Host Specificity and Differential Pathogenicity of Pectobacterium Strains from Dicot and Monocot Hosts

 , ,
, ,
Abstract
1. Introduction
2. Materials and Methods
2.1. Bacterial Strains, Plants and Growth Media
2.2. Virulence Assays
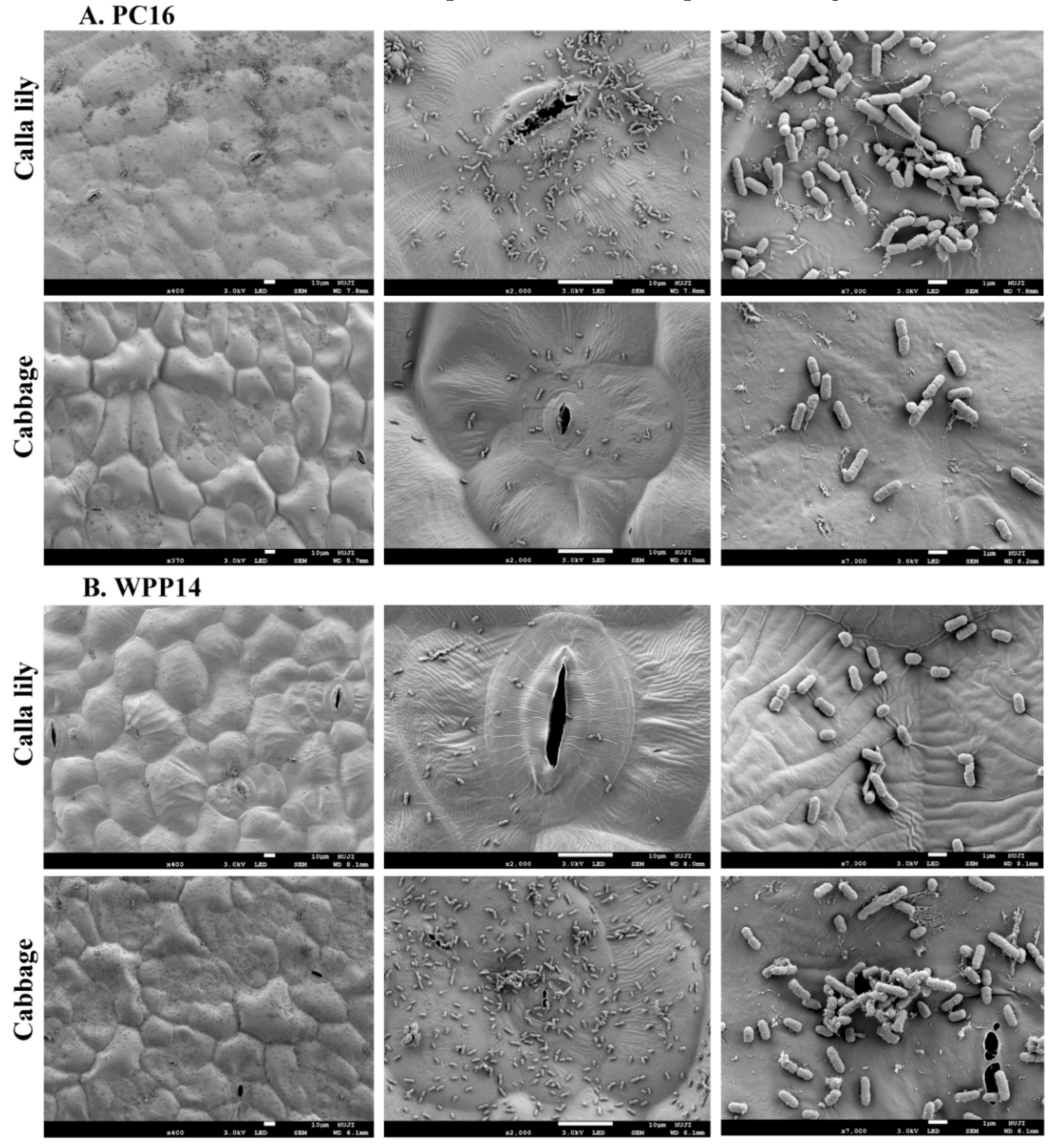
2.3. Scanning Electron. Microscopy (SEM)
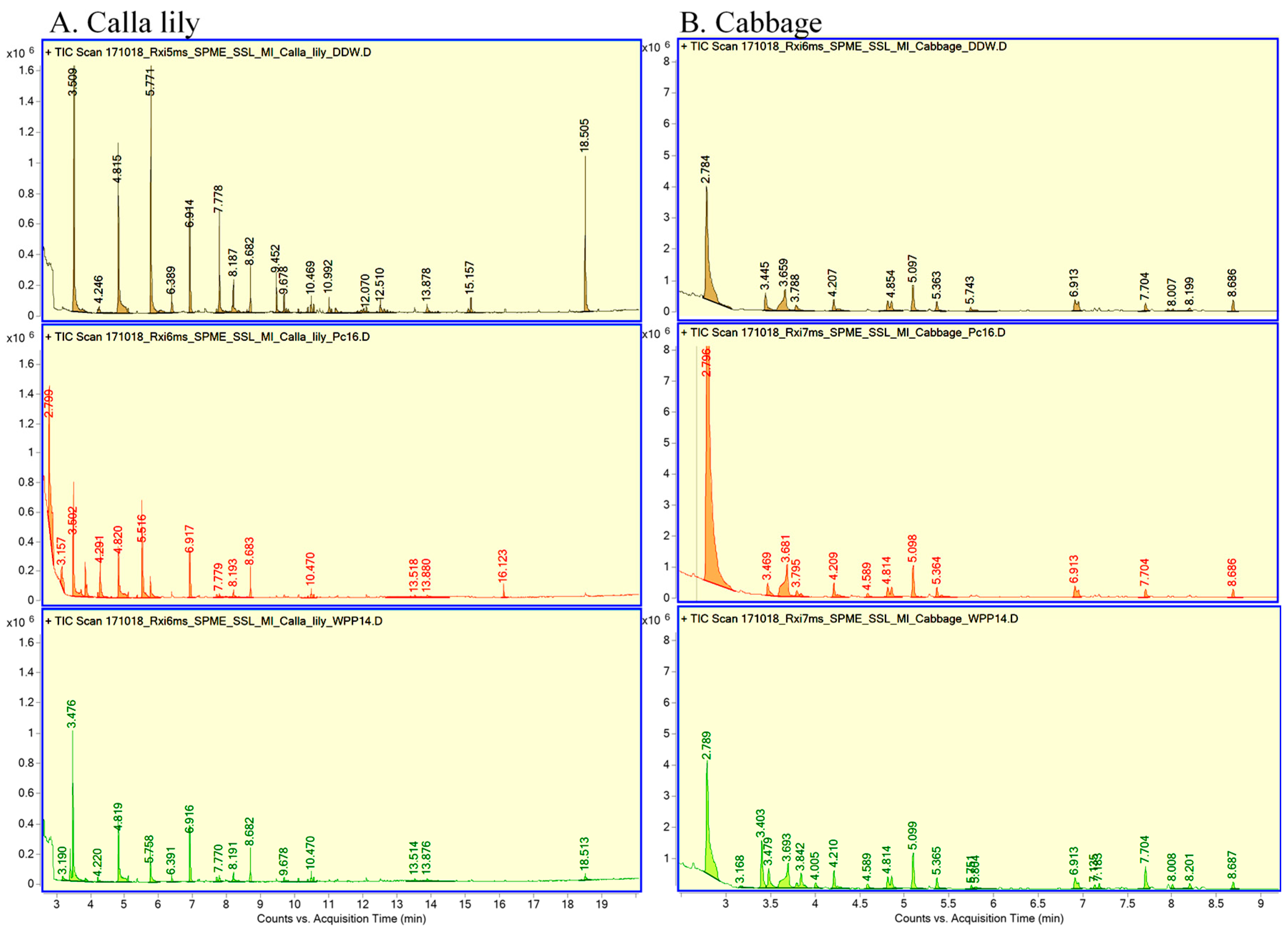
2.4. Analysis of Volatile Organic Compounds (VOCs)
2.5. Plant. Extracts
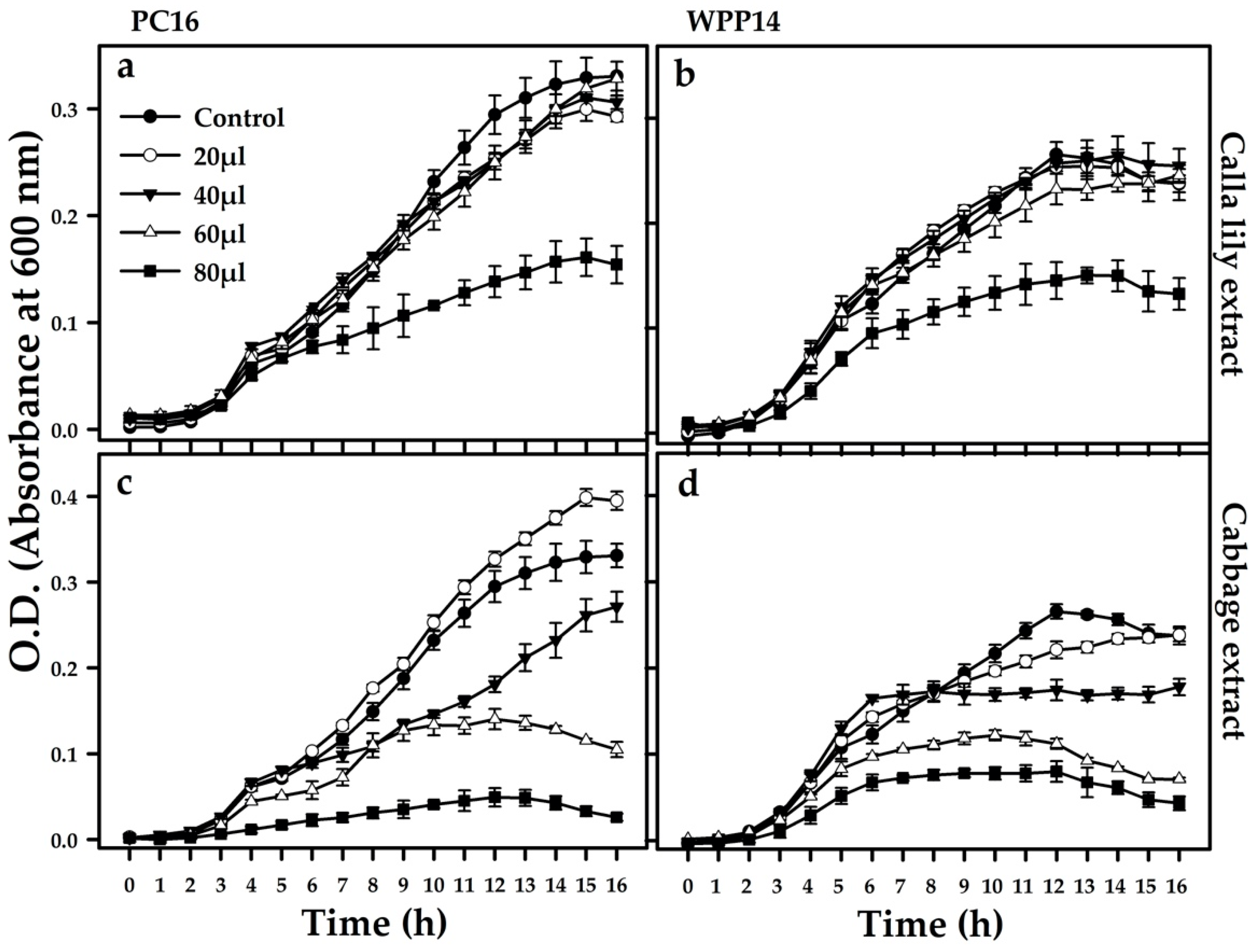
2.6. Growth Assessment
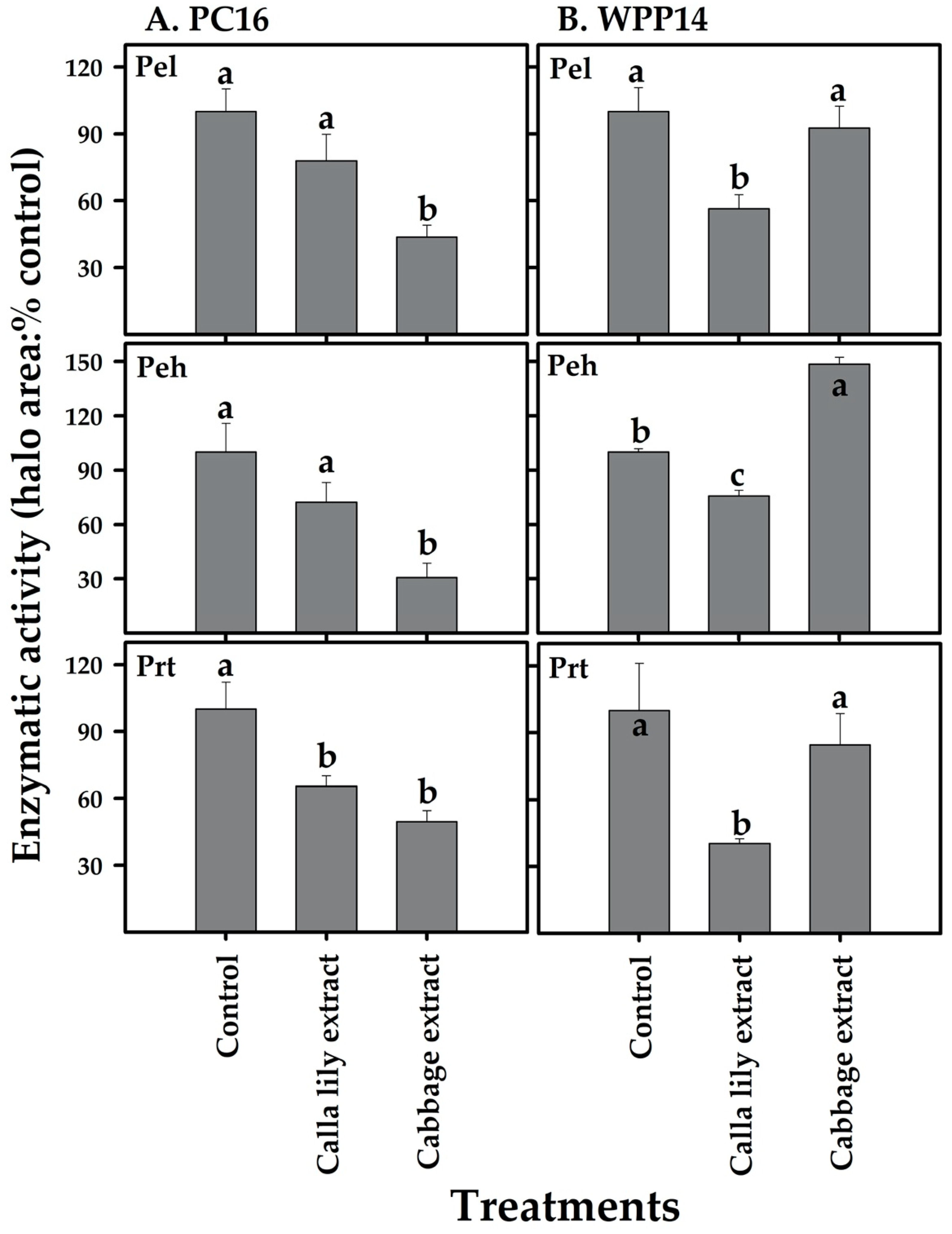
2.7. Exoenzyme Activity
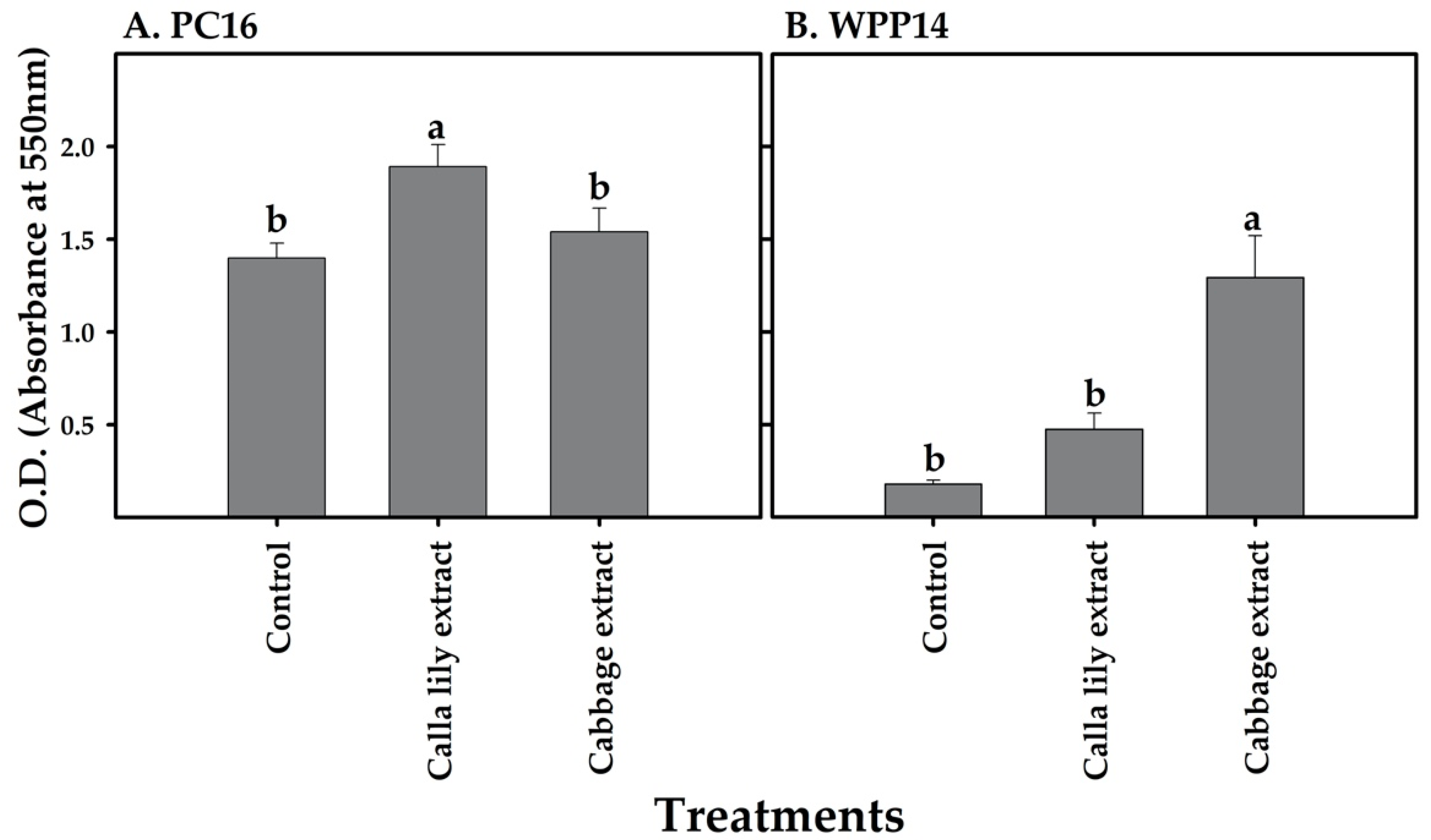
2.8. Biofilm Formation
2.9. Total RNA Extraction and cDNA Preparation
2.9.1. Extraction of RNA from the Pectobacteria
2.9.2. Extraction of RNA from Calla Lily and Cabbage
2.9.3. cDNA Preparation
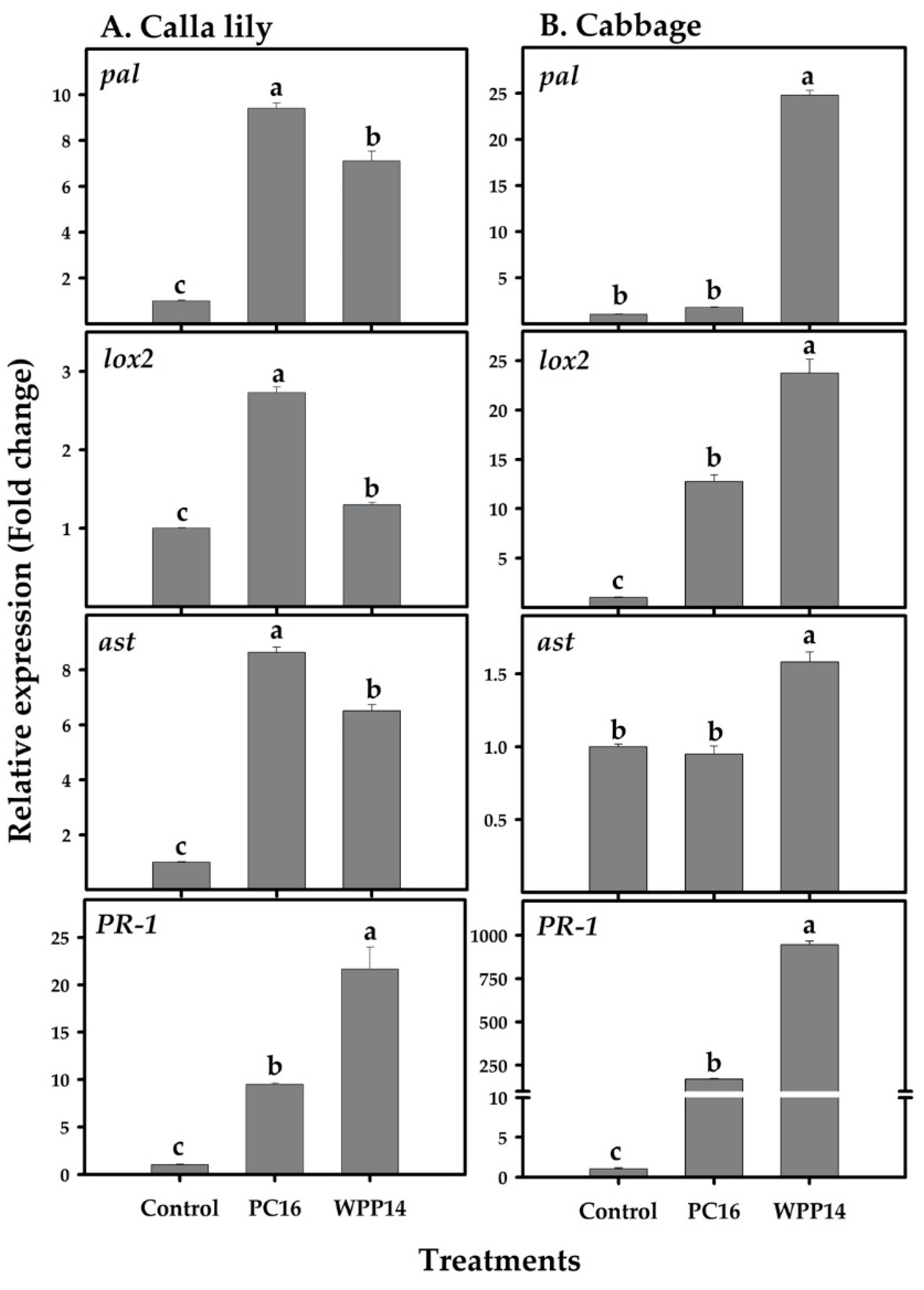
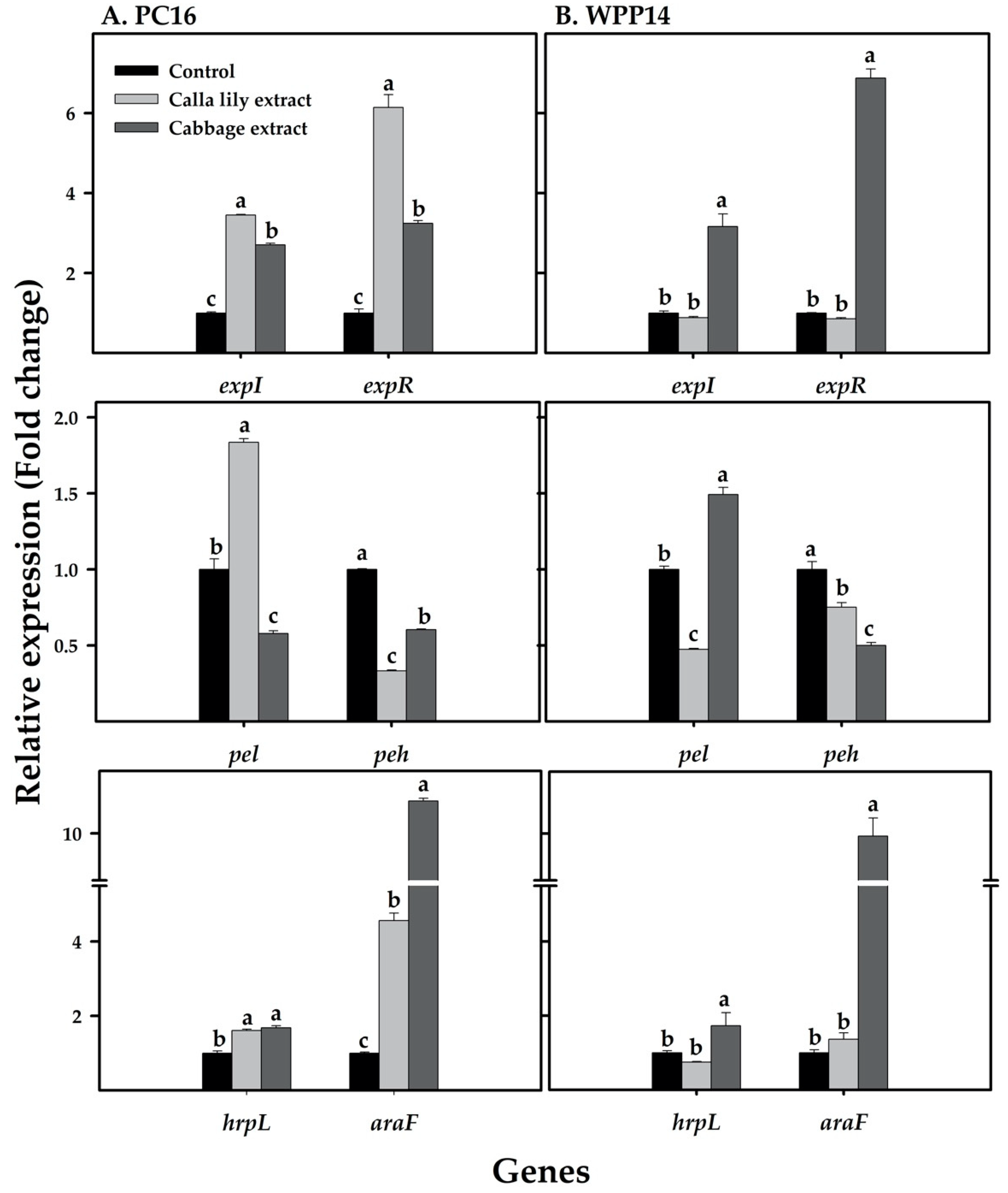
2.10. Primer Design and mRNA Quantification by Real-time qRT-PCR
2.11. Statistical Analysis
3. Results
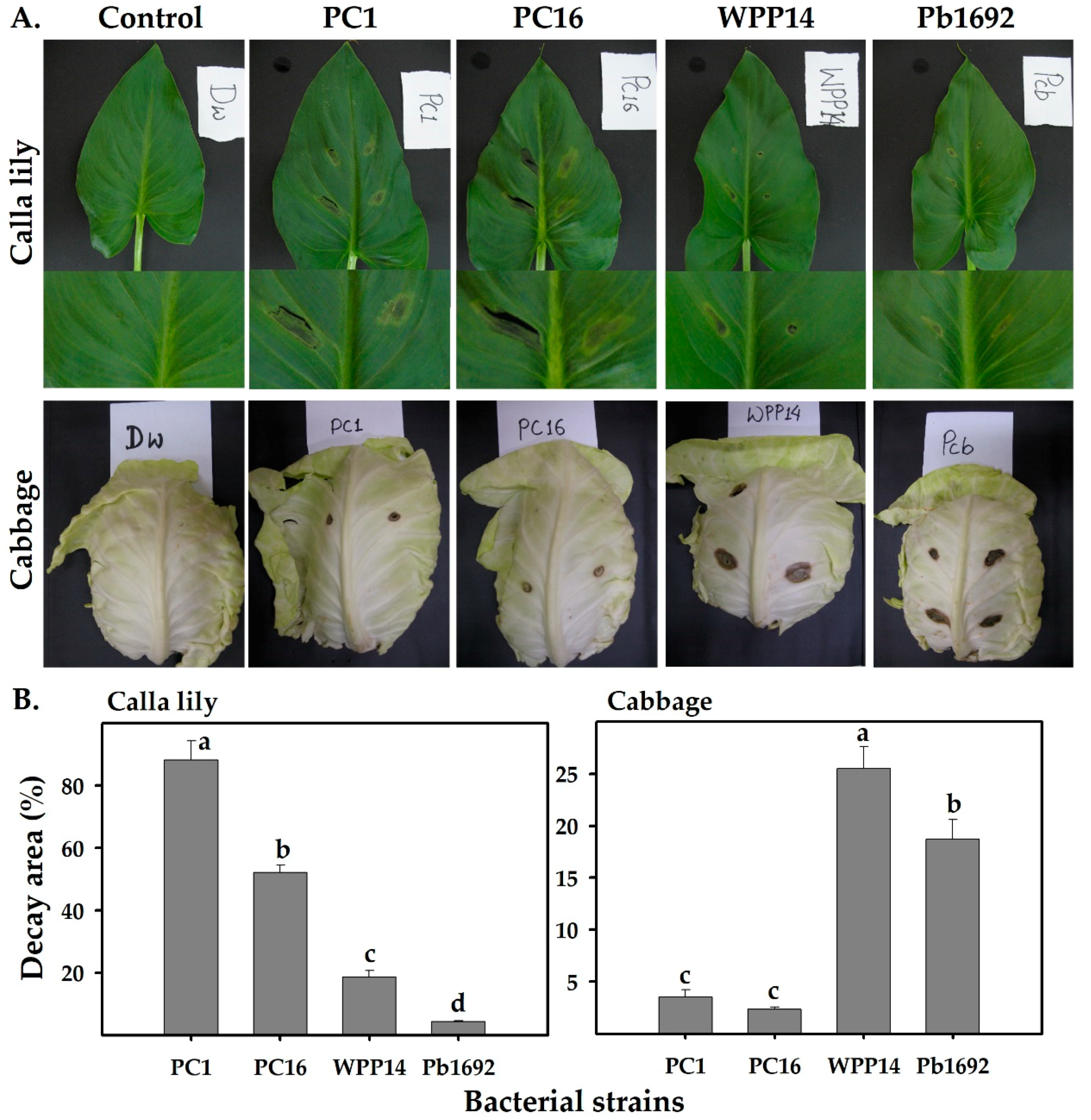
3.1. Virulence of Pectobacterium Strains on Different Hosts
3.2. Attachment of Pectobacterium Strains to Host Leaf Surfaces
3.3. Production of VOCs by Calla Lily and by Cabbage in Response to Inoculation of Pectobacterum
3.4. Expression of Defense-related Genes in Calla Lily and Cabbage in Response to Pectobacterium Infection
3.5. Effect of Host Phenolics on the Growth of the Pectobacterium Strains
3.6. Effect of Host Phenolic Extracts on Bacterial Exoenzyme Activity
3.7. Effect of Plant. Extracts on Biofilm Formation
3.8. Expression of Pectobacterium Virulence Genes in Response to Host Phenolic Extracts
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Ma, B.; Hibbing, M.E.; Kim, H.S.; Reedy, R.M.; Yedidia, I.; Breuer, J.; Breuer, J.; Glasner, J.D.; Perna, N.T.; Kelman, A.; et al. Host range and molecular phylogenies of the soft rot enterobacterial genera Pectobacterium and Dickeya. Phytopathol. 2007, 97, 1150–1163. [Google Scholar] [CrossRef]
- Pérombelon, M.C.M. Potato diseases caused by soft rot erwinias: An overview of pathogenesis. Plant Pathol. 2002, 51, 1–12. [Google Scholar] [CrossRef]
- Pitman, A.R.; Harrow, S.A.; Visnovsky, S.B. Genetic characterisation of Pectobacterium wasabiae causing soft rot disease of potato in New Zealand. Eur. J. Plant Pathol. 2010, 126, 423–435. [Google Scholar] [CrossRef]
- Charkowski, A.O. The changing face of bacterial soft-rot diseases. Annu. Rev. Phytopathol. 2018, 56, 269–288. [Google Scholar] [CrossRef] [PubMed]
- Van der Merwe, J.J.; Coutinho, T.A.; Korsten, L.; van der Waals, J.E. Pectobacterium carotovorum subsp. brasiliensis causing blackleg on potatoes in South Africa. Eur. J. Plant Pathol. 2010, 126, 175–185. [Google Scholar] [CrossRef]
- Adeolu, M.; Alnajar, S.; Naushad, S.; Gupta, R.S. Genome-based phylogeny and taxonomy of the ‘Enterobacteriales’: Proposal for Enterobacterales ord. nov. divided into the families Enterobacteriaceae, Erwiniaceae fam. nov., Pectobacteriaceae fam. nov., Yersiniaceae fam. nov., Hafniaceae fam. nov., Morganellaceae fam. nov., and Budviciaceae fam. nov. Int. J. Syst. Evol. Microbiol. 2016, 66, 5575–5599. [Google Scholar] [CrossRef] [PubMed]
- Li, L.; Yuan, L.; Shi, Y.; Xie, X.; Chai, A.; Wang, Q.; Li, B. Comparative genomic analysis of Pectobacterium carotovorum subsp. brasiliense SX309 provides novel insights into its genetic and phenotypic features. BMC Genomics 2019, 20, 486. [Google Scholar] [CrossRef]
- Pasanen, M.; Waleron, M.; Schott, T.; Cleenwerck, I.; Misztak, A.; Waleron, K.; Pritchard, L.; Bakr, R.; Degefu, Y.; van der Wolf, J.; et al. Pectobacterium parvum sp. nov., having a Salmonella SPI-1-like Type III secretion system and low virulence. Int. J. Syst. Evol. Microbiol. 2020, 70, 2440–2448. [Google Scholar] [CrossRef] [PubMed]
- Portier, P.; Pédron, J.; Taghouti, G.; Fischer-Le Saux, M.; Caullireau, E.; Bertrand, C.; Laurent, A.; Chawki, K.; Oulgazi, S.; Moumni, M.; et al. Elevation of Pectobacterium carotovorum subsp. odoriferum to species level as Pectobacterium odoriferum sp. nov., proposal of Pectobacterium brasiliense sp. nov. and Pectobacterium actinidiae sp. nov., emended description of Pectobacterium carotovorum and description of Pectobacterium versatile sp. nov., isolated from streams and symptoms on diverse plants. Int. J. Syst. Evol. Microbiol. 2019, 69, 3207–3216. [Google Scholar] [CrossRef] [PubMed]
- Waleron, M.; Misztak, A.; Waleron, M.; Jonca, J.; Furmaniak, M.; Waleron, K. Pectobacterium polonicum sp. nov. isolated from vegetable fields. Int. J. Syst. Evol. Microbiol. 2019, 69, 1751–1759. [Google Scholar] [CrossRef]
- Zhang, Y.C.; Fan, Q.R.; Loria, R. A re-evaluation of the taxonomy of phytopathogenic genera Dickeya and Pectobacterium using whole-genome sequencing data. Syst. Appl. Microbiol. 2016, 39, 252–259. [Google Scholar] [CrossRef] [PubMed]
- Nabhan, S.; De Boer, S.H.; Maiss, E.; Wydra, K. Pectobacterium aroidearum sp nov., a soft rot pathogen with preference for monocotyledonous plants. Int. J. Syst. Evol. Microbiol. 2013, 63, 2520–2525. [Google Scholar] [CrossRef] [PubMed]
- Yishay, M.; Burdman, S.; Valverde, A.; Luzzatto, T.; Ophir, R.; Yedidia, I. Differential pathogenicity and genetic diversity among Pectobacterium carotovorum ssp. carotovorum isolates from monocot and dicot hosts support early genomic divergence within this taxon. Environ. Microbiol. 2008, 10, 2746–2759. [Google Scholar] [CrossRef] [PubMed]
- Ahmed, F.A.; Larrea-Sarmiento, A.; Alvarez, A.M.; Arif, M. Genome-informed diagnostics for specific and rapid detection of Pectobacterium species using recombinase polymerase amplification coupled with a lateral flow device. Sci. Rep. 2018, 8, 15972. [Google Scholar] [CrossRef] [PubMed]
- Waleron, M.; Misztak, A.; Waleron, M.; Franczuk, M.; Jońca, J.; Wielgomas, B.; Mikiciński, A.; Popović, T.; Waleron, K. Pectobacterium zantedeschiae sp. nov. a new species of a soft rot pathogen isolated from Calla lily (Zantedeschia spp.). Syst. Appl. Microbiol. 2019, 42, 275–283. [Google Scholar] [CrossRef]
- Seo, S.T.; Furuya, N.; Lim, C.K.; Takanami, Y.; Tsuchiya, K. Phenotypic and genetic diversity of Erwinia carotovora ssp. carotovora strains from Asia. J. Phytopathol. 2002, 150, 120–127. [Google Scholar] [CrossRef]
- Smith, C.; Bartz, J.A. Variation in the pathogenicity and aggressiveness of strains of Erwinia carotovora subsp. carotovora isolated from different hosts. Plant Dis. 1990, 74, 505–509. [Google Scholar] [CrossRef]
- Wright, P.J. A soft rot of calla (Zantedeschia spp.) caused by Erwinia carotovora subspecies carotovora. N. Z. J. Crop. Hortic. Sci. 1998, 26, 331–334. [Google Scholar] [CrossRef]
- Norman, D.J.; Yuen, J.M.F.; Resendiz, R.; Boswell, J. Characterization of Erwinia populations from nursery retention ponds and lakes infecting ornamental plants in Florida. Plant Dis. 2003, 87, 193–196. [Google Scholar] [CrossRef]
- Ying, F.X.; Hu, X.F.; Chen, J.S. First report of soft rot caused by Pectobacterium carotovorum on Pinellia ternata in China. Plant Dis. 2007, 91, 1359. [Google Scholar] [CrossRef]
- van der Wolf, J.M.; de Haan, E.G.; Kastelein, P.; Krijger, M.; de Haas, B.H.; Velvis, H.; Mendes, O.; Kooman-Gersmann, M.; van der Zouwen, P.S. Virulence of Pectobacterium carotovorum subsp. brasiliense on potato compared with that of other Pectobacterium and Dickeya species under climatic conditions prevailing in the Netherlands. Plant Pathol. 2017, 66, 571–583. [Google Scholar] [CrossRef]
- Glasner, J.D.; Marquez-Villavicencio, M.; Kim, H.S.; Jahn, C.E.; Ma, B.; Biehl, B.S.; Rissman, A.I.; Mole, B.; Yi, X.; Yang, C.H.; et al. Niche-specificity and the variable fraction of the Pectobacterium pan-genome. Mol. Plant-Microbe Interact. 2008, 21, 1549–1560. [Google Scholar] [CrossRef] [PubMed]
- Yap, M.N.; Barak, J.D.; Charkowski, A.O. Genomic diversity of Erwinia carotovora subsp. carotovora and its correlation with virulence. Appl. Environ. Microbiol. 2004, 70, 3013–3023. [Google Scholar] [CrossRef]
- Kersey, C.M.; Agyemang, P.A.; Dumenyo, C.K. CorA, the magnesium/nickel/cobalt transporter, affects virulence and extracellular enzyme production in the soft rot pathogen Pectobacterium carotovorum. Mol. Plant Pathol. 2012, 13, 58–71. [Google Scholar] [CrossRef]
- Luzzatto, T.; Golan, A.; Yishay, M.; Bilkis, I.; Ben-Ari, J.; Yedidia, I. Priming of antimicrobial phenolics during induced resistance response towards Pectobacterium carotovorum in the ornamental monocot calla lily. J. Agric. Food Chem. 2007, 55, 10315–10322. [Google Scholar] [CrossRef]
- Chatterjee, A.; Cui, Y.; Liu, Y.; Dumenyo, C.K.; Chatterjee, A.K. Inactivation of rsmA leads to overproduction of extracellular pectinases, cellulases, and proteases in Erwinia carotovora subsp. carotovora in the absence of the starvation cell density-sensing signal, N-(3-oxohexanoyl)-L-homoserine lactone. Appl. Environ. Microbiol. 1995, 61, 1959–1967. [Google Scholar] [CrossRef]
- O’Toole, G.A. Microtiter dish biofilm formation assay. J. Vis. Exp. 2011, 30, e2437. [Google Scholar] [CrossRef]
- Joshi, J.R.; Burdman, S.; Lipsky, A.; Yariv, S.; Yedidia, I. Plant phenolic acids affect the virulence of Pectobacterium aroidearum and P. carotovorum ssp. brasiliense via quorum sensing regulation. Mol. Plant Pathol. 2016, 17, 487–500. [Google Scholar] [CrossRef]
- Ponzio, C.; Gols, R.; Pieterse, C.M.J.; Dicke, M. Ecological and phytohormonal aspects of plant volatile emission in response to single and dual infestations with herbivores and phytopathogens. Funct. Ecol. 2013, 27, 587–598. [Google Scholar] [CrossRef]
- Sharifi, R.; Lee, S.M.; Ryu, C.M. Microbe-induced plant volatiles. New Phytol. 2018, 220, 684–691. [Google Scholar] [CrossRef]
- Whitney, E.D.; Mackey, B.E. Differences in aggressiveness of Erwinia carotovora subsp. betavasculorum strains and their reactions to sugar beet cultivars. Plant Dis. 1989, 73, 220–222. [Google Scholar] [CrossRef]
- Helias, V.; Le Roux, A.C.; Bertheau, Y.; Andrivon, D.; Gauthier, J.P.; Jouan, B. Characterisation of Erwinia carotovora subspecies and detection of Erwinia carotovora subsp. atroseptica in potato plants, soil and water extracts with PCR-based methods. Eur. J. Plant Pathol. 1998, 104, 685–699. [Google Scholar] [CrossRef]
- Nabhan, S.; Wydra, K.; Linde, M.; Debener, T. The use of two complementary DNA assays, AFLP and MLSA, for epidemic and phylogenetic studies of pectolytic enterobacterial strains with focus on the heterogeneous species Pectobacterium carotovorum. Plant Pathol. 2012, 61, 498–508. [Google Scholar] [CrossRef]
- Nykyri, J.; Niemi, O.; Koskinen, P.; Nokso-Koivisto, J.; Pasanen, M.; Broberg, M.; Plyusnin, I.; Törönen, P.; Holm, L.; Pirhonen, M.; et al. Revised phylogeny and novel horizontally acquired virulence determinants of the model soft rot phytopathogen Pectobacterium wasabiae SCC3193. PLoS Pathog. 2012, 8, e1003013. [Google Scholar] [CrossRef]
- Barnard, A.M.L.; Bowden, S.D.; Burr, T.; Coulthurst, S.J.; Monson, R.E.; Salmond, G.P.C. Quorum sensing, virulence and secondary metabolite production in plant soft-rotting bacteria. Philos. Trans. R. Soc. Lond. B. Biol. Sci. 2007, 362, 1165–1183. [Google Scholar] [CrossRef]
- El Abed, S.; Ibnsouda, S.K.; Latrache, H.; Hamadi, F. Scanning electron microscopy (SEM) and environmental SEM: Suitable tools for study of adhesion stage and biofilm formation. In Scanning Electron Microscopy; Kazmiruk, V., Ed.; Intechopen: London, UK, 2012. [Google Scholar] [CrossRef]
- Qiu, R.; Qu, D.; Trengove, R.; Agarwal, M.; Hardy, G.E.S.; Ren, Y.L. Headspace solid-phase microextraction and gas chromatography-mass spectrometry for analysis of VOCs produced by Phytophthora cinnamomi. Plant Dis. 2014, 98, 1099–1105. [Google Scholar] [CrossRef]
- Taniguchi, S.; Hosokawa-Shinonaga, Y.; Tamaoki, D.; Yamada, S.; Akimitsu, K.; Gomi, K. Jasmonate induction of the monoterpene linalool confers resistance to rice bacterial blight and its biosynthesis is regulated by JAZ protein in rice. Plant Cell Environ. 2014, 37, 451–461. [Google Scholar] [CrossRef]
- Costello, B.P.J.D.; Evans, P.; Ewen, R.J.; Gunson, H.E.; Ratcliffe, N.M.; Spencer-Phillips, P.T.N. Identification of volatiles generated by potato tubers (Solanum tuberosum CV-Maris Piper) infected by Erwinia carotovora, Bacillus polymyxa and Arthrobacter spp. Plant Pathol. 1999, 48, 345–351. [Google Scholar] [CrossRef]
- Vikram, A.; Lui, L.H.; Hossain, A.; Kushalappa, A.C. Metabolic fingerprinting to discriminate diseases of stored carrots. Ann. Appl. Biol. 2006, 148, 17–26. [Google Scholar] [CrossRef]
- Glazebrook, J. Contrasting mechanisms of defense against biotrophic and necrotrophic pathogens. Annu. Rev. Phytopathol. 2005, 43, 205–227. [Google Scholar] [CrossRef]
- Davidsson, P.R.; Kariola, T.; Niemi, O.; Palva, E.T. Pathogenicity of and plant immunity to soft rot pectobacteria. Front. Plant Sci. 2013, 4, 191. [Google Scholar] [CrossRef]
- Liu, H.; Coulthurst, S.J.; Pritchard, L.; Hedley, P.E.; Ravensdale, M.; Humphris, S.; Burr, T.; Takle, G.; Brurberg, M.-B.; Birch, P.R.J.; et al. Quorum sensing coordinates brute force and stealth modes of infection in the plant pathogen Pectobacterium atrosepticum. PLoS Pathog. 2008, 4, e1000093. [Google Scholar] [CrossRef] [PubMed]
- Vidal, S.; de Leon, I.P.; Denecke, J.; Palva, E.T. Salicylic acid and the plant pathogen Erwinia carotovora induce defense genes via antagonistic pathways. Plant J. 1997, 11, 115–123. [Google Scholar] [CrossRef]
- Yan, L.; Zhai, Q.; Wei, J.; Li, S.; Wang, B.; Huang, T.; Du, M.; Sun, J.; Kang, L.; Li, C.-B.; et al. Role of tomato lipoxygenase D in wound-induced jasmonate biosynthesis and plant immunity to insect herbivores. PLoS Genet. 2013, 9, e1003964. [Google Scholar] [CrossRef]
- Gigot, C.; Ongena, M.; Fauconnier, M.-L.; Wathelet, J.-P.; Du Jardin, P.; Thonart, P. The lipoxygenase metabolic pathway in plants: Potential for industrial production of natural green leaf volatiles. Biotechnol. Agron. Soc. Environ. 2010, 14, 451–460. [Google Scholar]
- Mitsuhara, I.; Iwai, T.; Seo, S.; Yanagawa, Y.; Kawahigasi, H.; Hirose, S.; Ohkawa, Y.; Ohashi, Y. Characteristic expression of twelve rice PR1 family genes in response to pathogen infection, wounding, and defense-related signal compounds (121/180). Mol. Genet. Genomics 2008, 279, 415–427. [Google Scholar] [CrossRef]
- Cheynier, V.; Comte, G.; Davies, K.M.; Lattanzio, V.; Martens, S. Plant phenolics: Recent advances on their biosynthesis, genetics, and ecophysiology. Plant Physiol. Biochem. 2013, 72, 1–20. [Google Scholar] [CrossRef]
- Brauc, S.; De Vooght, E.; Claeys, M.; Hofte, M.; Angenon, G. Influence of over-expression of cytosolic aspartate aminotransferase on amino acid metabolism and defence responses against Botrytis cinerea infection in Arabidopsis thaliana. J. Plant Physiol. 2011, 168, 1813–1819. [Google Scholar] [CrossRef]
- Ngadze, E.; Icishahayo, D.; Coutinho, T.A.; van der Waals, J.E. Role of Polyphenol Oxidase, Peroxidase, Phenylalanine Ammonia Lyase, Chlorogenic Acid, and Total Soluble Phenols in Resistance of Potatoes to Soft Rot. Plant Dis. 2012, 96, 186–192. [Google Scholar] [CrossRef]
- Djami-Tchatchou, A.T.; Goszczynska, T.; Reddy, G.V.S.; Ntushelo, K. Gene expression response of Arabidopsis thaliana to inoculation with Pectobacterium carotovorum subsp. carotovorum. J. Plant Pathol. 2017, 99, 599–607. [Google Scholar] [CrossRef]
- Farahani, A.S.; Taghavi, S.M.; Afsharifar, A.; Niazi, A. Changes in expression of pathogenesis-related gene 1, pathogenesis-related gene 2, phenylalanine ammonialyase and catalase in tomato in response to Pectobacterium carotovorum subsp. carotovorum. J. Plant Pathol. 2016, 98, 525–530. [Google Scholar] [CrossRef]
- Song, Y.; Chen, D.; Lu, K.; Sun, Z.; Zeng, R. Enhanced tomato disease resistance primed by arbuscular mycorrhizal fungus. Front. Plant Sci. 2015, 6, 786. [Google Scholar] [CrossRef]
- Mikulic-Petkovsek, M.; Schmitzer, V.; Stampar, F.; Veberic, R.; Koron, D. Changes in phenolic content induced by infection with Didymella applanata and Leptosphaeria coniothyrium, the causal agents of raspberry spur and cane blight. Plant Pathol. 2014, 63, 185–192. [Google Scholar] [CrossRef]
- Chen, P.W.; Singh, P.; Zimmerli, L. Priming of the Arabidopsis pattern-triggered immunity response upon infection by necrotrophic Pectobacterium carotovorum bacteria. Mol. Plant Pathol. 2013, 14, 58–70. [Google Scholar] [CrossRef]
- Luzzatto, T.; Yishay, M.; Lipsky, A.; Ion, A.; Belausov, E.; Yedidia, I. Efficient, long-lasting resistance against the soft rot bacterium Pectobacterium carotovorum in calla lily provided by the plant activator methyl jasmonate. Plant Pathol. 2007, 56, 692–701. [Google Scholar] [CrossRef]
- Castiblanco, L.F.; Sundin, G.W. New insights on molecular regulation of biofilm formation in plant-associated bacteria. J. Integr. Plant Biol. 2016, 58, 362–372. [Google Scholar] [CrossRef]
- Jahn, C.E.; Selimi, D.A.; Barak, J.D.; Charkowski, A.O. The Dickeya dadantii biofilm matrix consists of cellulose nanofibres, and is an emergent property dependent upon the type III secretion system and the cellulose synthesis operon. Microbiology 2011, 157, 2733–2744. [Google Scholar] [CrossRef]
- Plyuta, V.; Zaitseva, J.; Lobakova, E.; Zagoskina, N.; Kuznetsov, A.; Khmel, I. Effect of plant phenolic compounds on biofilm formation by Pseudomonas aeruginosa. APMIS 2013, 121, 1073–1081. [Google Scholar] [CrossRef] [PubMed]
- Pollumaa, L.; Alamae, T.; Mae, A. Quorum sensing and expression of virulence in pectobacteria. Sensors 2012, 12, 3327–3349. [Google Scholar] [CrossRef]
- Barnard, A.M.L.; Salmond, G.P.C. Quorum sensing in Erwinia species. Anal. Bioanal. Chem. 2007, 387, 415–423. [Google Scholar] [CrossRef] [PubMed]
- Hernandez-Morales, A.; De la Torre-Zavala, S.; Ibarra-Laclette, E.; Hernandez-Flores, J.L.; Jofre-Garfias, A.E.; Martinez-Antonio, A.; Alvarez-Morales, A. Transcriptional profile of Pseudomonas syringae pv. phaseolicola NPS3121 in response to tissue extracts from a susceptible Phaseolus vulgaris L. cultivar. BMC Microbiol. 2009, 9, 257. [Google Scholar] [CrossRef] [PubMed]
- Watt, T.F.; Vucur, M.; Baumgarth, B.; Watt, S.A.; Niehaus, K. Low molecular weight plant extract induces metabolic changes and the secretion of extracellular enzymes, but has a negative effect on the expression of the type-III secretion system in Xanthomonas campestris pv. campestris. J. Biotechnol. 2009, 140, 59–67. [Google Scholar] [CrossRef] [PubMed]
- Saarilahti, H.T.; Pirhonen, M.; Karlsson, M.-B.; Flego, D.; Palva, E.T. Expression of pehA-bla gene fusions in Erwinia carotovora subsp. carotovora and isolation of regulatory mutants affecting polygalacturonase production. Molec. Gen. Genet. 1992, 234, 81–88. [Google Scholar] [CrossRef]
- Ham, J.H.; Cui, Y.; Alfano, J.R.; Rodriguez-Palenzuela, P.; Rojas, C.M.; Chatterjee, A.K.; Collmer, A. Analysis of Erwinia chrysanthemi EC16 pelE :: uidA, peIL :: uidA, and hrpN :: uidA mutants reveals strain-specific atypical regulation of the Hrp type III secretion system. Mol. Plant-Microbe Interact. 2004, 17, 184–194. [Google Scholar] [CrossRef] [PubMed]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Genes. | Forward Primer (5′→3′) | Reverse Primer (5′→3′) |
|---|---|---|
| A. PC16 and WPP14 | ||
| recA (reference gene) | CAG CAT CGA TGA ACG CAC AG | GGT TTA CCG ATG GGC CGT AT |
| pel | CTT CTT CAT GGC CGA TCC CA | CAA CGG ACT GTG GCT GAT TG |
| peh | TAC CGC TAC GAG TAC GAC GA | GAT CCC ACC AGC TCA CCT TT |
| expI | TAC AAT AGC GGC AGG CAC TC | TGA GAA TCA GGA AGC ATT GGC |
| expR | TGA GGT CAT GAG ATG TCG CC | TTA TGC CGT CGT AGC GAT CC |
| hrpL | AAC GTC GTC ATG GTT GCT GA | AAA CAC ATC GAA CCG ACC CC |
| araF | GGC TGA TTA TCG GTA TGA ACG | CCT GTT GCC TGA CCT TTG |
| B. Calla lily | ||
| actin (reference gene) | GAC TCA AAT CAT GTT AGA GAC ATT CAA | GTA CGG CCA CTG GCA TAG A |
| pal | GAC CTC GTC CCG CTC TCC TAC A | CTC CAC AGC AGA GAC GTG GTG AC |
| lox2 | CAT CAA GCT GCC AAG AGG TT | GCA ACC AAG AAA ATC CGT CT |
| ast | GCC AGT GGT GAT CCT GAG AG | GGA TGC TGA GGC AAC CTA CT |
| PR-1 | GGT AGA ACC TCT TCT GGG GAT G | AGT TGC TTC GGT AGT CGT AGT AC |
| C. Cabbage | ||
| tubulin (reference gene) | GGC TTC ACC ATT TAC CCT TC | TCC AAG AGG ACA GCA ACA TC |
| pal | ATC CCT AAC CGC CGA AGA G | TCC GCT AAC ACC GAT TGA AC |
| lox2 | GCT CTC CGT TCC ATA AAC CA | TAT TTG ACT GTG ATG CCG TGA |
| ast | TCC AAC CAC CAC AAC ATC TG | CAC ATC ATC CAT CAA TCC TTT G |
| PR-1 | GGA GAG AAC ATC GCT TGG AG | CAC ACA ACC TGC GTA TAG TGG |
| RT (min) | Identified Volatile Compounds | Calla lily Control | Calla lily + PC16 | Calla lily + WPP14 |
|---|---|---|---|---|
| 2.36 | Methyl acetate | + | ||
| 2.78 | Ethyl acetate | + | ||
| 3.80 | Isopentyl alcohol | + | ||
| 4.20 | 4-Methylheptane | + 1 | + | |
| 4.25 | Toluene | + | ||
| 4.28 | Isobutyl acetate | + | ||
| 5.09 | 2,4-Dimethyl-1-heptene | + | + | |
| 5.38 | 4-Methyloctane | + | ||
| 5.51 | Isopentyl acetate | + | ||
| 7.13 | Branched alkanes | + | + | + |
| 7.18 | Branched alkanes | + | + | + |
| 7.70 | Dibutoxy-methane | + | + | + |
| 7.96 | Branched alkanes | + | ||
| 8.01 | Branched alkanes | + | ||
| 10.09 | 2,6,10,14-tetramethyl heptadecane | + | + | + |
| 16.1 | Similar to sandaracopimaradiene | + |
| RT (min) | Identified Volatile Compounds | Cabbage Control | Cabbage + PC16 | Cabbage + WPP14 |
|---|---|---|---|---|
| 2.10 | Ethanol | + 1 | + | + |
| 2.78 | Ethyl acetate | + | + | + |
| 3.65 | 3-Hydroxy-2-butanone | + | + | + |
| 3.84 | Isopentyl alcohol | + | ||
| 4.0 | Methyl disulfide | + | ||
| 4.20 | 4-Methylheptane | + | + | + |
| 4.58 | Octane | + | + | + |
| 4.88 | 2,4-dimethylheptane | + | + | + |
| 5.10 | 2,4-dimethylheptene | + | + | + |
| 5.36 | 4-methyloctane | + | + | + |
| 7.13 | Branched alkanes | + | + | + |
| 7.18 | Branched alkanes | + | + | + |
| 7.71 | Dibutoxy-methane | + | + | + |
| 7.96 | Branched alkanes | + | + | |
| 8.01 | Branched alkanes | + | + | |
| 8.26 | Not identified | + | ||
| 8.34 | Not identified | + | ||
| 9.73 | Not identified | + | ||
| 10.09 | 2,6,10,14-tetramethyl heptadecane | + | + | |
| 10.37 | Not identified | + | ||
| 11.20 | Not identified | + |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Khadka, N.; Joshi, J.R.; Reznik, N.; Chriker, N.; Nudel, A.; Zelinger, E.; Kerem, Z.; Yedidia, I. Host Specificity and Differential Pathogenicity of Pectobacterium Strains from Dicot and Monocot Hosts. Microorganisms 2020, 8, 1479. https://doi.org/10.3390/microorganisms8101479
Khadka N, Joshi JR, Reznik N, Chriker N, Nudel A, Zelinger E, Kerem Z, Yedidia I. Host Specificity and Differential Pathogenicity of Pectobacterium Strains from Dicot and Monocot Hosts. Microorganisms. 2020; 8(10):1479. https://doi.org/10.3390/microorganisms8101479
Chicago/Turabian StyleKhadka, Nirmal, Janak Raj Joshi, Noam Reznik, Nofar Chriker, Adi Nudel, Einat Zelinger, Zohar Kerem, and Iris Yedidia. 2020. "Host Specificity and Differential Pathogenicity of Pectobacterium Strains from Dicot and Monocot Hosts" Microorganisms 8, no. 10: 1479. https://doi.org/10.3390/microorganisms8101479
APA StyleKhadka, N., Joshi, J. R., Reznik, N., Chriker, N., Nudel, A., Zelinger, E., Kerem, Z., & Yedidia, I. (2020). Host Specificity and Differential Pathogenicity of Pectobacterium Strains from Dicot and Monocot Hosts. Microorganisms, 8(10), 1479. https://doi.org/10.3390/microorganisms8101479