
Alleviation of Adipose Tissue Inflammation and Obesity Suppression by a Probiotic Strain That Induces GLP-1 Secretion
 ,
,
Abstract
1. Introduction
2. Materials and Methods
2.1. Preparation of Cell-Free Culture Supernatant (CFS) of Lactic Acid Bacteria
2.2. In Vitro Assay for GLP-1 Secretion
2.3. RNA Extraction and Quantitative RT-PCR (qRT-PCR) Analysis
2.4. Acid Resistance Test
2.5. HFD-Induced Mouse Model of Obesity
2.6. Glucose Tolerance Test (GTT)
2.7. Insulin Tolerance Test (ITT)
2.8. Measurement of Metabolic Hormone Levels in Serum
2.9. Histological Analysis
2.10. Isolation of Immune Cells in Adipose Tissue
2.11. Immune Cell Analysis by Flow Cytometry
2.12. Statistical Analysis
3. Results
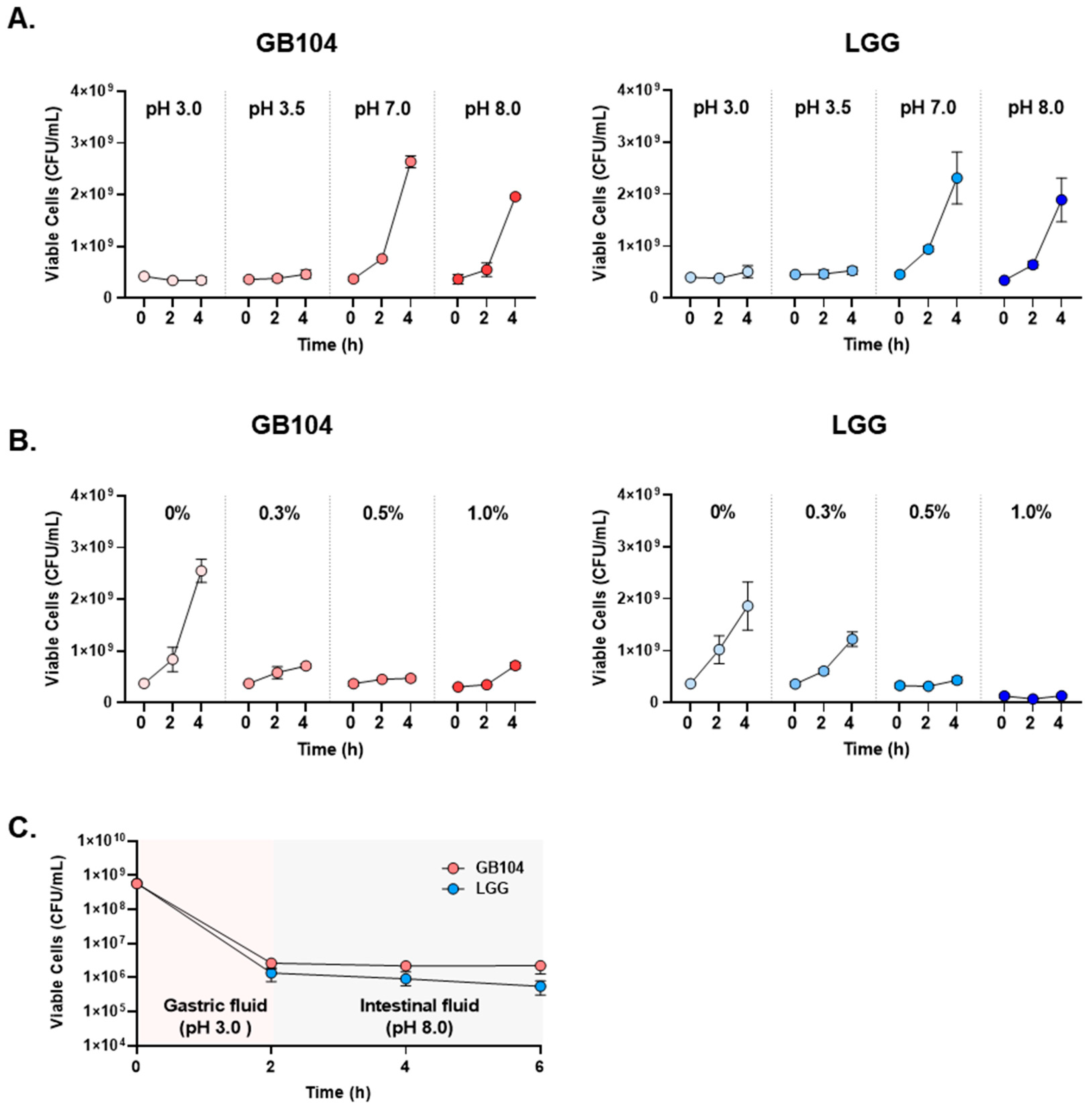
3.1. GB104 Promotes GLP-1 Secretion from Enteroendocrine Cells and Exhibits Similar Acid Resistance to LGG
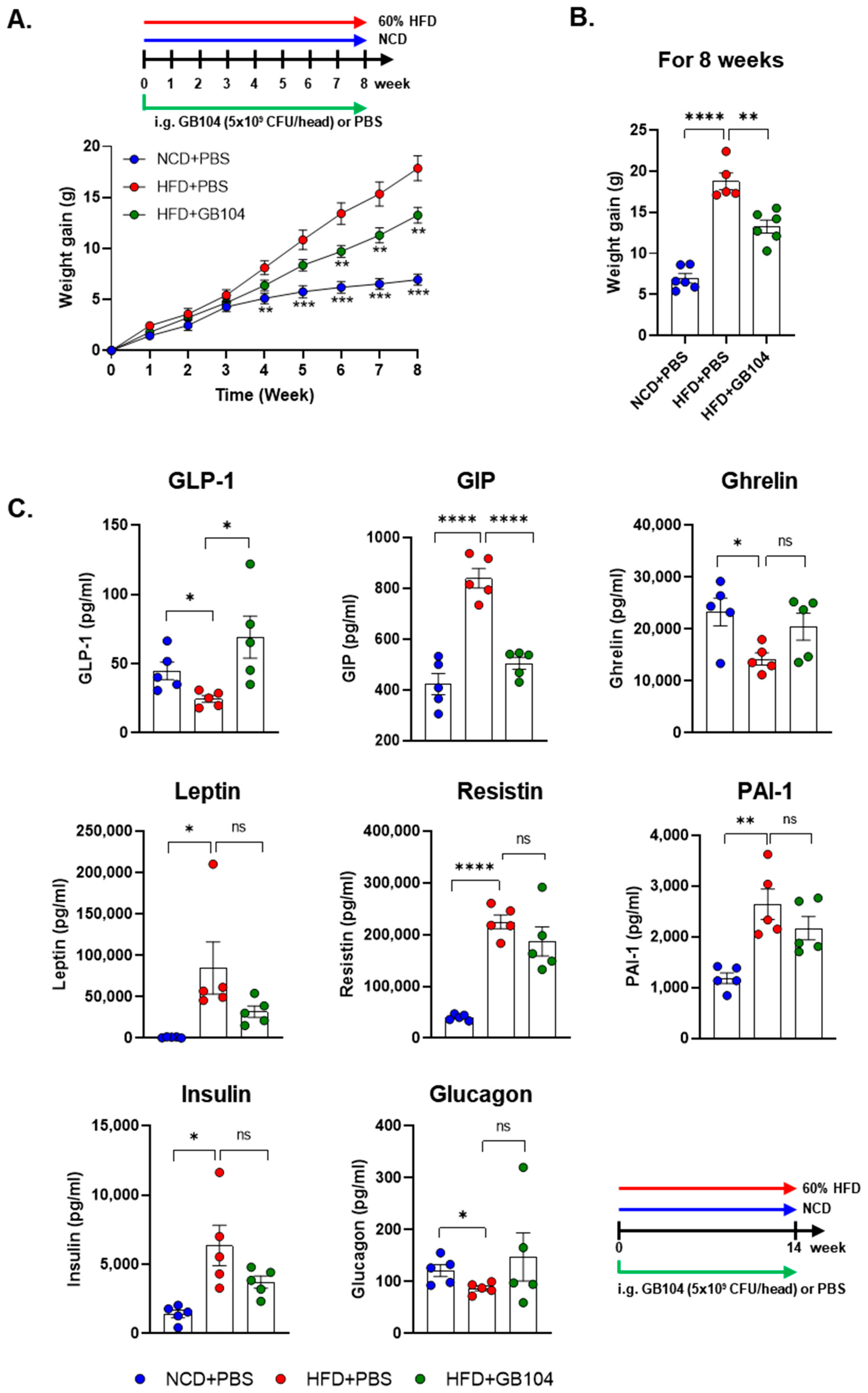
3.2. GB104 Increases Blood GLP-1 Levels and Inhibits Weight Gain in an HFD-Induced Mouse Model of Obesity
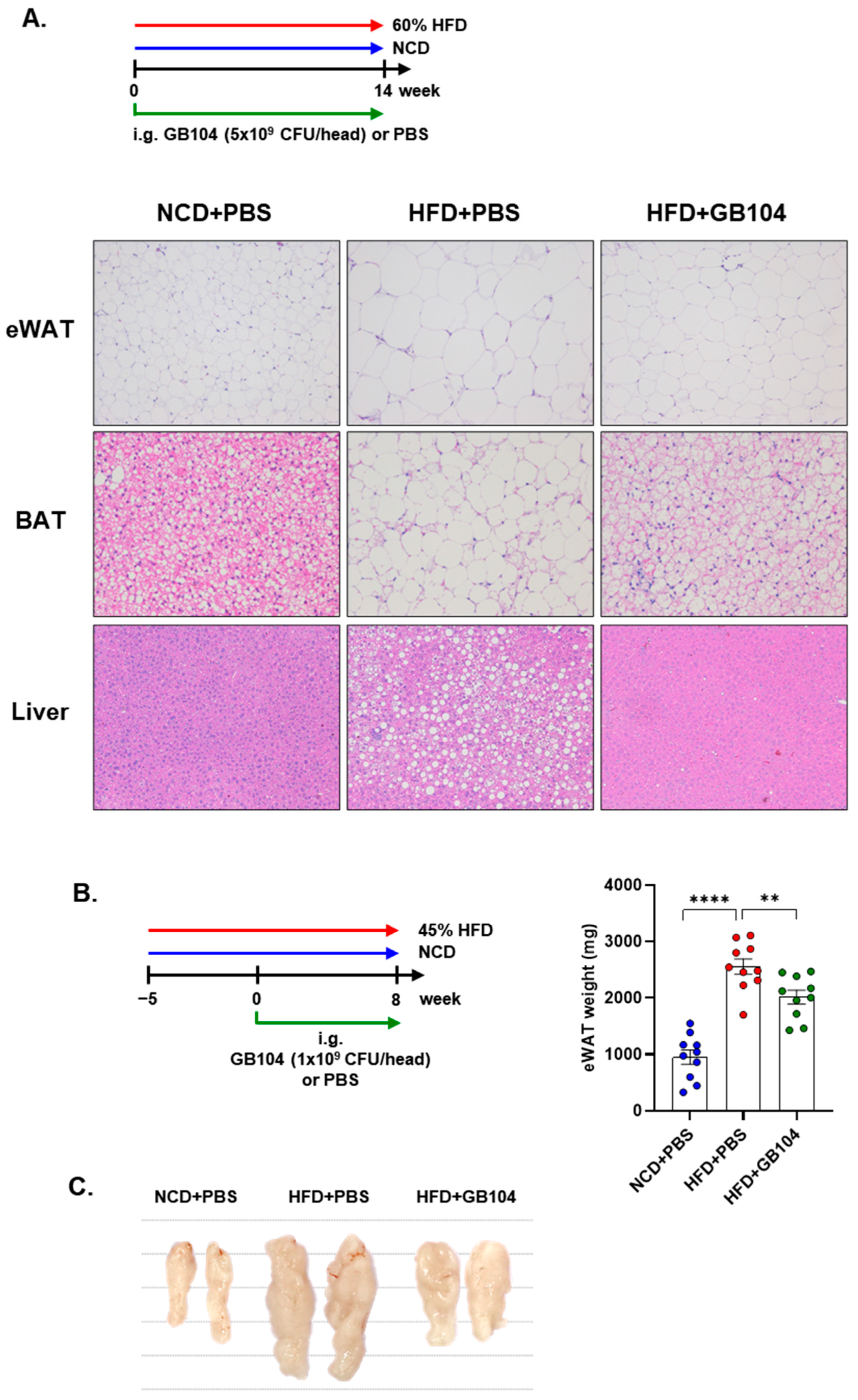
3.3. GB104 Improves Blood Glucose Levels and Suppresses Fat Accumulation
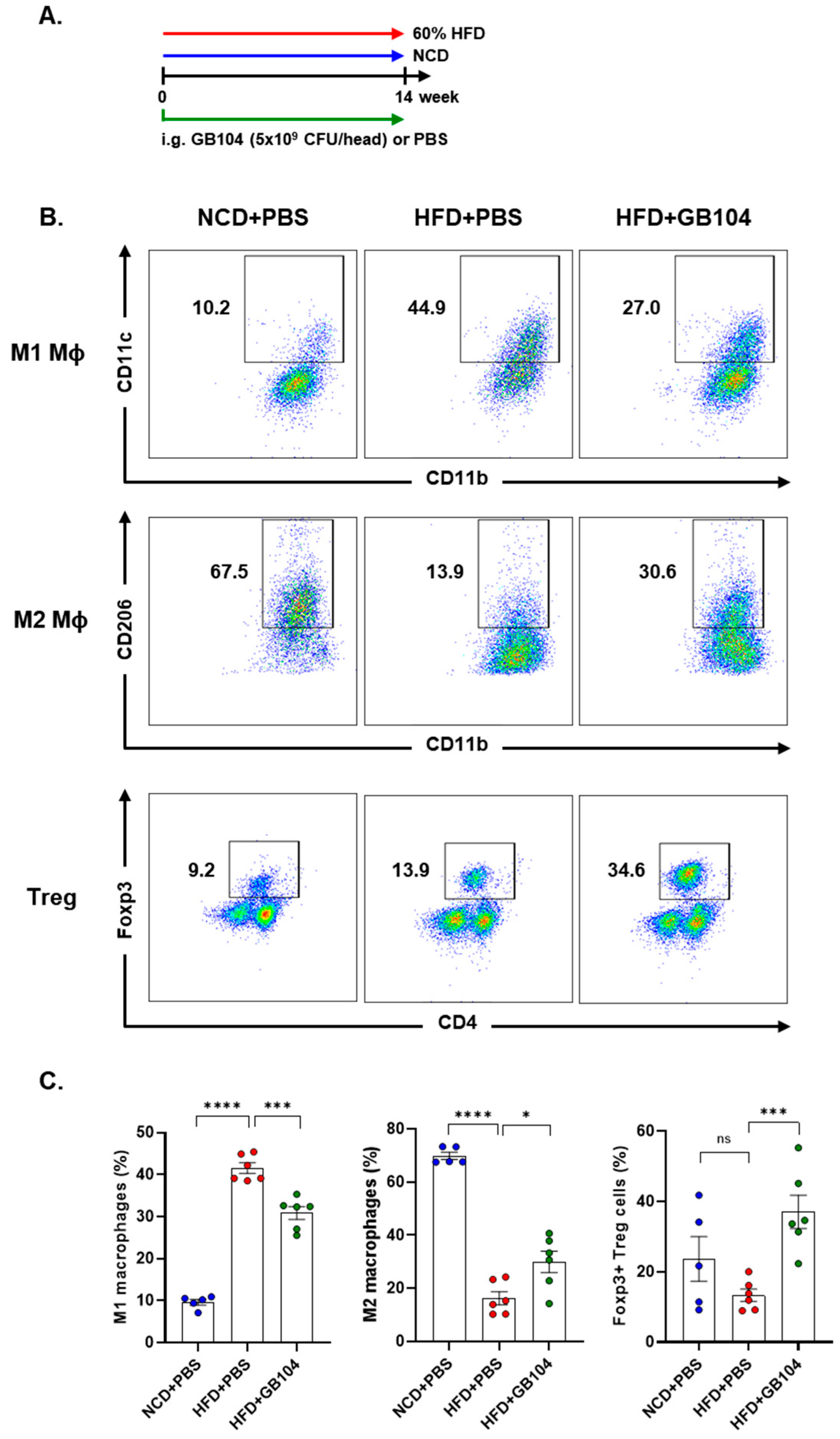
3.4. GB104 Alleviates Adipose Tissue Inflammation Induced by HFD
4. Discussion
5. Conclusions
6. Patents
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
| GLP-1 | glucagon-like peptide-1 |
| HFD | high-fat diet |
| GLP-1RA | GLP-1 receptor agonist |
| GPCR | G protein coupled receptor |
| GCG | proglucagon gene |
| PCSK | proprotein convertase subtilisin/kexin |
| GIP | gastric inhibitory polypeptide |
| Treg | regulatory T cells |
| CNS | central nervous system |
| NTS | nucleus tractus solitary |
| CSF | cell-free culture supernatant |
| GAPDH | glyceraldehyde-3-phosphate dehydrogenase |
| ATCC | American Type Culture Collection |
| KCTC | Korean Collection for Type Cultures |
| GTT | glucose tolerance test |
| ITT | insulin tolerance test |
| NBF | neutral buffered formalin |
| H&E | hematoxylin and eosin |
| NEAA | non-essential amino acids |
| EDTA | ethylenediaminetetraacetic acid |
| MHC | major histocompatibility complex |
References
- Rohm, T.V.; Meier, D.T.; Olefsky, J.M.; Donath, M.Y. Inflammation in obesity, diabetes, and related disorders. Immunity 2022, 55, 31–55. [Google Scholar] [CrossRef] [PubMed]
- Fujisaka, S.; Usui, I.; Bukhari, A.; Ikutani, M.; Oya, T.; Kanatani, Y.; Tsuneyama, K.; Nagai, Y.; Takatsu, K.; Urakaze, M.; et al. Regulatory mechanisms for adipose tissue M1 and M2 macrophages in diet-induced obese mice. Diabetes 2009, 58, 2574–2582. [Google Scholar] [CrossRef] [PubMed]
- Russo, L.; Lumeng, C.N. Properties and functions of adipose tissue macrophages in obesity. Immunology 2018, 155, 407–417. [Google Scholar] [CrossRef] [PubMed]
- Feuerer, M.; Herrero, L.; Cipolletta, D.; Naaz, A.; Wong, J.; Nayer, A.; Lee, J.; Goldfine, A.B.; Benoist, C.; Shoelson, S.; et al. Lean, but not obese, fat is enriched for a unique population of regulatory T cells that affect metabolic parameters. Nat. Med. 2009, 15, 930–939. [Google Scholar] [CrossRef]
- Jung, U.J.; Choi, M.S. Obesity and its metabolic complications: The role of adipokines and the relationship between obesity, inflammation, insulin resistance, dyslipidemia and nonalcoholic fatty liver disease. Int. J. Mol. Sci. 2014, 15, 6184–6223. [Google Scholar] [CrossRef]
- Ouchi, N.; Parker, J.L.; Lugus, J.J.; Walsh, K. Adipokines in inflammation and metabolic disease. Nat. Rev. Immunol. 2011, 11, 85–97. [Google Scholar] [CrossRef]
- Wang, Q.; Perrard, X.D.; Perrard, J.L.; Mansoori, A.; Raya, J.L.; Hoogeveen, R.; Smith, C.W.; Ballantyne, C.M.; Wu, H. Differential effect of weight loss with low-fat diet or high-fat diet restriction on inflammation in the liver and adipose tissue of mice with diet-induced obesity. Atherosclerosis 2011, 219, 100–108. [Google Scholar] [CrossRef]
- Gribble, F.M.; Reimann, F. Function and mechanisms of enteroendocrine cells and gut hormones in metabolism. Nat. Rev. Endocrinol. 2019, 15, 226–237. [Google Scholar] [CrossRef]
- Friedman, J.M. Leptin and the endocrine control of energy balance. Nat. Metab. 2019, 1, 754–764. [Google Scholar] [CrossRef]
- Maffei, M.; Halaas, J.; Ravussin, E.; Pratley, R.E.; Lee, G.H.; Zhang, Y.; Fei, H.; Kim, S.; Lallone, R.; Ranganathan, S.; et al. Leptin levels in human and rodent: Measurement of plasma leptin and ob RNA in obese and weight-reduced subjects. Nat. Med. 1995, 1, 1155–1161. [Google Scholar] [CrossRef]
- Briggs, D.I.; Enriori, P.J.; Lemus, M.B.; Cowley, M.A.; Andrews, Z.B. Diet-induced obesity causes ghrelin resistance in arcuate NPY/AgRP neurons. Endocrinology 2010, 151, 4745–4755. [Google Scholar] [CrossRef] [PubMed]
- Seino, Y.; Fukushima, M.; Yabe, D. GIP and GLP-1, the two incretin hormones: Similarities and differences. J. Diabetes Investig. 2010, 1, 8–23. [Google Scholar] [CrossRef] [PubMed]
- Yamane, S.; Harada, N.; Inagaki, N. Mechanisms of fat-induced gastric inhibitory polypeptide/glucose-dependent insulinotropic polypeptide secretion from K cells. J. Diabetes Investig. 2016, 7 (Suppl. 1), 20–26. [Google Scholar] [CrossRef] [PubMed]
- Haedersdal, S.; Lund, A.; Knop, F.K.; Vilsboll, T. The Role of Glucagon in the Pathophysiology and Treatment of Type 2 Diabetes. Mayo Clin. Proc. 2018, 93, 217–239. [Google Scholar] [CrossRef]
- Avtanski, D.; Pavlov, V.A.; Tracey, K.J.; Poretsky, L. Characterization of inflammation and insulin resistance in high-fat diet-induced male C57BL/6J mouse model of obesity. Anim. Model. Exp. Med. 2019, 2, 252–258. [Google Scholar] [CrossRef]
- Zheng, Z.; Zong, Y.; Ma, Y.; Tian, Y.; Pang, Y.; Zhang, C.; Gao, J. Glucagon-like peptide-1 receptor: Mechanisms and advances in therapy. Signal Transduct. Target. Ther. 2024, 9, 234. [Google Scholar] [CrossRef]
- Drucker, D.J. Mechanisms of Action and Therapeutic Application of Glucagon-like Peptide-1. Cell Metab. 2018, 27, 740–756. [Google Scholar] [CrossRef]
- Muller, T.D.; Finan, B.; Bloom, S.R.; D’Alessio, D.; Drucker, D.J.; Flatt, P.R.; Fritsche, A.; Gribble, F.; Grill, H.J.; Habener, J.F.; et al. Glucagon-like peptide 1 (GLP-1). Mol. Metab. 2019, 30, 72–130. [Google Scholar] [CrossRef]
- Baggio, L.L.; Drucker, D.J. Glucagon-like peptide-1 receptors in the brain: Controlling food intake and body weight. J. Clin. Investig. 2014, 124, 4223–4226. [Google Scholar] [CrossRef]
- Andersen, A.; Lund, A.; Knop, F.K.; Vilsboll, T. Glucagon-like peptide 1 in health and disease. Nat. Rev. Endocrinol. 2018, 14, 390–403. [Google Scholar] [CrossRef]
- Tran, S.; Kramer, C.K.; Zinman, B.; Choi, H.; Retnakaran, R. Effect of chronic liraglutide therapy and its withdrawal on time to postchallenge peak glucose in type 2 diabetes. Am. J. Physiol. Endocrinol. Metab. 2018, 314, E287–E295. [Google Scholar] [CrossRef] [PubMed]
- Wilding, J.P.H.; Batterham, R.L.; Davies, M.; Van Gaal, L.F.; Kandler, K.; Konakli, K.; Lingvay, I.; McGowan, B.M.; Oral, T.K.; Rosenstock, J.; et al. Weight regain and cardiometabolic effects after withdrawal of semaglutide: The STEP 1 trial extension. Diabetes Obes. Metab. 2022, 24, 1553–1564. [Google Scholar] [CrossRef] [PubMed]
- Han, H.; Yi, B.; Zhong, R.; Wang, M.; Zhang, S.; Ma, J.; Yin, Y.; Yin, J.; Chen, L.; Zhang, H. From gut microbiota to host appetite: Gut microbiota-derived metabolites as key regulators. Microbiome 2021, 9, 162. [Google Scholar] [CrossRef] [PubMed]
- Kang, C.H.; Jeong, Y.A.; Han, S.H.; Kim, J.S.; Kim, Y.; Park, H.M.; Choi, S.I.; Paek, N.S. In vitro Probiotic Evaluation of Potential Antiobesity Lactic Acid Bacteria Isolated from Human Vagina and Shellfish. Korean Soc. Biotechnol. Bioeng. 2018, 33, 161–167. [Google Scholar] [CrossRef]
- Kim, J.H.; Jeun, E.J.; Hong, C.P.; Kim, S.H.; Jang, M.S.; Lee, E.J.; Moon, S.J.; Yun, C.H.; Im, S.H.; Jeong, S.G.; et al. Extracellular vesicle-derived protein from Bifidobacterium longum alleviates food allergy through mast cell suppression. J. Allergy Clin. Immunol. 2016, 137, 507–516.e8. [Google Scholar] [CrossRef]
- Kim, J.; Jeon, S.G.; Kwak, M.J.; Park, S.J.; Hong, H.; Choi, S.B.; Lee, J.H.; Kim, S.W.; Kim, A.R.; Park, Y.K.; et al. Triglyceride-Catabolizing Lactiplantibacillus plantarum GBCC_F0227 Shows an Anti-Obesity Effect in a High-Fat-Diet-Induced C57BL/6 Mouse Obesity Model. Microorganisms 2024, 12, 1086. [Google Scholar] [CrossRef]
- Hong, C.P.; Yun, C.H.; Lee, G.W.; Park, A.; Kim, Y.M.; Jang, M.H. TLR9 regulates adipose tissue inflammation and obesity-related metabolic disorders. Obesity 2015, 23, 2199–2206. [Google Scholar] [CrossRef]
- Drucker, D.J. The benefits of GLP-1 drugs beyond obesity. Science 2024, 385, 258–260. [Google Scholar] [CrossRef]
- Pratley, R.E.; Kang, J.; Trautmann, M.E.; Hompesch, M.; Han, O.; Stewart, J.; Sorli, C.H.; Jacob, S.; Yoon, K.H. Body weight management and safety with efpeglenatide in adults without diabetes: A phase II randomized study. Diabetes Obes. Metab. 2019, 21, 2429–2439. [Google Scholar] [CrossRef]
- Wharton, S.; Blevins, T.; Connery, L.; Rosenstock, J.; Raha, S.; Liu, R.; Ma, X.; Mather, K.J.; Haupt, A.; Robins, D.; et al. Daily Oral GLP-1 Receptor Agonist Orforglipron for Adults with Obesity. N. Engl. J. Med. 2023, 389, 877–888. [Google Scholar] [CrossRef]
- Saxena, A.R.; Frias, J.P.; Brown, L.S.; Gorman, D.N.; Vasas, S.; Tsamandouras, N.; Birnbaum, M.J. Efficacy and Safety of Oral Small Molecule Glucagon-Like Peptide 1 Receptor Agonist Danuglipron for Glycemic Control Among Patients with Type 2 Diabetes: A Randomized Clinical Trial. JAMA Netw. Open 2023, 6, e2314493. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene | Forward Sequence (5′ to 3′) | Reverse Sequence (3′ to 5′) |
|---|---|---|
| hGAPDH | GGAGCGAGATCCCTCCAAAAT | GGCTGTTGTCATACTTCTCATGG |
| hGCG | ACCAGAAGACAGCAGAAATG | GAATGTGCCCTGTGAATG |
| hPCSK1 | CAGAAGGCTTTTGAATATGGTGT | GGAGGCACTGCTGATGGAGAT |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kim, A.-R.; Jeon, S.-G.; Park, S.-J.; Hong, H.; Kim, B.K.; Kim, H.-R.; Hong, C.-P.; Yang, B.-G. Alleviation of Adipose Tissue Inflammation and Obesity Suppression by a Probiotic Strain That Induces GLP-1 Secretion. Microorganisms 2025, 13, 1211. https://doi.org/10.3390/microorganisms13061211
Kim A-R, Jeon S-G, Park S-J, Hong H, Kim BK, Kim H-R, Hong C-P, Yang B-G. Alleviation of Adipose Tissue Inflammation and Obesity Suppression by a Probiotic Strain That Induces GLP-1 Secretion. Microorganisms. 2025; 13(6):1211. https://doi.org/10.3390/microorganisms13061211
Chicago/Turabian StyleKim, A-Ram, Seong-Gak Jeon, So-Jung Park, Heeji Hong, Byung Kwon Kim, Hyung-Ran Kim, Chun-Pyo Hong, and Bo-Gie Yang. 2025. "Alleviation of Adipose Tissue Inflammation and Obesity Suppression by a Probiotic Strain That Induces GLP-1 Secretion" Microorganisms 13, no. 6: 1211. https://doi.org/10.3390/microorganisms13061211
APA StyleKim, A.-R., Jeon, S.-G., Park, S.-J., Hong, H., Kim, B. K., Kim, H.-R., Hong, C.-P., & Yang, B.-G. (2025). Alleviation of Adipose Tissue Inflammation and Obesity Suppression by a Probiotic Strain That Induces GLP-1 Secretion. Microorganisms, 13(6), 1211. https://doi.org/10.3390/microorganisms13061211