Optimization of a Method for the Concentration of Genetic Material in Bacterial and Fungal Communities on Fresh Apple Peel Surfaces


Abstract
1. Introduction
2. Materials and Methods
2.1. Bath Sonication
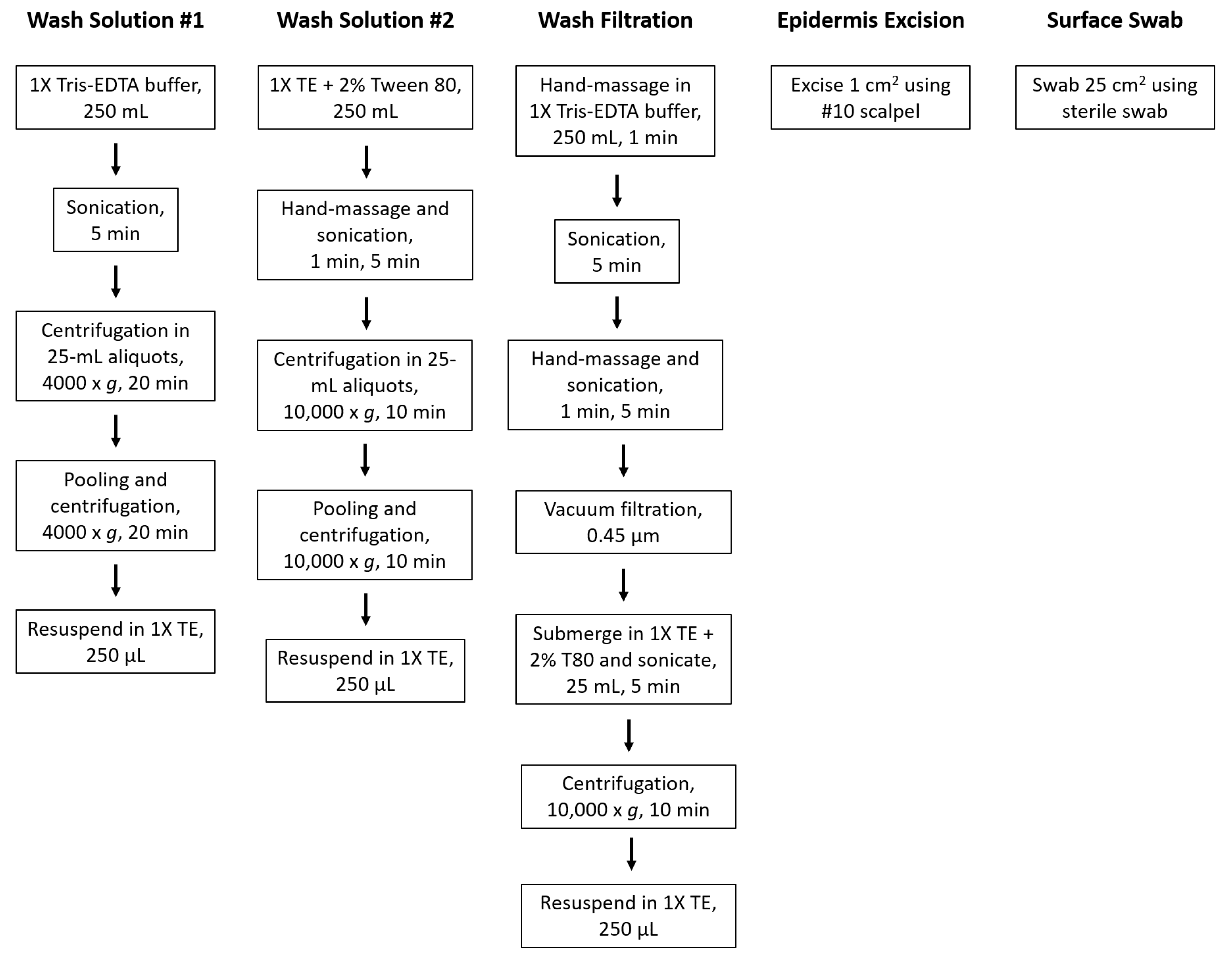
2.1.1. Wash Solution Variation and Method 1
2.1.2. Wash Solution Variation and Method 2
2.2. Wash Filtration
2.3. Epidermis Excision
2.4. Surface Swabbing
2.5. Microbial DNA Extraction
2.6. Quantification of Genetic Material
2.7. Quantitative Real-Time Polymerase Chain Reaction (qPCR) of Host, Bacterial, and Fungal Genes
2.7.1. Preparation of Microbiome Samples
2.7.2. Preparation of Bacterial, Fungal, and Plant Standards
2.7.3. qPCR Design
2.8. Statistical Analysis
3. Results
4. Discussion
5. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- USDA Economic Research Service. U.S. Per Capita Loss-Adjusted Fruit Availability, 2017. Available online: https://www.ers.usda.gov/data-products/chart-gallery/gallery/chart-detail/?chartId=58322 (accessed on 22 August 2020).
- U.S. Food and Drug Administration. Final Qualitative Assessment of Risk to Public Health from on Farm Contamination of Produce; U.S. Food and Drug Administration: Beltsville, MD, USA, 2016.
- Angelo, K.M.; Conrad, A.R.; Saupe, A.; Dragoo, H.; West, N.; Sorenson, A.; Barnes, A.; Doyle, M.; Beal, J.; Jackson, K.A.; et al. Multistate outbreak of Listeria monocytogenes infections linked to whole apples used in commercially produced, prepackaged caramel apples: United States, 2014–2015. Epidemiol. Infect. 2017, 145, 848–856. [Google Scholar] [CrossRef]
- U.S. Food and Drug Administration. North Bay Produce Voluntarily Recalls Fresh Apples Because of Possible Health Risk, 2019. Available online: https://www.fda.gov/safety/recalls-market-withdrawals-safety-alerts/north-bay-produce-voluntarily-recalls-fresh-apples-because-possible-health-risk (accessed on 22 August 2020).
- U.S. Food and Drug Administration. Fresh Pak Inc. Recalls Lot Specific Sliced Apple Products Because of Possible Health Risk, 2017. Available online: https://www.fda.gov/safety/recalls-market-withdrawals-safety-alerts/fresh-pak-inc-recalls-lot-specific-sliced-apple-products-because-possible-health-risk (accessed on 22 August 2020).
- U.S. Food and Drug Administration. Jack Brown Produce, Inc. Recalls Gala, Fuji, Honeycrisp and Golden Delicious Apples Due to Possible Health Risk, 2017. Available online: https://www.fda.gov/safety/recalls-market-withdrawals-safety-alerts/jack-brown-produce-inc-recalls-gala-fuji-honeycrisp-and-golden-delicious-apples-due-possible-health (accessed on 22 August 2020).
- U.S. Food and Drug Administration. Meijer Voluntarily Recalls Select Meijer Brand Fresh Packaged Products Containing Apples Due to Potential Health Risk, 2017. Available online: https://www.fda.gov/safety/recalls-market-withdrawals-safety-alerts/meijer-voluntarily-recalls-select-meijer-brand-fresh-packaged-products-containing-apples-due (accessed on 22 August 2020).
- Jagadeesan, B.; Gerner-Smidt, P.; Allard, M.W.; Leuillet, S.; Winkler, A.; Xiao, Y.; Chaffron, S.; Van Der Vossen, J.; Tang, S.; Katase, M.; et al. The use of next generation sequencing for improving food safety: Translation into practice. Food Microbiol. 2019, 79, 96–115. [Google Scholar] [CrossRef] [PubMed]
- Einson, J.E.; Rani, A.; You, X.; Rodriguez, A.A.; Randell, C.L.; Barnaba, T.; Mammel, M.K.; Kotewicz, M.L.; Elkins, C.A.; Sela, D.A. A Vegetable Fermentation Facility Hosts Distinct Microbiomes Reflecting the Production Environment. Appl. Environ. Microbiol. 2018, 84, 01680-18. [Google Scholar] [CrossRef] [PubMed]
- Jung, J.Y.; Lee, S.H.; Kim, J.M.; Park, M.S.; Bae, J.-W.; Hahn, Y.; Madsen, E.L.; Jeon, C.O. Metagenomic Analysis of Kimchi, a Traditional Korean Fermented Food. Appl. Environ. Microbiol. 2011, 77, 2264–2274. [Google Scholar] [CrossRef]
- Lusk, T.S.; Ottesen, A.; White, J.R.; Allard, M.W.; Brown, E.W.; Kase, J. Characterization of microflora in Latin-style cheeses by next-generation sequencing technology. BMC Microbiol. 2012, 12, 254. [Google Scholar] [CrossRef]
- Bokulich, N.A.; Mills, D.A. Facility-Specific “House” Microbiome Drives Microbial Landscapes of Artisan Cheesemaking Plants. Appl. Environ. Microbiol. 2013, 79, 5214–5223. [Google Scholar] [CrossRef]
- Allard, S.M.; Walsh, C.S.; Wallis, A.E.; Ottesen, A.; Brown, E.W.; Micallef, S.A. Solanum lycopersicum (tomato) hosts robust phyllosphere and rhizosphere bacterial communities when grown in soil amended with various organic and synthetic fertilizers. Sci. Total Environ. 2016, 573, 555–563. [Google Scholar] [CrossRef] [PubMed]
- Glenn, D.; Bassett, C.; Dowd, S. Effect of pest management system on ‘Empire’ apple leaf phyllosphere populations. Sci. Hortic. 2015, 183, 58–65. [Google Scholar] [CrossRef]
- Leff, J.W.; Fierer, N. Bacterial Communities Associated with the Surfaces of Fresh Fruits and Vegetables. PLoS ONE 2013, 8, e59310. [Google Scholar] [CrossRef]
- Lopez-Velasco, G.; Carder, P.A.; Welbaum, G.E.; Ponder, M.A. Diversity of the spinach (Spinacia oleracea) spermosphere and phyllosphere bacterial communities. FEMS Microbiol. Lett. 2013, 346, 146–154. [Google Scholar] [CrossRef]
- Kim, D.; Hong, S.; Kim, Y.-T.; Ryu, S.; Kim, H.B.; Lee, J.-H. Metagenomic Approach to Identifying Foodborne Pathogens on Chinese Cabbage. J. Microbiol. Biotechnol. 2018, 28, 227–235. [Google Scholar] [CrossRef] [PubMed]
- Yang, X.; Noyes, N.R.; Doster, E.; Martin, J.N.; Linke, L.M.; Magnuson, R.J.; Yang, H.; Geornaras, I.; Woerner, D.R.; Jones, K.L.; et al. Use of Metagenomic Shotgun Sequencing Technology To Detect Foodborne Pathogens within the Microbiome of the Beef Production Chain. Appl. Environ. Microbiol. 2016, 82, 2433–2443. [Google Scholar] [CrossRef] [PubMed]
- Dzieciol, M.; Schornsteiner, E.; Muhterem-Uyar, M.; Stessl, B.; Wagner, M.; Schmitz-Esser, S. Bacterial diversity of floor drain biofilms and drain waters in a Listeria monocytogenes contaminated food processing environment. Int. J. Food Microbiol. 2016, 223, 33–40. [Google Scholar] [CrossRef]
- Jarvis, K.G.; White, J.R.; Grim, C.J.; Ewing, L.; Ottesen, A.; Beaubrun, J.J.-G.; Pettengill, J.B.; Brown, E.W.; Hanes, D. Cilantro microbiome before and after nonselective pre-enrichment for Salmonella using 16S rRNA and metagenomic sequencing. BMC Microbiol. 2015, 15, 1–3. [Google Scholar] [CrossRef] [PubMed]
- Ottesen, A.; Gonzalez, A.; White, J.R.; Pettengill, J.B.; Li, C.; Allard, S.; Rideout, S.L.; Allard, M.W.; Hill, T.; Evans, P.; et al. Baseline survey of the anatomical microbial ecology of an important food plant: Solanum lycopersicum (tomato). BMC Microbiol. 2013, 13, 1–12. [Google Scholar] [CrossRef]
- Leonard, S.R.; Mammel, M.K.; Lacher, D.W.; Elkins, C.A. Application of Metagenomic Sequencing to Food Safety: Detection of Shiga Toxin-Producing Escherichia coli on Fresh Bagged Spinach. Appl. Environ. Microbiol. 2015, 81, 8183–8191. [Google Scholar] [CrossRef]
- Doyle, C.J.; O’Toole, P.W.; Cotter, P.D. Metagenome-based surveillance and diagnostic approaches to studying the microbial ecology of food production and processing environments. Environ. Microbiol. 2017, 19, 4382–4391. [Google Scholar] [CrossRef]
- Tan, X.; Chung, T.; Chen, Y.; Macarisin, D.; Laborde, L.; Kovac, J. The occurrence of Listeria monocytogenes is associated with built environment microbiota in three tree fruit processing facilities. Microbiome 2019, 7, 115–118. [Google Scholar] [CrossRef]
- Bokulich, N.A.; Lewis, Z.T.; Boundy-Mills, K.; Mills, D.A. A new perspective on microbial landscapes within food production. Curr. Opin. Biotechnol. 2016, 37, 182–189. [Google Scholar] [CrossRef]
- Berg, G.; Krechel, A.; Ditz, M.; Sikora, R.A.; Ulrich, A.; Hallmann, J. Endophytic and ectophytic potato-associated bacterial communities differ in structure and antagonistic function against plant pathogenic fungi. FEMS Microbiol. Ecol. 2005, 51, 215–229. [Google Scholar] [CrossRef]
- Wassermann, B.; Müller, H.; Berg, G. An Apple a Day: Which Bacteria Do We Eat With Organic and Conventional Apples? Front. Microbiol. 2019, 10, 1629. [Google Scholar] [CrossRef] [PubMed]
- Jacobs, J.L.; Sundin, G.W. Effect of Solar UV-B Radiation on a Phyllosphere Bacterial Community. Appl. Environ. Microbiol. 2001, 67, 5488–5496. [Google Scholar] [CrossRef] [PubMed]
- Smalla, K.; Goodall, T.I.; Houlden, A.; Gallego, V. Microbial diversity in the phyllosphere and rhizosphere of field grown crop plants: Microbial specialisation at the plant surface. Microb. Ecol. Aer. Plant Surf. 2009, 21, 21–36. [Google Scholar] [CrossRef]
- Fernández, V.; Sotiropoulos, T.; Brown, P.H. Foliar Fertilisation: Scientific Principles and Field Practices; International Fertilizer Industry Association: Paris, France, 2013. [Google Scholar]
- Thoendel, M.; Jeraldo, P.; Greenwood-Quaintance, K.E.; Yao, J.Z.; Chia, N.; Hanssen, A.D.; Abdel, M.P.; Patel, R. Comparison of microbial DNA enrichment tools for metagenomic whole genome sequencing. J. Microbiol. Methods 2016, 127, 141–145. [Google Scholar] [CrossRef]
- Biesbroek, G.; Sanders, E.A.M.; Roeselers, G.; Wang, X.; Caspers, M.P.M.; Trzcinski, K.; Bogaert, D.; Keijser, B.J.F. Deep Sequencing Analyses of Low Density Microbial Communities: Working at the Boundary of Accurate Microbiota Detection. PLoS ONE 2012, 7, e32942. [Google Scholar] [CrossRef]
- Dunbar, J.; White, S.; Forney, L. Genetic Diversity through the Looking Glass: Effect of Enrichment Bias. Appl. Environ. Microbiol. 1997, 63, 1326–1331. [Google Scholar] [CrossRef]
- Muniesa, M.; Blanch, A.R.; Lucena, F.; Jofre, J. Bacteriophages May Bias Outcome of Bacterial Enrichment Cultures. Appl. Environ. Microbiol. 2005, 71, 4269–4275. [Google Scholar] [CrossRef]
- Pettengill, J.B.; McAvoy, E.; White, J.R.; Allard, M.W.; Brown, E.W.; Ottesen, A. Using metagenomic analyses to estimate the consequences of enrichment bias for pathogen detection. BMC Res. Notes 2012, 5, 378. [Google Scholar] [CrossRef]
- Metzker, M.L. Sequencing technologies—The next generation. Nat. Rev. Genet. 2009, 11, 31–46. [Google Scholar] [CrossRef]
- Pareek, C.S.; Smoczynski, R.; Tretyn, A. Sequencing technologies and genome sequencing. J. Appl. Genet. 2011, 52, 413–435. [Google Scholar] [CrossRef]
- Hayden, E.C. Genome sequencing: The third generation. Nature 2009, 457, 768–769. [Google Scholar] [CrossRef]
- Pollock, J.; Glendinning, L.; Wisedchanwet, T.; Watson, M. The Madness of Microbiome: Attempting To Find Consensus “Best Practice” for 16S Microbiome Studies. Appl. Environ. Microbiol. 2018, 84, e02627-17. [Google Scholar] [CrossRef] [PubMed]
- Greathouse, K.L.; Sinha, R.; Vogtmann, E. DNA extraction for human microbiome studies: The issue of standardization. Genome Biol. 2019, 20, 212–214. [Google Scholar] [CrossRef] [PubMed]
- Carrigg, C.; Rice, O.; Kavanagh, S.; Collins, G.; O’Flaherty, V. DNA extraction method affects microbial community profiles from soils and sediment. Appl. Microbiol. Biotechnol. 2007, 77, 955–964. [Google Scholar] [CrossRef]
- Gerasimidis, K.; Bertz, M.; Quince, C.; Brunner, K.; Bruce, A.; Combet, E.; Calus, S.; Loman, N.J.; Ijaz, U.Z. The effect of DNA extraction methodology on gut microbiota research applications. BMC Res. Notes 2016, 9, 365. [Google Scholar] [CrossRef]
- Bürgmann, H.; Pesaro, M.; Widmer, F.; Zeyer, J. A strategy for optimizing quality and quantity of DNA extracted from soil. J. Microbiol. Methods 2001, 45, 7–20. [Google Scholar] [CrossRef]
- Yuan, S.; Cohen, D.B.; Ravel, J.; Abdo, Z.; Forney, L.J. Evaluation of Methods for the Extraction and Purification of DNA from the Human Microbiome. PLoS ONE 2012, 7, e33865. [Google Scholar] [CrossRef]
- Guo, F.; Zhang, T. Biases during DNA extraction of activated sludge samples revealed by high throughput sequencing. Appl. Microbiol. Biotechnol. 2012, 97, 4607–4616. [Google Scholar] [CrossRef]
- Henderson, G.; Cox, F.; Kittelmann, S.; Miri, V.H.; Zethof, M.; Noel, S.J.; Waghorn, G.C.; Janssen, P.H. Effect of DNA Extraction Methods and Sampling Techniques on the Apparent Structure of Cow and Sheep Rumen Microbial Communities. PLoS ONE 2013, 8, e74787. [Google Scholar] [CrossRef]
- Kennedy, N.A.; Walker, A.W.; Berry, S.H.; Duncan, S.H.; Farquarson, F.M.; Louis, P.; Thomson, J.M.; Satsangi, J.; Flint, H.J.; Parkhill, J.; et al. The Impact of Different DNA Extraction Kits and Laboratories upon the Assessment of Human Gut Microbiota Composition by 16S rRNA Gene Sequencing. PLoS ONE 2014, 9, e88982. [Google Scholar] [CrossRef]
- Hart, M.L.; Meyer, A.; Johnson, P.J.; Ericsson, A.C. Comparative Evaluation of DNA Extraction Methods from Feces of Multiple Host Species for Downstream Next-Generation Sequencing. PLoS ONE 2015, 10, e0143334. [Google Scholar] [CrossRef] [PubMed]
- Desneux, J.; Pourcher, A.-M. Comparison of DNA extraction kits and modification of DNA elution procedure for the quantitation of subdominant bacteria from piggery effluents with real-time PCR. Microbiologyopen 2014, 3, 437–445. [Google Scholar] [CrossRef] [PubMed]
- MacKenzie, B.W.; Waite, D.W.; Taylor, M.W. Evaluating variation in human gut microbiota profiles due to DNA extraction method and inter-subject differences. Front. Microbiol. 2015, 6, 130. [Google Scholar] [CrossRef]
- Verschuren, L.M.G.; Calus, M.P.L.; Jansman, A.J.M.; Bergsma, R.; Knol, E.F.; Gilbert, H.; Zemb, O.; Jansman, A.J.M. Fecal microbial composition associated with variation in feed efficiency in pigs depends on diet and sex. J. Anim. Sci. 2018, 96, 1405–1418. [Google Scholar] [CrossRef] [PubMed]
- Zemb, O.; Achard, C.S.; Hamelin, J.; De Almeida, M.; Gabinaud, B.; Cauquil, L.; Verschuren, L.M.; Godon, J. Absolute quantitation of microbes using 16S rRNA gene metabarcoding: A rapid normalization of relative abundances by quantitative PCR targeting a 16S rRNA gene spike-in standard. Microbiologyopen 2020, 9. [Google Scholar] [CrossRef]
- De Beeck, M.O.; Lievens, B.; Busschaert, P.; Declerck, S.; Vangronsveld, J.; Colpaert, J.V. Comparison and Validation of Some ITS Primer Pairs Useful for Fungal Metabarcoding Studies. PLoS ONE 2014, 9, e97629. [Google Scholar] [CrossRef] [PubMed]
- Cheng, T.; Xu, C.; Lei, L.; Li, C.; Zhang, Y.; Zhou, S. Barcoding the kingdom Plantae: New PCR primers forITSregions of plants with improved universality and specificity. Mol. Ecol. Resour. 2015, 16, 138–149. [Google Scholar] [CrossRef]
- Sare, A.R.; Stouvenakers, G.; Eck, M.; Lampens, A.; Goormachtig, S.; Jijakli, M.H.; Massart, S. Standardization of Plant Microbiome Studies: Which Proportion of the Microbiota is Really Harvested? Microorganisms 2020, 8, 342. [Google Scholar] [CrossRef]
- Stagnati, L.; Soffritti, G.; Lanubile, A.; Busconi, M. Comparison of six methods for the recovery of PCR-compatible microbial DNA from an agricultural biogas plant. Appl. Microbiol. Biotechnol. 2017, 101, 3907–3917. [Google Scholar] [CrossRef]
- Leverentz, B.; Conway, W.S.; Janisiewicz, W.; Abadias, M.; Kurtzman, C.P.; Camp, M.J. Biocontrol of the Food-Borne Pathogens Listeria monocytogenes and Salmonella enterica Serovar Poona on Fresh-Cut Apples with Naturally Occurring Bacterial and Yeast Antagonists. Appl. Environ. Microbiol. 2006, 72, 1135–1140. [Google Scholar] [CrossRef]
- Shen, Y.; Nie, J.-Y.; Dong, Y.; Kuang, L.; Li, Y.; Zhang, J. Compositional shifts in the surface fungal communities of apple fruits during cold storage. Postharvest Biol. Technol. 2018, 144, 55–62. [Google Scholar] [CrossRef]
- Abdelfattah, A.; Whitehead, S.R.; Macarisin, D.; Liu, J.; Burchard, E.; Freilich, S.; Dardick, C.; Droby, S.; Wisniewski, M. Effect of Washing, Waxing and Low-Temperature Storage on the Postharvest Microbiome of Apple. Microorganisms 2020, 8, 944. [Google Scholar] [CrossRef] [PubMed]
- Chiodini, R.J.; Dowd, S.E.; Chamberlin, W.M.; Galandiuk, S.; Davis, B.; Glassing, A. Microbial Population Differentials between Mucosal and Submucosal Intestinal Tissues in Advanced Crohn’s Disease of the Ileum. PLoS ONE 2015, 10, e0134382. [Google Scholar] [CrossRef] [PubMed]
- Glassing, A.; Dowd, S.; Galandiuk, S.; Davis, B.; Chiodini, R.J. Inherent bacterial DNA contamination of extraction and sequencing reagents may affect interpretation of microbiota in low bacterial biomass samples. Gut Pathog. 2016, 8, 24. [Google Scholar] [CrossRef]
- Varma, A.; Padh, H.; Shrivastava, N. Plant genomic DNA isolation: An art or a science. Biotechnol. J. 2007, 2, 386–392. [Google Scholar] [CrossRef]
- Murray, M.; Thompson, W.F. Rapid isolation of high molecular weight plant DNA. Nucleic Acids Res. 1980, 8, 4321–4326. [Google Scholar] [CrossRef]
- Maria, P.A.; Hirsikorpi, M.; Kamarainen, T.; Jaakola, J.; Hohtola, A. DNA isolation methods for medicinal and aromatic plants. Plant Mol. Biol Rep. 2001, 19, 273. [Google Scholar]
- Ruggieri, J.; Kemp, R.; Forman, S.; Van Eden, M.E. Techniques for Nucleic Acid Purification from Plant, Animal, and Microbial Samples. In Methods in Molecular Biology; Springer Science and Business Media LLC: Berlin, Germany, 2016; pp. 41–52. [Google Scholar]
- Zaccone, C.; Beneduce, L.; Lotti, C.; Martino, G.; Plaza, C. DNA occurrence in organic matter fractions isolated from amended, agricultural soils. Appl. Soil Ecol. 2018, 130, 134–142. [Google Scholar] [CrossRef]
- Sharma, R.; John, S.J.; Damgaard, D.M.; McAllister, T.A. Extraction of PCR-Quality Plant and Microbial DNA from Total Rumen Contents. Biotechniques 2003, 34, 92–97. [Google Scholar] [CrossRef]
- Pietrysiak, E.; Kummer, J.M.; Hanrahan, I.; Ganjyal, G.M. Efficacy of Surfactant Combined with Peracetic Acid in Removing Listeria innocua from Fresh Apples. J. Food Prot. 2019, 82, 1965–1972. [Google Scholar] [CrossRef]
- Kroupitski, Y.; Pinto, R.; Belausov, E.; Sela, S. Distribution of Salmonella typhimurium in romaine lettuce leaves. Food Microbiol. 2011, 28, 990–997. [Google Scholar] [CrossRef] [PubMed]
- Huang, K.; Tian, Y.; Salvi, D.; Karwe, M.; Nitin, N. Influence of Exposure Time, Shear Stress, and Surfactants on Detachment of Escherichia coli O157:H7 from Fresh Lettuce Leaf Surfaces During Washing Process. Food Bioprocess Technol. 2017, 11, 621–633. [Google Scholar] [CrossRef]
- Pietrysiak, E.; Kummer, J.; Hanrahan, I.; Ganjyal, G. The hurdle effect of hot air impingement drying and surfactant-sanitizer wash on removal of Listeria innocua from fresh apples. J. Food Prot. 2020. [Google Scholar] [CrossRef] [PubMed]
- Starke, R.; Jehmlich, N.; Alfaro, T.; Dohnalkova, A.; Capek, P.; Bell, S.L.; Hofmockel, K.S. Incomplete cell disruption of resistant microbes. Sci. Rep. 2019, 9, 1–5. [Google Scholar] [CrossRef]
- Tiam, S.K.; Comte, K.; Dalle, C.; Duval, C.; Pancrace, C.; Gugger, M.; Marie, B.; Yéprémian, C.; Bernard, C. Development of a new extraction method based on high-intensity ultra-sonication to study RNA regulation of the filamentous cyanobacteria Planktothrix. PLoS ONE 2019, 14, e0222029. [Google Scholar] [CrossRef]
- Zips, A.; Schaule, G.; Flemming, H.C. Ultrasound as a means of detaching biofilms. Biofouling 1990, 2, 323–333. [Google Scholar] [CrossRef]
- Pchelintsev, N.A.; Adams, P.D.; Nelson, D.M. Critical Parameters for Efficient Sonication and Improved Chromatin Immunoprecipitation of High Molecular Weight Proteins. PLoS ONE 2016, 11, e0148023. [Google Scholar] [CrossRef]
- Yagi, N.; Satonaka, K.; Horio, M.; Shimogaki, H.; Tokuda, Y.; Maeda, S. The Role of DNase and EDTA on DNA Degradation in Formaldehyde Fixed Tissues. Biotech. Histochem. 1996, 71, 123–129. [Google Scholar] [CrossRef]
- Saeki, K.; Kunito, T.; Sakai, M. Effect of Tris-HCl Buffer on DNA Adsorption by a Variety of Soil Constituents. Microbes Environ. 2011, 26, 88–91. [Google Scholar] [CrossRef]
- Frontino, G.C. Comparison of Methods for Detection of Listeria on Wooden Shelves Used for Cheese Aging: Challenges Associated With Sampling Porous Surfaces. Master’s Thesis, University of Vermont, Burlington, VT, USA, 2019. [Google Scholar]
- Brauge, T.; Barre, L.; Leleu, G.; André, S.; Denis, C.; Hanin, A.; Frémaux, B.; Guilbaud, M.; Herry, J.-M.; Oulahal, N.; et al. European survey and evaluation of sampling methods recommended by the standard EN ISO 18593 for the detection of Listeria monocytogenes and Pseudomonas fluorescens on industrial surfaces. FEMS Microbiol. Lett. 2020, 367, 057. [Google Scholar] [CrossRef]
- Branck, T.A.; Hurley, M.J.; Prata, G.N.; Crivello, C.A.; Marek, P.J. Efficacy of a Sonicating Swab for Removal and Capture of Listeria monocytogenes in Biofilms on Stainless Steel. Appl. Environ. Microbiol. 2017, 83, e00109–e00117. [Google Scholar] [CrossRef] [PubMed]


{kind=link}
| Primer | Primer Sequence | Primer Target (Product Size; bp) | Cycle Information | Source |
|---|---|---|---|---|
| 343F | TACGGRAGGCAGCAG | Bacterial 16S (510) | (95 °C, 30 s; 60 °C, 60 s) × 40 | [51,52] |
| 784R | TACCAGGGTATCTAATCCT | |||
| ITS86F | GTGAATCATCGAATCTTTGAA | Fungal ITS2 (369) | (95 °C, 30 s; 55 °C, 30 s; 72 °C, 60 s) × 40 | [53] |
| ITS4R | TCCTCCGCTTATTGATATGC | |||
| ITS-p5F | CCTTATCAYTTAGAGGAAGGAG | Plant ITS1 (408) | (94 °C, 30 s; 55 °C, 40 s; 72 °C, 60 s) × 34 | [54] |
| ITS-u2R | GCGTTCAAAGAYTCGATGRTTC |
| Sample Collection Method | Mean DNA ± Std. Error (pg/μL) | Samples Below Limit of Detection (%) |
|---|---|---|
| Wash solution #1 | 24.93 ± 13.6 AB 1 | 6 (66.7) |
| Wash solution #2 | 614.82 ± 366.8 AB | 1 (11.1) |
| Wash filtration | 5.33 ± 4.9 A | 8 (88.9) |
| Epidermis excision | 661.67 ± 153.2 B | 0 (0) |
| Surface swabbing | ≤0.50 A | 9 (100) |
| Sample Collection Method | Mean Microbial Populations ± Std. Dev. (Log10 CFU/Sample) | |
|---|---|---|
| Bacterial | Fungal | |
| Wash solution #1 | 4.22 ± 0.15 A 1 | 5.39 ± 0.20 A |
| Wash solution #2 | 5.24 ± 0.54 B | 5.11 ± 0.39 A |
| Wash filtration | 4.86 ± 0.35 B | 5.18 ± 0.26 A |
| Epidermis excision | 2.25 ± 0.42 C | 2.00 ± 0.00 B |
| Surface swabbing | 1.22 ± 0.28 D | 2.67 ± 0.22 C |
| Sample Collection Method | Estimated Log10 Copy Number of Gene Target ± Std. Dev. | ||
|---|---|---|---|
| Bacterial | Fungal | Plant | |
| Wash solution #1 | 1.90 ± 0.00 A 1 | 5.30 ± 0.82 A | ND 2 A |
| Wash solution #2 | 3.04 ± 0.77 A | 7.79 ± 0.20 B | ND A |
| Wash filtration | 2.15 ± 0.83 A | 8.61 ± 0.30 C | ND A |
| Epidermis excision | 2.77 ± 0.82 A | 5.29 ± 0.25 A | 6.25 ± 1.23 B |
| Surface swabbing | 1.94 ± 0.66 A | 6.71 ± 0.69 D | ND A |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hamilton, A.; Harper, S.J.; Critzer, F. Optimization of a Method for the Concentration of Genetic Material in Bacterial and Fungal Communities on Fresh Apple Peel Surfaces. Microorganisms 2020, 8, 1480. https://doi.org/10.3390/microorganisms8101480
Hamilton A, Harper SJ, Critzer F. Optimization of a Method for the Concentration of Genetic Material in Bacterial and Fungal Communities on Fresh Apple Peel Surfaces. Microorganisms. 2020; 8(10):1480. https://doi.org/10.3390/microorganisms8101480
Chicago/Turabian StyleHamilton, Alexis, Scott J. Harper, and Faith Critzer. 2020. "Optimization of a Method for the Concentration of Genetic Material in Bacterial and Fungal Communities on Fresh Apple Peel Surfaces" Microorganisms 8, no. 10: 1480. https://doi.org/10.3390/microorganisms8101480
APA StyleHamilton, A., Harper, S. J., & Critzer, F. (2020). Optimization of a Method for the Concentration of Genetic Material in Bacterial and Fungal Communities on Fresh Apple Peel Surfaces. Microorganisms, 8(10), 1480. https://doi.org/10.3390/microorganisms8101480