Cryptococcus can complete its sexual cycle during a pathogenic association with plants. Cell Host Microbe 2007, 1, 263–273. [Google Scholar] [CrossRef]
- Liu, T.B.; Wang, Y.; Stukes, S.; Chen, Q.; Casadevall, A.; Xue, C. The F-Box protein Fbp1 regulates sexual reproduction and virulence in Cryptococcus neoformans. Eukaryot Cell 2011, 10, 791–802. [Google Scholar] [CrossRef]
- Perfect, J.R.; Ketabchi, N.; Cox, G.M.; Ingram, C.W.; Beiser, C.L. Karyotyping of Cryptococcus neoformans as an epidemiological tool. J. Clin. Microbiol. 1993, 31, 3305–3309. [Google Scholar] [CrossRef] [PubMed]
- Nielsen, K.; Cox, G.M.; Wang, P.; Toffaletti, D.L.; Perfect, J.R.; Heitman, J. Sexual cycle of Cryptococcus neoformans var. grubii and virulence of congenic a and alpha isolates. Infect. Immun. 2003, 71, 4831–4841. [Google Scholar] [PubMed]
- Fan, C.L.; Han, L.T.; Jiang, S.T.; Chang, A.N.; Zhou, Z.Y.; Liu, T.B. The Cys2His2 zinc finger protein Zfp1 regulates sexual reproduction and virulence in Cryptococcus neoformans. Fungal Genet. Biol. 2019, 124, 59–72. [Google Scholar] [CrossRef]
- Fraser, J.A.; Subaran, R.L.; Nichols, C.B.; Heitman, J. Recapitulation of the sexual cycle of the primary fungal pathogen Cryptococcus neoformans var. gattii: Implications for an outbreak on Vancouver Island, Canada. Eukaryot Cell 2003, 2, 1036–1045. [Google Scholar] [CrossRef]
- Kim, M.S.; Kim, S.Y.; Yoon, J.K.; Lee, Y.W.; Bahn, Y.S. An efficient gene-disruption method in Cryptococcus neoformans by double-joint PCR with NAT-split markers. Biochem. Biophys. Res. Commun. 2009, 390, 983–988. [Google Scholar] [CrossRef]
- Basenko, E.Y.; Pulman, J.A.; Shanmugasundram, A.; Harb, O.S.; Crouch, K.; Starns, D.; Warrenfeltz, S.; Aurrecoechea, C.; Stoeckert, C.J., Jr.; Kissinger, J.C.; et al. FungiDB: An Integrated Bioinformatic Resource for Fungi and Oomycetes. J. Fungi 2018, 4, 39. [Google Scholar] [CrossRef] [PubMed]
- Lee, S.C.; Heitman, J. Function of Cryptococcus neoformans KAR7 (SEC66) in karyogamy during unisexual and opposite-sex mating. Eukaryot Cell 2012, 11, 783–794. [Google Scholar] [CrossRef]
- Berridge, M.J.; Downes, C.P.; Hanley, M.R. Neural and developmental actions of lithium: A unifying hypothesis. Cell 1989, 59, 411–419. [Google Scholar] [CrossRef]
- Spiegelberg, B.D.; Dela Cruz, J.; Law, T.H.; York, J.D. Alteration of lithium pharmacology through manipulation of phosphoadenosine phosphate metabolism. J. Biol. Chem. 2005, 280, 5400–5405. [Google Scholar] [CrossRef]
- Stambolic, V.; Ruel, L.; Woodgett, J.R. Lithium inhibits glycogen synthase kinase-3 activity and mimics wingless signalling in intact cells. Curr. Biol. 1996, 6, 1664–1668. [Google Scholar] [CrossRef]
- Dichtl, B.; Stevens, A.; Tollervey, D. Lithium toxicity in yeast is due to the inhibition of RNA processing enzymes. EMBO J. 1997, 16, 7184–7195. [Google Scholar] [CrossRef]
- Murguia, J.R.; Belles, J.M.; Serrano, R. The yeast HAL2 nucleotidase is an in vivo target of salt toxicity. J. Biol. Chem. 1996, 271, 29029–29033. [Google Scholar] [CrossRef]
- Zhao, J.; Lin, W.; Ma, X.; Lu, Q.; Ma, X.; Bian, G.; Jiang, L. The protein kinase Hal5p is the high-copy suppressor of lithium-sensitive mutations of genes involved in the sporulation and meiosis as well as the ergosterol biosynthesis in Saccharomyces cerevisiae. Genomics 2010, 95, 290–298. [Google Scholar] [CrossRef] [PubMed]
- Vermes, A.; Guchelaar, H.J.; Dankert, J. Flucytosine: A review of its pharmacology, clinical indications, pharmacokinetics, toxicity and drug interactions. J. Antimicrob. Chemother. 2000, 46, 171–179. [Google Scholar] [CrossRef]
- Ghoshal, K.; Jacob, S.T. Specific inhibition of pre-ribosomal RNA processing in extracts from the lymphosarcoma cells treated with 5-fluorouracil. Cancer Res. 1994, 54, 632–636. [Google Scholar] [PubMed]
- Santi, D.V.; Hardy, L.W. Catalytic mechanism and inhibition of tRNA (uracil-5-)methyltransferase: Evidence for covalent catalysis. Biochemistry 1987, 26, 8599–8606. [Google Scholar] [CrossRef]
- Patton, J.R. Ribonucleoprotein particle assembly and modification of U2 small nuclear RNA containing 5-fluorouridine. Biochemistry 1993, 32, 8939–8944. [Google Scholar] [CrossRef]
- Yarbro, J.W. Mechanism of action of hydroxyurea. Semin. Oncol. 1992, 19, 1–10. [Google Scholar] [PubMed]
- Lundin, C.; North, M.; Erixon, K.; Walters, K.; Jenssen, D.; Goldman, A.S.; Helleday, T. Methyl methanesulfonate (MMS) produces heat-labile DNA damage but no detectable in vivo DNA double-strand breaks. Nucleic Acids Res. 2005, 33, 3799–3811. [Google Scholar] [CrossRef]
- Costanzo, M.; VanderSluis, B.; Koch, E.N.; Baryshnikova, A.; Pons, C.; Tan, G.; Wang, W.; Usaj, M.; Hanchard, J.; Lee, S.D.; et al. A global genetic interaction network maps a wiring diagram of cellular function. Science 2016, 353. [Google Scholar] [CrossRef] [PubMed]
- Giaever, G.; Chu, A.M.; Ni, L.; Connelly, C.; Riles, L.; Veronneau, S.; Dow, S.; Lucau-Danila, A.; Anderson, K.; Andre, B.; et al. Functional profiling of the Saccharomyces cerevisiae genome. Nature 2002, 418, 387–391. [Google Scholar] [CrossRef]
- Sanchez, R.; Sali, A. Large-scale protein structure modeling of the Saccharomyces cerevisiae genome. Proc. Natl. Acad. Sci. USA 1998, 95, 13597–13602. [Google Scholar] [CrossRef]
- Dahlberg, K.R.; Etten, J.L.V. Physiology and Biochemistry of Fungal Sporulation. Annu. Rev. Phytopathol. 1982, 20, 281–301. [Google Scholar] [CrossRef]
- Huang, M.; Hull, C.M. Sporulation: How to survive on planet Earth (and beyond). Curr. Genet. 2017, 63, 831–838. [Google Scholar] [CrossRef] [PubMed]
- Lara-Ortiz, T.; Riveros-Rosas, H.; Aguirre, J. Reactive oxygen species generated by microbial NADPH oxidase NoxA regulate sexual development in Aspergillus nidulans. Mol. Microbiol. 2003, 50, 1241–1255. [Google Scholar] [CrossRef]
- Cano-Dominguez, N.; Alvarez-Delfin, K.; Hansberg, W.; Aguirre, J. NADPH oxidases NOX-1 and NOX-2 require the regulatory subunit NOR-1 to control cell differentiation and growth in Neurospora crassa. Eukaryot Cell 2008, 7, 1352–1361. [Google Scholar] [CrossRef]
- Zhang, C.; Lin, Y.; Wang, J.; Wang, Y.; Chen, M.; Norvienyeku, J.; Li, G.; Yu, W.; Wang, Z. FgNoxR, a regulatory subunit of NADPH oxidases, is required for female fertility and pathogenicity in Fusarium graminearum. FEMS Microbiol. Lett. 2016, 363, fnv223. [Google Scholar] [CrossRef][Green Version]
- Fisher, S.K.; Novak, J.E.; Agranoff, B.W. Inositol and higher inositol phosphates in neural tissues: Homeostasis, metabolism and functional significance. J. Neurochem. 2002, 82, 736–754. [Google Scholar] [CrossRef]
- Barnett, J.A. The utilization of sugars by yeasts. Adv. Carbohydr. Chem. Biochem. 1976, 32, 125–234. [Google Scholar] [CrossRef]
- Healy, M.E.; Dillavou, C.L.; Taylor, G.E. Diagnostic medium containing inositol, urea, and caffeic acid for selective growth of Cryptococcus neoformans. J. Clin. Microbiol. 1977, 6, 387–391. [Google Scholar]
- Liu, T.B.; Kim, J.C.; Wang, Y.; Toffaletti, D.L.; Eugenin, E.; Perfect, J.R.; Kim, K.J.; Xue, C. Brain inositol is a novel stimulator for promoting Cryptococcus penetration of the blood-brain barrier. PLoS Pathog. 2013, 9, e1003247. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Strains/Plasmids | Genotype/Properties | Source/Reference |
|---|---|---|
| C. neoformans | ||
| H99 | MATα | [43] |
| KN99a | MATa | [44] |
| TBL101 | MATα PACTIN-Nop1-mCherry::NAT | [45] |
| TBL102 | MATaPACTIN-Nop1-mCherry::NAT | [45] |
| TBL337 | MATα olp1::NEO | In this study |
| TBL348 | MATaolp1::NEO | In this study |
| TBL349 | MATα olp1::NEO OLP1::NAT | In this study |
| TBL352 | MATa olp1::NEO OLP1:: NAT | In this study |
| TBL353 | MATα olp1Δ::NEO PACT1N-OLP1-mCherry::NAT | In this study |
| TBL354 | MATaolp1Δ::NEO PACT1N-OLP1-mCherry::NAT | In this study |
| TBL371 | MATα olp1Δ::NEO PACTIN-Nop1-mCherry::NAT | In this study |
| TBL372 | MATaolp1Δ::NEO PACTIN-Nop1-mCherry::NAT | In this study |
| TBL373 | MATα olp1Δ::NEO POLP1-OLP1-mCherry::NAT | In this study |
| TBL374 | MATaolp1Δ::NEO POLP1-OLP1-mCherry::NAT | In this study |
| Plasmids | ||
| pJAF1 | Ampr Plasmid harboring NEO marker | [46] |
| pTBL1 | Ampr Plasmid harboring NAT marke | [45] |
| pTBL3 | Ampr Plasmid harboring mCherry-GPD1 terminator and NAT marker | [45] |
| pTBL5 | Ampr Plasmid harboring mCherry-GPD1 terminator and NAT marker under ACTIN promoter | [45] |
| pTBL68 | Ampr Vector for PACTIN-NOP1-mCherry-NAT for nuclear positioning | [45] |
| pTBL85 | Ampr Vector for PACTIN-mCherry-OLP1 for Olp1 localization | In this study |
| pTBL118 | Ampr Vector for POLP1-OLP1-mCherry for Olp1 localization | In this study |
| pTBL205 | Ampr Vector for POLP1-OLP1-NAT for complementation | In this study |
| Primers | Targeted Genes | Sequence (5′-3′) |
|---|---|---|
| TL17 | M13F | GTAAAACGACGGCCAG |
| TL18 | M13R | CAGGAAACAGCTATGAC |
| TL19 | NEO split F | GGGCGCCCGGTTCTTTTTGTCA |
| TL20 | NEO split R | TTGGTGGTCGAATGGGCAGGTAGC |
| TL59 | NEO R4 | TGTGGATGCTGGCGGAGGATA |
| TL67 | STE20A α F | CCAAAAGCTGATGCTGTGGA |
| TL68 | STE20A α R | AGGACATCTATAGCAGAT |
| TL69 | STE20Aa F | TCCACTGGCAACCCTGCGAG |
| TL70 | STE20Aa R | ATCAGAGACAGAGGAGCAAGAC |
| TL217 | GAPDH qRT-PCR F | TGAGAAGGACCCTGCCAACA |
| TL218 | GAPDH qRT-PCR R | ACTCCGGCTTGTAGGCATCAA |
| TL404 | OLP1 KO F1 | CTCCCCAGACAAGCACATTCC |
| TL405 | OLP1 KO R1 | CTGGCCGTCGTTTTACGACGCGTCTACACCACTCAGCAA |
| TL1253 | OLP1 KO F2 | GTCATAGCTGTTTCCTGCAAGATTCTGTGCGTATGGTGTGC |
| TL1254 | OLP1 KO R2 | GTTTGTTCTTTTGGCGGGTTTGAG |
| TL1255 | OLP1 KO F3 | ATATGAATTGCTGCGTGTGACC |
| TL1256 | OLP1 KO R3 | GCTTATGCTCCTTCTTCCAGTATT |
| TL410 | OLP1 KO F4 | TCCAAAGAAGAAGACAGCAACC |
| TL515 | OLP1 mCherry F | TTAGTAAACTCGCCCAACATGTCTGGATCCATGCCTATTCACACTCTTGCTTC |
| TL516 | OLP1 mCherry R | CTTGCTCACCATTCTAGAACTAGTGGATCCTTTGCCCTCTGGCTTGGTTCTG |
| TL539 | OLP1 QPCR F1 | TACGAGCCTCTCGACGATAC |
| TL540 | OLP1 QPCR R1 | TCTGGCTTGGTTCTGTCTTTAC |
| TL556 | OLP1 Comp F | GATATCGAATTCCTGCAGCCCGGGGGATCCTCCAAAGAAGAAGACAGCAACCTA |
| TL1257 | OLP1 Comp R | CGGTGGCGGCCGCTCTAGAACTAGTGGATCTTTGCCTACAGGATTTTGGTCACT |
| TL727 | OLP1 Pro-OLP1 F1 | TCGACGGTATCGATAAGCTTCGCCGACGACCAAGATACAG |
| TL729 | OLP1 Pro-OLP1 R1 | ATTCTAGAACTAGTGGATCCTTTGCCCTCTGGCTTGGTTCTGTC |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yu, Q.-K.; Han, L.-T.; Wu, Y.-J.; Liu, T.-B. The Role of Oxidoreductase-Like Protein Olp1 in Sexual Reproduction and Virulence of Cryptococcus neoformans. Microorganisms 2020, 8, 1730. https://doi.org/10.3390/microorganisms8111730
Yu Q-K, Han L-T, Wu Y-J, Liu T-B. The Role of Oxidoreductase-Like Protein Olp1 in Sexual Reproduction and Virulence of Cryptococcus neoformans. Microorganisms. 2020; 8(11):1730. https://doi.org/10.3390/microorganisms8111730
Chicago/Turabian StyleYu, Qi-Kun, Lian-Tao Han, Yu-Juan Wu, and Tong-Bao Liu. 2020. "The Role of Oxidoreductase-Like Protein Olp1 in Sexual Reproduction and Virulence of Cryptococcus neoformans" Microorganisms 8, no. 11: 1730. https://doi.org/10.3390/microorganisms8111730
APA StyleYu, Q.-K., Han, L.-T., Wu, Y.-J., & Liu, T.-B. (2020). The Role of Oxidoreductase-Like Protein Olp1 in Sexual Reproduction and Virulence of Cryptococcus neoformans. Microorganisms, 8(11), 1730. https://doi.org/10.3390/microorganisms8111730