Schizosaccharomyces pombe Can Reduce Acetic Acid Produced by Baijiu Spontaneous Fermentation Microbiota


Abstract
1. Introduction
2. Materials and Methods
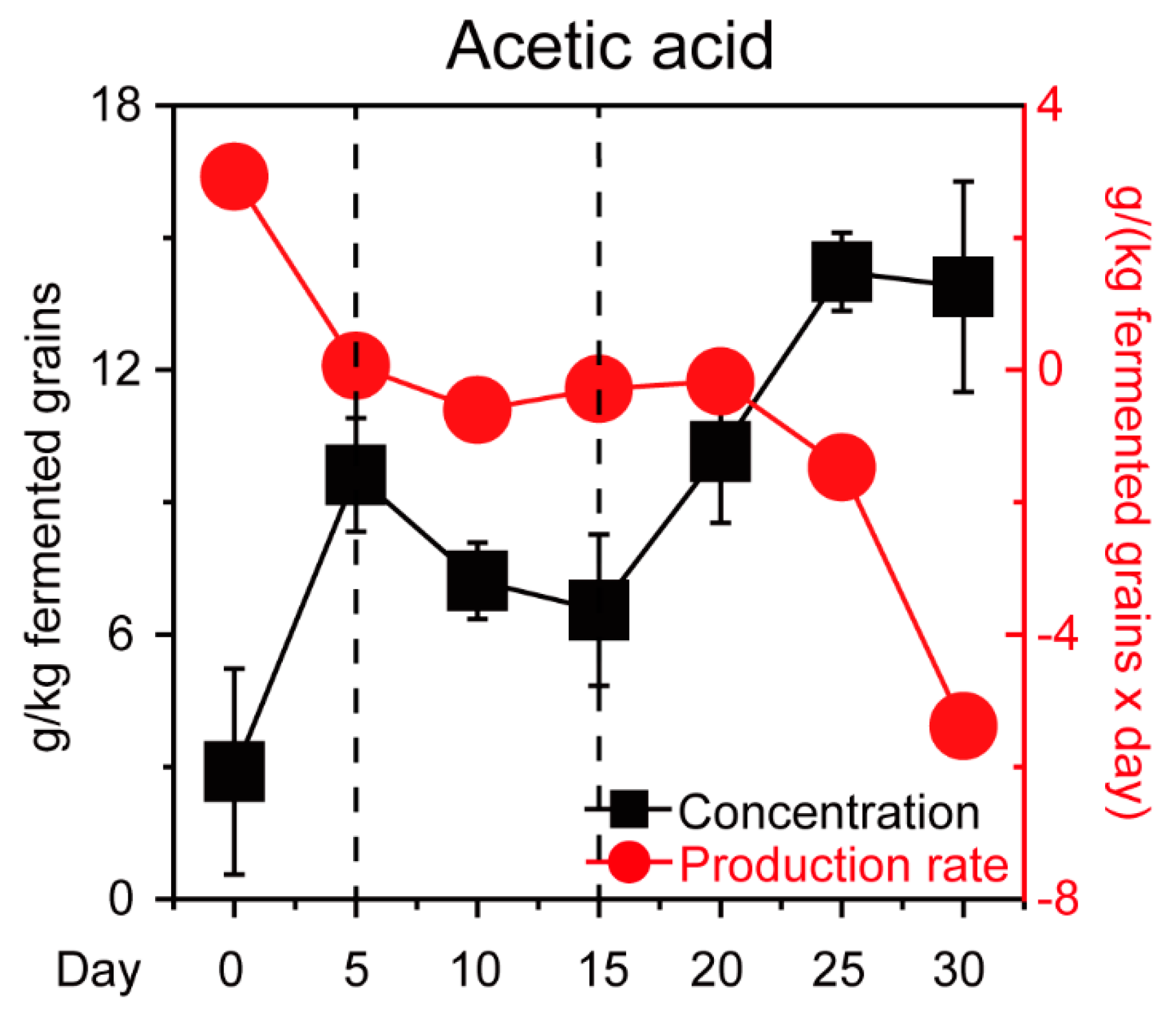
2.1. Experimental Design and Sample Collection
2.2. RNA Extraction, Metatranscriptomic Sequencing, and Analysis
2.3. Culture Fermentation
2.4. Reverse-Transcription and Real-Time Quantitative PCR
2.5. Metabolites Analysis
2.6. Statistical Analysis
3. Results
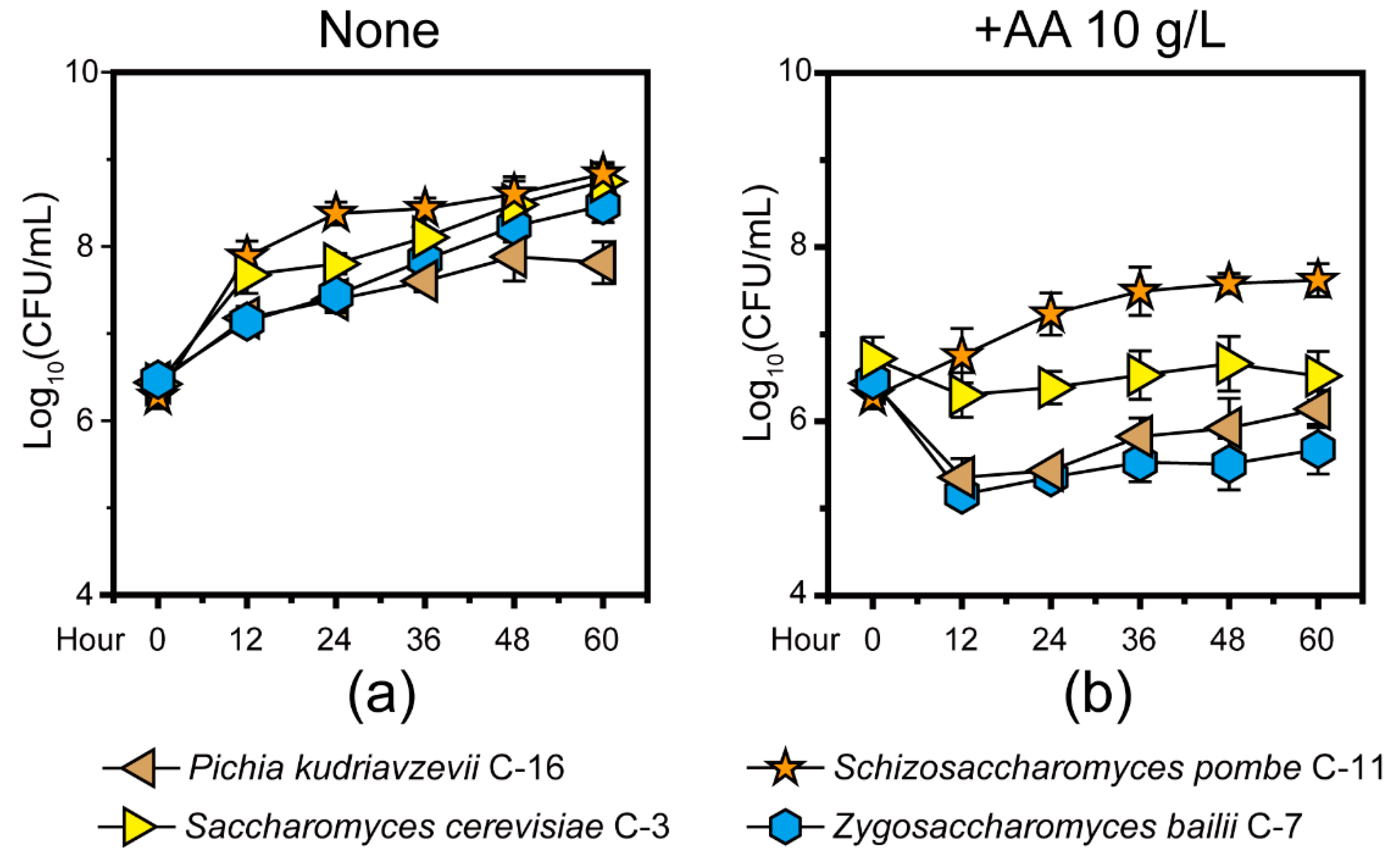
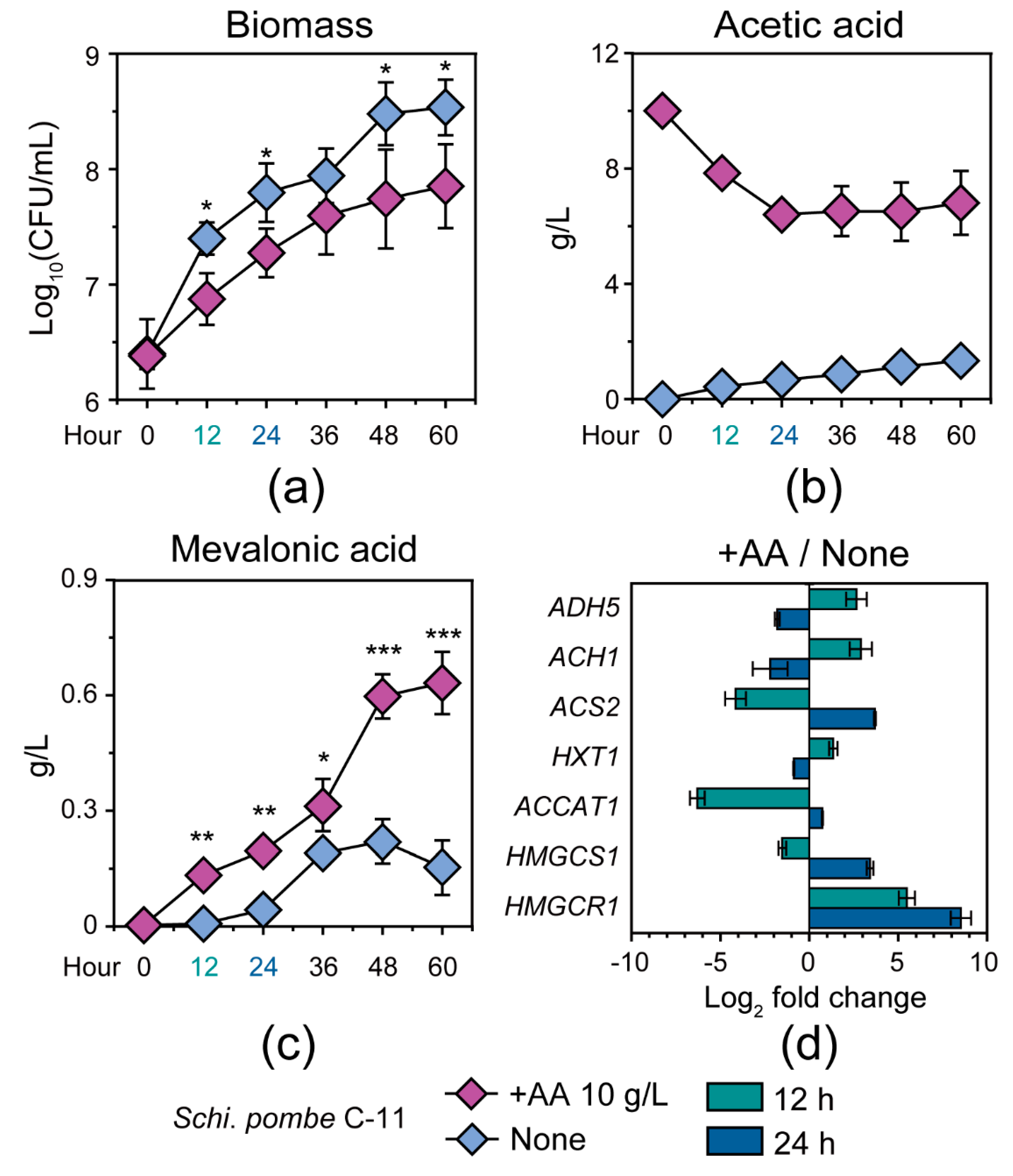
3.1. Potential Capacity of Non-Saccharomyces Yeast Schi. pombe in Response to Acetic Acid Stress
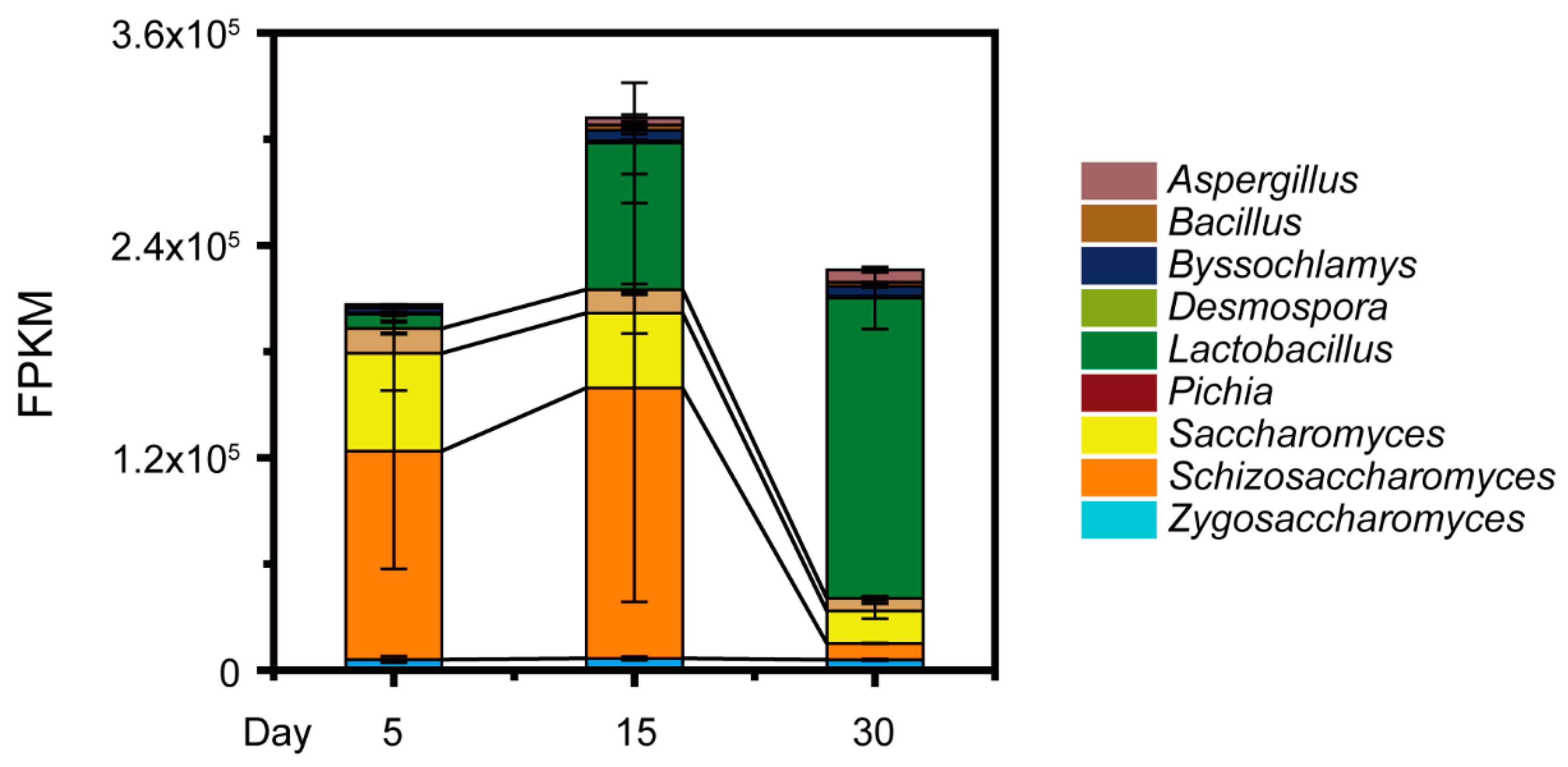
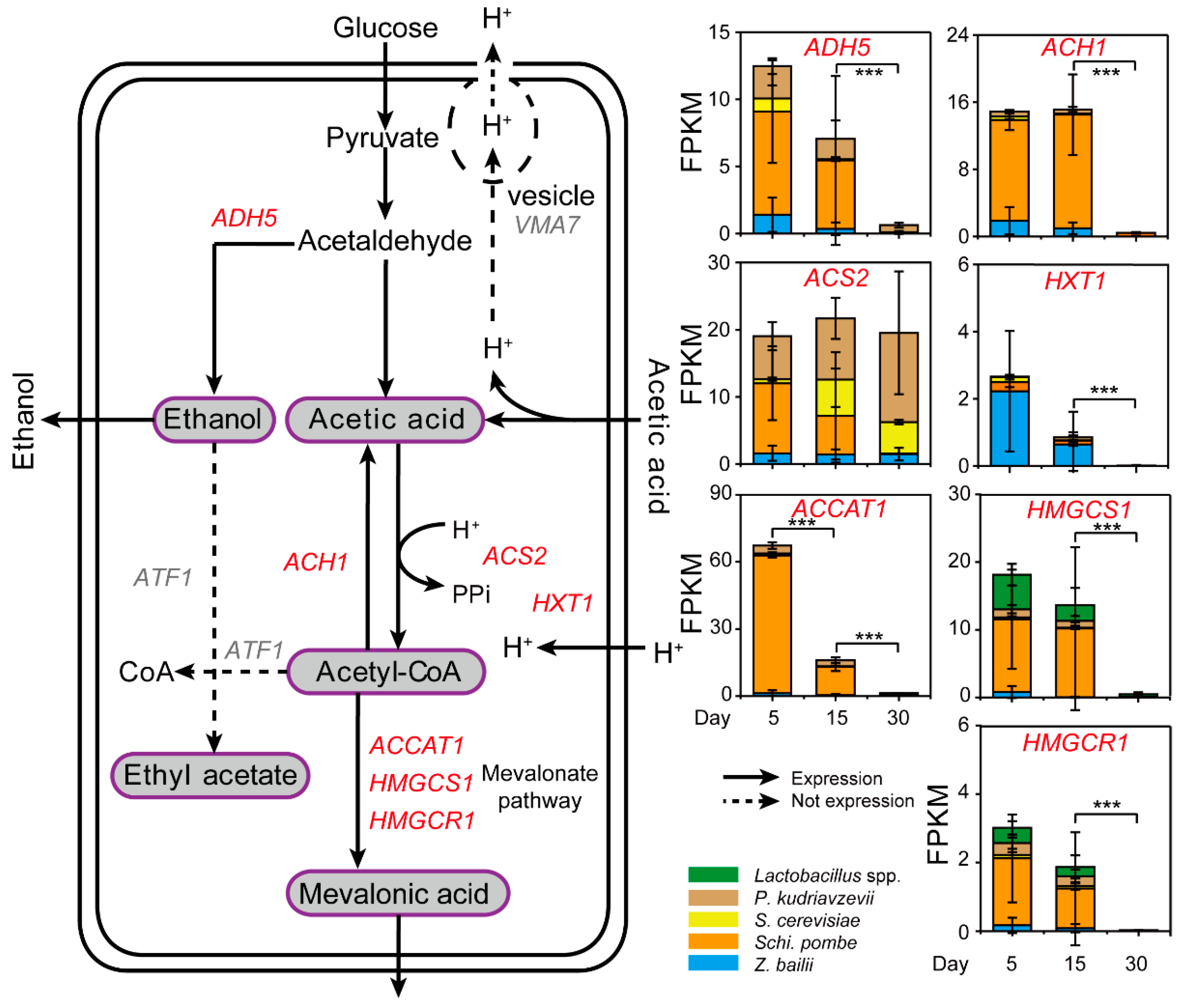
3.2. Schi. pombe Enhanced Conversion Efficiency of Acetic Acid to Generate Mevalonic Acid by Multiple Gene Upregulation
3.3. Response Mechanism of Schi. pombe to Acetic Acid Accumulation
4. Discussion
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Roca-Domenech, G.; Cordero-Otero, R.; Rozes, N.; Cleroux, M.; Pernet, A.; de Orduna, R.M. Metabolism of Schizosaccharomyces pombe under reduced osmotic stress conditions afforded by fed-batch alcoholic fermentation of white grape must. Food Res. Int. 2018, 113, 401–406. [Google Scholar] [CrossRef]
- Liu, S.; Laaksonen, O.; Kortesniemi, M.; Kalpio, M.; Yang, B. Chemical composition of bilberry wine fermented with non-Saccharomyces yeasts (Torulaspora delbrueckii and Schizosaccharomyces pombe) and Saccharomyces cerevisiae in pure, sequential and mixed fermentations. Food Chem. 2018, 266, 262–274. [Google Scholar] [CrossRef]
- Wei, J.; Zhang, Y.; Yuan, Y.; Dai, L.; Yue, T. Characteristic fruit wine production via reciprocal selection of juice and non-Saccharomyces species. Food Microbiol. 2019, 79, 66–74. [Google Scholar] [CrossRef] [PubMed]
- Du, H.; Wang, X.; Zhang, Y.; Xu, Y. Exploring the impacts of raw materials and environments on the microbiota in Chinese Daqu starter. Int. J. Food Microbiol. 2019, 297, 32–40. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.; Du, H.; Zhang, Y.; Xu, Y. Environmental microbiota drives microbial succession and metabolic profiles during Chinese liquor fermentation. Appl. Environ. Microbiol. 2018, 84. [Google Scholar] [CrossRef] [PubMed]
- Jin, G.; Zhu, Y.; Xu, Y. Mystery behind Chinese liquor fermentation. Trends Food Sci. Tech. 2017, 63, 18–28. [Google Scholar] [CrossRef]
- Zha, M.; Sun, B.; Wu, Y.; Yin, S.; Wang, C. Improving flavor metabolism of Saccharomyces cerevisiae by mixed culture with Wickerhamomyces anomalus for Chinese Baijiu making. J. Biosci. Bioeng. 2018, 126, 189–195. [Google Scholar] [CrossRef]
- Xu, Y.; Zhi, Y.; Wu, Q.; Du, R. Zygosaccharomyces bailii is a potential producer of various flavor compounds in Chinese Maotai-flavor liquor fermentation. Front. Microbiol. 2017, 8, 2609. [Google Scholar] [CrossRef]
- Song, Z.; Du, H.; Zhang, Y.; Xu, Y. Unraveling core functional microbiota in traditional solid-state fermentation by high-throughput amplicons and metatranscriptomics sequencing. Front. Microbiol. 2017, 8, 1294. [Google Scholar] [CrossRef]
- Wang, D.; Chen, L.; Yang, F.; Wang, H.; Wang, L. Yeasts and their importance to the flavour of traditional Chinese liquor: A review. J. Inst. Brew. 2019, 125, 214–221. [Google Scholar] [CrossRef]
- Jiang, J.; Liu, Y.; Li, H.; Yang, Q.; Wu, Q.; Chen, S.; Tang, J.; Xu, Y. Modeling and regulation of higher alcohol production through the combined effects of the C/N ratio and microbial interaction. J. Agric. Food Chem. 2019, 67, 10694–10701. [Google Scholar] [CrossRef] [PubMed]
- Fan, G.; Teng, C.; Xu, D.; Fu, Z.; Liu, P.; Wu, Q.; Yang, R.; Li, X. Improving ethyl acetate production in Baijiu manufacture by Wickerhamomyces anomalus and Saccharomyces cerevisiae mixed culture fermentations. Biomed Res. Int. 2019, 2019, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Du, H.; Song, Z.; Xu, Y. Ethyl carbamate formation regulated by lactic acid bacteria and nonconventional yeasts in solid-state fermentation of Chinese Moutai-flavor liquor. J Agric. Food Chem. 2017, 66, 387–392. [Google Scholar] [CrossRef] [PubMed]
- Xu, Y.; Ji, K. Moutai (Maotai): Production and sensory properties. In Alcoholic Beverages; John, P., Ed.; Woodhead Publishing: London, UK, 2012; pp. 315–330. [Google Scholar]
- Wu, Q.; Xu, Y.; Chen, L. Diversity of yeast species during fermentative process contributing to Chinese Maotai-flavour liquor making. Lett. Appl. Microbiol. 2012, 55, 301–307. [Google Scholar] [CrossRef] [PubMed]
- Su, J.; Wang, T.; Wang, Y.; Li, Y.Y.; Li, H. The use of lactic acid-producing, malic acid-producing, or malic acid-degrading yeast strains for acidity adjustment in the wine industry. Appl. Microbiol. Biotechnol. 2014, 98, 2395–2413. [Google Scholar] [CrossRef]
- Benito, A.; Calderon, F.; Benito, S. Schizosaccharomyces pombe isolation protocol. In Methods Molecular Biology, 1st ed.; Singleton, T., Ed.; Springer: Berlin/Heidelberg, Germany, 2018; pp. 227–234. [Google Scholar]
- Benito, S. The impacts of Schizosaccharomyces on winemaking. Appl. Microbiol. Biotechnol. 2019, 103, 4291–4312. [Google Scholar] [CrossRef]
- Wu, Q.; Chen, L.; Xu, Y. Yeast community associated with the solid-state fermentation of traditional Chinese Maotai-flavor liquor. Int. J. Food Microbiol. 2013, 166, 323–330. [Google Scholar] [CrossRef]
- Wu, Q.; Chen, B.; Xu, Y. Regulating yeast flavor metabolism by controlling saccharification reaction rate in simultaneous saccharification and fermentation of Chinese Maotai-flavor liquor. Int. J. Food Microbiol. 2015, 200, 39–46. [Google Scholar] [CrossRef]
- Zhang, J.; Presley, G.N.; Hammel, K.E.; Ryu, J.S.; Menke, J.R.; Figueroa, M.; Hu, D.; Orr, G.; Schilling, J.S. Localizing gene regulation reveals a staggered wood decay mechanism for the brown rot fungus Postia placenta. Proc. Natl. Acad. Sci. USA 2016, 113, 10968–10973. [Google Scholar] [CrossRef]
- Azvolinsky, A.; DeFrancesco, L.; Waltz, E.; Webb, S. 20 years of Nature Biotechnology research tools. Nature Biotech. 2016, 34, 256. [Google Scholar] [CrossRef]
- Louis, S.; Tappu, R.M.; Damms-Machado, A.; Huson, D.H.; Bischoff, S.C. Characterization of the gut microbial community of obese patients following a weight-loss intervention using whole metagenome shotgun sequencing. PLoS ONE 2016, 11, e0149564. [Google Scholar] [CrossRef] [PubMed]
- Buchfink, B.; Xie, C.; Huson, D.H. Fast and sensitive protein alignment using DIAMOND. Nat. Methods 2015, 12, 59. [Google Scholar] [CrossRef] [PubMed]
- Burstein, D.; Harrington, L.B.; Strutt, S.C.; Probst, A.J.; Anantharaman, K.; Thomas, B.C.; Doudna, J.A.; Banfield, J.F. New CRISPR-Cas systems from uncultivated microbes. Nature 2017, 542, 237. [Google Scholar] [CrossRef] [PubMed]
- Kanehisa, M.; Furumichi, M.; Tanabe, M.; Sato, Y.; Morishima, K. KEGG: New perspectives on genomes, pathways, diseases and drugs. Nucleic Acids Res. 2016, 45, D353–D361. [Google Scholar] [CrossRef]
- Abergel, A.; Metivier, S.; Samuel, D.; Jiang, D.; Kersey, K.; Pang, P.S.; Svarovskaia, E.; Knox, S.J.; Loustaud-Ratti, V.; Asselah, T. Ledipasvir plus sofosbuvir for 12 weeks in patients with hepatitis C genotype 4 infection. Hepatology 2016, 64, 1049–1056. [Google Scholar]
- Meng, X.; Wu, Q.; Wang, L.; Wang, D.; Chen, L.; Xu, Y. Improving flavor metabolism of Saccharomyces cerevisiae by mixed culture with Bacillus licheniformis for Chinese Maotai-flavor liquor making. J. Ind. Microbiol. Biotechnol. 2015, 42, 1601–1608. [Google Scholar] [CrossRef]
- Wang, X.; Jiang, D.; Yang, D. Fast-tracking determination of homozygous transgenic lines and transgene stacking using a reliable quantitative real-time PCR assay. Appl. Biochem. Biotechnol. 2015, 175, 996–1006. [Google Scholar] [CrossRef]
- Ye, J.; Coulouris, G.; Zaretskaya, I.; Cutcutache, I.; Rozen, S.; Madden, T.L. Primer-BLAST: A tool to design target-specific primers for polymerase chain reaction. BMC Bioinform. 2012, 13, 134. [Google Scholar] [CrossRef]
- Zott, K.; Claisse, O.; Lucas, P.; Coulon, J.; Lonvaud-Funel, A.; Masneuf-Pomarede, I. Characterization of the yeast ecosystem in grape must and wine using real-time PCR. Food Microbiol. 2010, 27, 559–567. [Google Scholar] [CrossRef]
- Rodriguez, S.; Denby, C.M.; Van Vu, T.; Baidoo, E.E.; Wang, G.; Keasling, J.D. ATP citrate lyase mediated cytosolic acetyl-CoA biosynthesis increases mevalonate production in Saccharomyces cerevisiae. Microb. Cell Fac. 2016, 15, 48. [Google Scholar] [CrossRef]
- Benito, S.; Palomero, F.; Calderón, F.; Palmero, D.; Suárez-Lepe, J.A. Selection of appropriate Schizosaccharomyces strains for winemaking. Food Microbiol. 2014, 42, 218–224. [Google Scholar] [CrossRef] [PubMed]
- Benito, A.; Jeffares, D.; Palomero, F.; Calderon, F.; Bai, F.; Bahler, J.; Benito, S. Selected Schizosaccharomyces pombe strains have characteristics that are beneficial for winemaking. PLoS ONE 2016, 11, e0151102. [Google Scholar] [CrossRef] [PubMed]
- Geng, P.; Zhang, L.; Shi, G.Y. Omics analysis of acetic acid tolerance in Saccharomyces cerevisiae. World J. Microbiol. Biotechnol. 2017, 33, 94. [Google Scholar] [CrossRef]
- Meijnen, J.P.; Randazzo, P.; Foulquie-Moreno, M.R.; van den Brink, J.; Vandecruys, P.; Stojiljkovic, M.; Dumortier, F.; Zalar, P.; Boekhout, T.; Gunde-Cimerman, N.; et al. Polygenic analysis and targeted improvement of the complex trait of high acetic acid tolerance in the yeast Saccharomyces cerevisiae. Biotechnol. Biofuels 2016, 9, 5. [Google Scholar] [CrossRef]
- Rodrigues, F.; Zeeman, A.M.; Cardoso, H.; Sousa, M.J.; Steensma, H.Y.; Côrte-Real, M.; Leão, C. Isolation of an acetyl-CoA synthetase gene (ZbACS2) from Zygosaccharomyces bailii. Yeast 2004, 21, 325–331. [Google Scholar] [CrossRef]
- Rodrigues, F.; Sousa, M.J.; Ludovico, P.; Santos, H.; Côrte-Real, M.; Leão, C. The fate of acetic acid during glucose co-metabolism by the spoilage yeast Zygosaccharomyces bailii. PLoS ONE 2012, 7, e52402. [Google Scholar] [CrossRef]
- Kruis, A.J.; Mars, A.E.; Kengen, S.W.; Borst, J.W.; van der Oost, J.; Weusthuis, R.A. Alcohol acetyltransferase Eat1 is located in yeast mitochondria. Appl. Environ. Microbiol. 2018, 84. [Google Scholar] [CrossRef]
- Mason, A.B.; Dufour, J.P. Alcohol acetyltransferases and the significance of ester synthesis in yeast. Yeast 2000, 16, 1287–1298. [Google Scholar] [CrossRef]
- Orij, R.; Brul, S.; Smits, G.J. Intracellular pH is a tightly controlled signal in yeast. BBA Gen. Subj. 2011, 1810, 933–944. [Google Scholar] [CrossRef]
- Ding, B.J.; Lager, I.; Bansal, S.; Durrett, T.P.; Stymne, S.; Löfstedt, C. The yeast ATF1 acetyltransferase efficiently acetylates insect pheromone alcohols: Implications for the biological production of moth pheromones. Lipids 2016, 51, 469–475. [Google Scholar] [CrossRef]
- Dirkmann, M.; Nowack, J.; Schulz, F. An in vitro biosynthesis of sesquiterpenes starting from acetic acid. ChemBioChem 2018, 19, 2146–2151. [Google Scholar] [CrossRef] [PubMed]
- Sun, W.; Qin, L.; Xue, H.; Yu, Y.; Ma, Y.; Wang, Y.; Li, C. Novel trends for producing plant triterpenoids in yeast. Crit. Rev. Biotechnol. 2019, 39, 618–632. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Primer Name | Sequence (5′-3′) | Primer Size (bp) | Reference |
|---|---|---|---|
| PkF | GTTTGAGCGTCGTTTCCATC | 20 | [31] |
| PkR | AGCTCCGACGCTCTTTACAC | 20 | |
| ScF | GTGCGCGGTCTTGCTAGGCT | 20 | this work |
| ScR | TACCTCTGGGCCCCGATTGC | 20 | |
| SpF | AGTGAAGCGGGAAAAGCTCA | 20 | this work |
| SpR | ATCGACCAAAGACGGGGTTC | 20 | |
| ZbF | CATGGTGTTTTGCGCC | 16 | [8] |
| ZbR | CGTCCGCCACGAAGTGGTAGA | 21 |
| Primer Name | Sequence (5′-3′) | Primer Size (bp) | Reference |
|---|---|---|---|
| ACH1SpF | TGCATACACCATCGGTTCGT | 20 | this work |
| ACH1SpR | GAGCACGTTCGGTAGGAGAC | 20 | |
| ACCAT1SpF | GCTTCTCTTCCTGCCACCAA | 20 | this work |
| ACCAT1SpR | TGGCCAAGATTGGCTGAGAC | 20 | |
| ACS2SpF | AGAGTCTGTTGCAGACCGTG | 20 | this work |
| ACS2SpR | CATACTGGCGGTAGGCTCAG | 20 | |
| ADH5SpF | AGTTGGATCCATGGGTGCTT | 20 | this work |
| ADH5SpR | TTCCGGTTTCGCTTCAGCAT | 20 | |
| HMGCS1SpF | GTGGCGTGAACGCTCTTTTT | 20 | this work |
| HMGCS1SpR | GGGGCATTAGGACCAACCAA | 20 | |
| HMGCR1SpF | AGAGGTCGGCAATTGGACTG | 20 | this work |
| HMGCR1SpR | AGTGCGAGCGATCAACTGAA | 20 | |
| HXT1SpF | GCCGGTACTGTGAAAAGGGA | 20 | this work |
| HXT1SpR | TCAGTTTGGATTGATGCGCTG | 21 | |
| UBC6SpF | TTGGCTGTTGCCATCCTTTG | 20 | this work |
| UBC6SpR | GGAAACGTCCGCTTGGAGTA | 20 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Song, Z.; Du, H.; Zhang, M.; Nie, Y.; Xu, Y. Schizosaccharomyces pombe Can Reduce Acetic Acid Produced by Baijiu Spontaneous Fermentation Microbiota. Microorganisms 2019, 7, 606. https://doi.org/10.3390/microorganisms7120606
Song Z, Du H, Zhang M, Nie Y, Xu Y. Schizosaccharomyces pombe Can Reduce Acetic Acid Produced by Baijiu Spontaneous Fermentation Microbiota. Microorganisms. 2019; 7(12):606. https://doi.org/10.3390/microorganisms7120606
Chicago/Turabian StyleSong, Zhewei, Hai Du, Menghui Zhang, Yao Nie, and Yan Xu. 2019. "Schizosaccharomyces pombe Can Reduce Acetic Acid Produced by Baijiu Spontaneous Fermentation Microbiota" Microorganisms 7, no. 12: 606. https://doi.org/10.3390/microorganisms7120606
APA StyleSong, Z., Du, H., Zhang, M., Nie, Y., & Xu, Y. (2019). Schizosaccharomyces pombe Can Reduce Acetic Acid Produced by Baijiu Spontaneous Fermentation Microbiota. Microorganisms, 7(12), 606. https://doi.org/10.3390/microorganisms7120606