Abstract
Ready-to-eat (RTE) foods can carry antimicrobial-resistant pathogens; however, few studies link real-world surveillance to practical interventions. This study addressed this gap by estimating the prevalence of Staphylococcus aureus (S. aureus) and methicillin-resistant S. aureus (MRSA) in ready-to-eat foods from Al-Qassim and evaluating a rapid, orthogonal confirmation workflow (culture → MALDI-TOF MS → Vitek 2 → mecA/mecC PCR). The in vitro activity of citrate-stabilized silver nanoparticles (AgNPs) against food-derived MRSA was quantified, and synergy with oxacillin (primary) and ciprofloxacin (secondary) was examined. Silver-susceptibility stability was assessed over 20 days of sub-MIC serial passage, with attention to whether β-lactam co-exposure constrained drift. We surveyed 149 RTE products and paired the confirmation workflow with mechanistic tests of AgNPs as antibiotic adjuvants. S. aureus was recovered from 24.2% of products and MRSA from 6.7%, with higher recovery from animal-source matrices and street-vendor outlets. MALDI-TOF MS provided rapid species confirmation and revealed two reproducible low-mass peaks (m/z 3990 and 4125) associated with MRSA, supporting spectral triage pending molecular confirmation. Antimicrobial susceptibility testing showed the expected β-lactam split (MRSA oxacillin/cefoxitin non-susceptible; MSSA oxacillin-susceptible but largely penicillin-resistant), with last-line agents retained. Citrate-stabilized AgNPs displayed consistent potency against food-derived MRSA (MIC 8–32 µg/mL; MIC50 16; MIC90 32) and were predominantly bactericidal (MBC/MIC ≤ 4 in 90%). Checkerboards demonstrated frequent AgNP–oxacillin synergy (median fractional inhibitory concentration index [FICI] 0.37; 4–16-fold oxacillin MIC reductions) and additive-to-synergistic effects with ciprofloxacin (median FICI 0.63), translating time–kill assays into rapid, sustained bactericidal activity without antagonism. During sub-MIC evolution, silver MICs rose modestly (median two-fold) and often regressed off drug; oxacillin co-exposure limited drift. RTE foods therefore represent credible MRSA exposure routes. Integrating MALDI-assisted triage with automated AST enables scalable surveillance, and standardized AgNP formulations emerge as promising β-lactam adjuvants—pending in situ efficacy, safety, and residue evaluation.
1. Introduction
Methicillin-resistant Staphylococcus aureus (MRSA) remains a high-priority pathogen at the human–food–environment interface [1,2]. Beyond its clinical burden, converging evidence shows that Staphylococcus aureus (S. aureus) including MRSA contaminates diverse ready-to-eat (RTE) foods, positioning food as a plausible vehicle for exposure and dissemination across One Health sectors [3,4]. Systematic reviews and meta-analyses report substantial S. aureus contamination in RTE categories and meats, with non-trivial pooled MRSA prevalence in animal-derived foods [5]. Complementary multi-country and regional assessments likewise document global MRSA contamination in meat products, broader S. aureus occurrence across food matrices including RTE items, and European surveillance confirming MRSA detection within harmonized monitoring frameworks [6,7,8].
Risk interpretation is nuanced. Authoritative reviews note that direct evidence for human infection via consumption of MRSA-contaminated foods remains limited; nonetheless, food can serve as a contamination source and reservoir for livestock-associated lineages. Genomic and epidemiologic investigations document transmission at animal–human interfaces (occupational and household) and suggest occasional foodborne routes [9,10].
Global prioritization initiatives consistently rank S. aureus—including resistant forms—among major public health concerns, reflecting high burden, transmission potential, and constrained therapeutic options. Large-scale burden analyses place S. aureus, especially MRSA, among leading contributors to deaths attributable to antimicrobial resistance (AMR), with regional assessments corroborating substantial MRSA-attributable deaths and disability-adjusted life years (DALYs) [11,12,13].
Robust identification of foodborne MRSA benefits from complementary platforms that combine speed, specificity, and confirmatory power. Matrix-assisted laser desorption/ionization time-of-flight (MALDI-TOF) mass spectrometry enables rapid species-level identification and, in exploratory workflows, has reported spectral patterns associated with MRSA versus methicillin-susceptible S. aureus (MSSA); however, performance varies by protocol and dataset, and reliable resistance determination still requires molecular confirmation [14,15]. Automated antimicrobial susceptibility testing (AST) systems such as Vitek 2 provide high categorical/essential agreement for Staphylococcus susceptibility testing and can support early MRSA flagging at scale [16,17].
Definitive confirmation relies on PCR targeting mecA/mecC (and, where appropriate, SCCmec–orfX junction assays) and may be complemented by penicillin-binding protein 2a (PBP2a) antigen detection in selected workflows; these remain contemporary reference standards in MRSA diagnostics [18]. Best practice screening uses cefoxitin as a surrogate for PBP2a-mediated resistance with harmonized CLSI/EUCAST breakpoints and quality control; nevertheless, mecC-associated detection gaps and occasional phenotype–genotype discrepancies warrant confirmatory mecA/mecC testing when discordance or epidemiologic significance is suspected [19,20].
Against the backdrop of escalating AMR, silver nanoparticles (AgNPs) exhibit a multitarget antibacterial mode of action in S. aureus—including membrane disruption, metabolic/proteostasis collapse, nucleic-acid interactions, and reactive-oxygen-species effects—and can potentiate conventional antibiotics. Checkerboard and time–kill studies report AgNP–antibiotic synergy with biofilm suppression, particularly with β-lactams, supporting their evaluation as adjuvants to restore or enhance drug efficacy [21,22,23]. Synergistic strategies that pair AgNPs with antibiotics—and, in some cases, adjunct modalities such as visible blue light or plant-derived polyphenols—produce augmented bactericidal activity against MRSA and biofilm-forming S. aureus, reinforcing the translational rationale for AgNP-enabled combination therapy [21,24,25].
Durability of AgNP efficacy, however, cannot be assumed. Experimental evolution and serial passage studies show that S. aureus can increase minimum inhibitory concentrations (MICs) under repeated sub-inhibitory exposure—potentially via nanoparticle aggregation driven by enhanced biofilm formation and secreted factors—while covalent immobilization of silver on cyanographene has restored activity against AgNP-resistant phenotypes [26,27]. Conversely, other work reported no stable resistance after 16 serial passages with Ag+ or AgNPs, highlighting context-dependent outcomes and the need for long-term tests in food-derived MRSA under clinically relevant AgNP ± antibiotic regimens [21]. To date, few studies have simultaneously tracked long-term AgNP susceptibility in food-derived MRSA while quantifying AgNP–antibiotic interactions under standardized conditions.
From a One Health perspective, heavy metals and other non-antibiotic stressors may co-select for antibiotic resistance, shaping antimicrobial-resistance-gene persistence in food-processing environments and along supply chains. Understanding how silver-based interventions intersect with such co-selection dynamics is essential for risk-aware deployment in food safety and clinical contexts [28].
Accordingly, the present study estimated the prevalence of S. aureus/MRSA in RTE foods from Al-Qassim; evaluated a rapid, orthogonal confirmation workflow (culture → MALDI-TOF MS → Vitek 2 → mecA/mecC PCR); quantified the in vitro activity of citrate-stabilized AgNPs and their interactions with oxacillin and ciprofloxacin against food-derived MRSA; and assessed the stability of silver susceptibility during 20 days of sub-MIC serial passage, including whether β-lactam co-exposure constrains drift. By integrating surveillance with standardized confirmation, synergy testing, and sub-MIC evolution assays, this work provides translational evidence at the food–human interface to inform One Health-aligned AMR-mitigation strategies in the RTE sector.
2. Materials and Methods
2.1. Study Design and Sampling
This cross-sectional survey was carried out in the Al-Qassim region of Saudi Arabia from December 2024 to April 2025. A total of 149 RTE items were collected using a cross-stratified approach by food category—meat-based (n = 40), poultry-based (n = 32), salads/vegetables (n = 28), dairy-based (n = 25), and bakery/sweets (n = 24)—and by point of sale—restaurants/takeaways (n = 60), supermarkets/delis (n = 54), and street vendors/markets (n = 35) (Table 1). Purchases were distributed across multiple municipalities and staggered across different days and districts to reduce clustering and improve representativeness. All items were transported in insulated coolers maintained at ≤ 8°C and were processed within 4 h of purchase.

Table 1.
Allocation of 149 RTE food samples across categories and outlet types, Al-Qassim. Sampling period: December 2024–April 2025.
2.2. Culture and Preliminary MRSA Screening
Food homogenates were pre-enriched in Tryptic Soy Broth (TSB; Oxoid, Thermo Fisher Scientific, Altrincham, UK) supplemented with 6.5% NaCl (Sigma-Aldrich/Merck, Darmstadt, Germany) at a 1:10 ratio (25 g in 225 mL) and incubated at 37 °C for 18–24 h. Enrichments were streaked onto Baird–Parker agar with egg yolk tellurite emulsion (Oxoid, Thermo Fisher Scientific, UK) and CHROMagar™ MRSA (CHROMagar, Paris, France), followed by incubation at 37 °C for 24–48 h. Up to three colonies per sample with morphology compatible with Staphylococcus aureus were purified on Tryptic Soy Agar (TSA; Oxoid, Thermo Fisher Scientific, UK) and evaluated by Gram stain (Remel Gram Stain Kit, Thermo Fisher Scientific), catalase (3% H2O2; Sigma-Aldrich/Merck), and coagulase (Oxoid Staphytect™ Plus latex agglutination, Thermo Fisher Scientific). DNase agar (Oxoid, Thermo Fisher Scientific, UK) was used when required by SOPs. Isolates meeting these criteria were classified as presumptive S. aureus and advanced to methicillin-resistance screening.
Preliminary MRSA screening used a cefoxitin-surrogate approach aligned with international standards. From overnight culture, a 0.5 McFarland suspension (0.85% sterile saline; Oxoid, Thermo Fisher Scientific, UK) was lawn-inoculated on Mueller–Hinton agar (MHA; Oxoid, Thermo Fisher Scientific, UK), a 30 µg cefoxitin disk (Oxoid antimicrobial susceptibility disk, Thermo Fisher Scientific) was applied, and plates were incubated at 35 ± 2 °C for 18–24 h. Zone diameters were interpreted according to contemporaneous CLSI M100 performance standards and CLSI M07 guidance, together with EUCAST breakpoints (cefoxitin as a surrogate for PBP2a-mediated resistance) [29,30]. Isolates with borderline zones, mixed phenotypes, or epidemiologically important profiles were retested and flagged for orthogonal confirmation with PBP2a antigen testing (Oxoid™ PBP2′ Latex Agglutination Test, DR0900A, Thermo Fisher Scientific) and/or mecA/mecC PCR (see Molecular Methods).
Quality control was included in every run using S. aureus ATCC 25923 (disk-diffusion QC), ATCC 29213 (MIC reference QC), and ATCC 43300 (MRSA, mecA-positive) (all from ATCC, Manassas, VA, USA). Media and disks were used within shelf life, stored per manufacturer specifications, and inspected for plate integrity before use.
2.3. Species Identification by MALDI-TOF MS
Species identification was performed by MALDI-TOF MS (MALDI Biotyper, Bruker Daltonics, Bremen, Germany) using MBT Compass software (v1.3) and the IVD reference library. Targets were calibrated before each run with the Bruker Bacterial Test Standard (BTS; Escherichia coli) according to the manufacturer’s acceptance criteria. For routine preparation, a well-isolated colony was spotted onto a polished steel target (96-well format), overlaid with 1 µL of 70% formic acid, air-dried, and overlaid with 1 µL of HCCA matrix (α-cyano-4-hydroxycinnamic acid; Bruker). Spectra were acquired in linear positive mode over 2–20 kDa using manufacturer-recommended parameters. Isolates yielding weak or ambiguous spectra were reprocessed by the ethanol–formic acid extraction protocol and reanalyzed.
The MALDI workflow served primarily to confirm S. aureus at the species level and secondarily, on an exploratory basis, to flag presumptive MRSA versus MSSA using a laboratory-validated rule set based on low-mass peak-intensity signatures across ≥2 independent spots. Indeterminate spectra were not called. Methicillin resistance was not assigned by MALDI; all isolates underwent cefoxitin-based phenotypic screening interpreted per contemporaneous CLSI M100/M07 and EUCAST criteria, and any resistant, discordant, or MALDI-flagged isolates were resolved by PBP2a antigen testing and/or mecA/mecC PCR (details in Molecular Methods).
2.4. Antimicrobial Susceptibility Testing (AST)
Phenotypic AST was performed on the VITEK® 2 Compact (bioMérieux, Marcy-l’Étoile, France) using the Staphylococcus card AST-GP71. The panel included cefoxitin (screen), oxacillin, penicillin, erythromycin, clindamycin, gentamicin, ciprofloxacin, levofloxacin, tetracycline, trimethoprim–sulfamethoxazole, linezolid, and vancomycin. Inocula were prepared from 18 to 24 h cultures, adjusted to a 0.5 McFarland standard in sterile saline, and loaded according to the manufacturer’s instructions. Results were interpreted against contemporaneous CLSI M100 and EUCAST breakpoints.
Quality control accompanied each new lot and whenever control criteria indicated repeat testing, using S. aureus ATCC 29213 and ATCC 43300. For key analytes—oxacillin/cefoxitin and ciprofloxacin—reference broth microdilution (BMD; CLSI M07/M100) was performed for all isolates, and vancomycin MICs were likewise confirmed by BMD (see Section 2.5).
2.5. Broth Microdilution MIC and MBC Testing
Reference MICs for antibiotics and silver nanoparticles (AgNPs) were determined by BMD in cation-adjusted Mueller–Hinton broth (CAMHB; Oxoid, Thermo Fisher Scientific, UK) using sterile, low-binding 96-well plates (final well volume 100 µL). Twofold serial dilutions were prepared at 2× strength in 50 µL and mixed 1:1 with 50 µL of inoculum to yield a final concentration of 5 × 105 CFU/mL. Plates were incubated at 35 ± 2 °C for 18–24 h and read visually. The MIC was defined as the lowest concentration with no visible growth. For minimum bactericidal concentration (MBC), 10 µL from non-turbid wells at or above the MIC were plated onto TSA and incubated for 24 h; the MBC was the lowest concentration producing ≥99.9% of kills relative to the starting inoculum. Unless otherwise noted, categorical and essential agreement statements refer to BMD MICs.
Antibiotic test ranges (twofold series; µg/mL): oxacillin (CAMHB + 2% NaCl; read at 24 h) 0.25–8; cefoxitin 0.5–32; vancomycin 0.125–16; linezolid 0.125–8; ciprofloxacin 0.004–2; gentamicin 0.125–32; erythromycin 0.125–64; clindamycin 0.03–16; tetracycline 0.125–64; and trimethoprim–sulfamethoxazole (1:19; reported as trimethoprim) 0.06/1.2–4/76. AgNPs: 0.5–128 µg/mL total silver (twofold series), prepared from quantified stocks and gently resuspended immediately before dispensing.
AgNP-specific precautions: CAMHB was prepared with low-chloride water; plates were protected from light; and edge wells were avoided to limit evaporation and nanoparticle ring effects. Where a solvent/dispersant was required for antibiotic stocks, the final concentration did not exceed 1% v/v and was matched in control wells. Controls included medium sterility and organism growth controls for each isolate. Visualization (checkerboard assays): Heatmaps are displayed as percent growth at 24 h with numeric color bar ticks (0, 25, 50, 75, 100%) and single-agent MICs are annotated on panel margins for reference.
2.6. Molecular Confirmation of MRSA
Genomic DNA was prepared by boiling lysis; a commercial kit was used for isolates with low yields. A single-tube multiplex PCR targeted mecA, mecC (also referred to as mecAL-GA251), spa, and—optionally—lukF-PV (PVL), using a validated scheme that distinguishes mecA from mecC while enabling simultaneous detection of spa and PVL [31]. Primer sets and expected amplicon sizes followed the EURL-AR MRSA Multiplex PCR-2 protocol [32]: mecA P4/P7 (162 bp), mecC MultiFP/MultiRP (138 bp), spa 1113F/1514R (~180–600 bp, variable), and lukF-PV pvl-F/pvl-R (~85 bp). The spa primers (1113F/1514R) derive from the standard spa-typing approach, and the PVL primers correspond to the original PVL detection method in S. aureus [33].
Thermocycling conditions were: 95 °C for 5 min; 35 cycles of 95 °C for 30 s, 55 °C for 30 s, and 72 °C for 45 s; followed by 72 °C for 5 min. Amplicons were resolved on 2% agarose and visualized with ethidium bromide (or a nonhazardous equivalent). Each run included S. aureus ATCC 43300 (mecA-positive), a confirmed mecC-positive external control, and a no-template control. Isolates were designated MRSA when mecA or mecC was detected at the expected size and the phenotype supported methicillin resistance. Any discordant result prompted repeat extraction and PCR, and—when indicated—PBP2a antigen testing as an orthogonal confirmatory method.
2.7. AgNPs: Source, Preparation, and Characterization
Citrate-stabilized, spherical AgNPs with a nominal core diameter of 10 nm (Sigma-Aldrich/Merck, cat. 730785) were used throughout. Stock dispersions (1–2 mg/mL) were stored at 4 °C, protected from light, gently resuspended immediately before use, and discarded after 30 days. Each lot was characterized before biological testing. Ultraviolet–visible (UV–Vis) spectroscopy verified the expected surface plasmon resonance for small spherical AgNPs (typically ~400–430 nm). Dynamic light scattering (DLS) provided hydrodynamic diameter and polydispersity index (PDI); zeta potential (via electrokinetic mobility) was measured to assess colloidal stability. For a subset, transmission electron microscopy (TEM) confirmed core size and morphology. Total silver in working stocks was quantified by inductively coupled plasma mass spectrometry (ICP-MS).
To minimize nanoparticle loss and artifact formation, all dilutions and measurements used low-protein-binding labware with handling aligned to dispersion-stability best practices. Methods followed standards from the International Organization for Standardization (ISO)—ISO 22412 (DLS) [34], ISO 13099-1 (zeta potential) [35], ISO 17294-2 (ICP-MS) [36]—and stability considerations were guided by the Organization for Economic Co-operation and Development (OECD) Test Guideline 318. UV–Vis peak positions and line shapes were consistent with published behavior for citrate-stabilized AgNPs of comparable size [37].
2.8. AgNP–Antibiotic Combination Testing: Checkerboard Assay
AgNP–antibiotic interactions were evaluated using a two-dimensional checkerboard microdilution assay in CAMHB on low-binding 96-well plates (final well volume 100 µL). Along the horizontal axis, AgNPs (reported as µg/mL total Ag) were dispensed in a twofold series, typically 0.5–128 µg/mL. Along the vertical axis, the partner antibiotic—oxacillin (primary) or ciprofloxacin (secondary)—was dispensed in a twofold series spanning its working range (see Section 2.5).
Each well received 50 µL of the 2× agent mixture and 50 µL of inoculum adjusted to 5 × 105 CFU/mL. Plates were incubated at 35 ± 2 °C for 18–24 h and read visually to avoid optical interference from AgNPs. For every isolate, the MIC of each agent alone (single-agent rows/columns) and in combination (matrix wells) was recorded. To preserve dispersion stability and accuracy, plates were protected from light and edge wells were avoided to limit evaporation and nanoparticle ring effects.
Controls included sterility and growth controls, vehicle/dispersant controls, AgNO3 (ionic silver) at total-silver levels matched to AgNPs, and single-agent controls at the highest in-plate concentrations. All experiments were performed in biological duplicates with technical triplicates per condition.
Interaction was expressed as the FICI, calculated as:
By convention, an FICI ≤ 0.5 indicates synergy, >0.5–1.0 additivity, >1.0–4.0 indifference, and >4.0 antagonism. When an MIC was off-scale (above the highest concentration tested), the top concentration was used to provide a conservative FICI estimate. FICI values were summarized as the median across technical replicates and the mean across biological duplicates. Representative combinations categorized as synergistic in the checkerboard were subsequently validated by time–kill assays (Section 2.9).
2.9. Time–Kill Kinetics
Time–kill assays were performed on a subset of representative MRSA isolates. Cultures (starting ~106 CFU/mL) were exposed to AgNPs and to antibiotics—alone and in combination—at 0.5 × −1 × the MIC for each agent. Aliquots were collected at 0, 2, 4, 8, and 24 h, serially diluted, and plated for enumeration (CFU/mL). Synergy was defined as a ≥2 log10 CFU/mL reduction with the combination compared with the most active single agent at 24 h. Bactericidal activity was defined as a ≥3 log10 CFU/mL reduction from baseline.
2.10. Biofilm Assays (Subset Analysis)
For a subset of MRSA isolates (≥10), biofilm formation was quantified in flat-bottom 96-well plates under static conditions (24 h, 37 °C). Following treatment with AgNPs with or without antibiotics, biofilms were assessed by crystal violet staining (OD590) and by viable counts from biofilm resuspensions to estimate the minimum biofilm eradication concentration (MBEC). All conditions were tested in triplicate.
2.11. Resistance Evolution Experiment (Serial Passage)
To examine the potential for resistance development, selected MRSA isolates (≥10, spanning food categories and outlet types) were serially passaged once daily for 20–30 passages in CAMHB containing 0.5 × MIC AgNPs. Parallel arms included AgNPs plus sub-MIC oxacillin and oxacillin alone. Every five passages, MICs for AgNPs and the partner antibiotics were re-determined by broth microdilution. Frozen archives (−80 °C) were prepared at each interval. When AgNP MICs increased, we additionally measured growth rates, collected UV–Vis spectra from the spent medium to detect AgNP aggregation or surface-plasmon resonance (SPR) shifts, and assessed zeta potential. Selected ancestor–evolved pairs were retained for targeted sequencing of candidate loci. Ancestral and endpoint stocks (with intermediates, where indicated) were archived at −80 °C in glycerol for planned whole-genome sequencing.
2.12. Data Management and Statistical Analysis
Prevalence estimates were summarized as % (95% CI) by food category and outlet type; comparisons used χ2 or Fisher’s exact tests, as appropriate. MIC data were reported as MIC50/MIC90 and geometric means. Checkerboard results were summarized as median (IQR) FICI and categorized as synergy/additive/indifferent/antagonism using standard thresholds. Time–kill data were analyzed from log10 CFU/mL trajectories, with synergy and bactericidal activity adjudicated at 24 h as defined above.
Trends in resistance evolution were modeled using linear mixed-effects regression of log2(MIC) versus passage, with isolate as a random intercept; the Benjamini–Hochberg procedure controlled the false discovery rate. With n = 149, two-sided exact binomial 95% confidence intervals yield approximate half-widths of ±3.5, ±4.8, ±6.4, and ±7.4 percentage points at true prevalences of 5%, 10%, 20%, and 30%, respectively, indicating ~3–8 percentage-point precision across the typical range. Analyses were performed in R (v4.3+) and GraphPad Prism (v10) with α = 0.05.
2.13. Biosafety and Quality Assurance
All work was conducted under Biosafety Level 2 conditions with appropriate chemical safety for silver handling and waste disposal. ATCC controls were included in each run, and new media/consumable lots were verified against controls before study use. Instruments underwent routine calibration and internal quality control; any deviations triggered repeat testing or exclusion according to SOPs.
3. Results
3.1. Sample Characteristics and Recovery of S. aureus
We analyzed 149 RTE items collected between December 2024 and April 2025 using the cross-stratified sampling frame described in Table 1. After enrichment and selective/differential plating, presumptive S. aureus was recovered from 36 of the 149 samples (24.2%). Recovery differed by food category: meat-based 11/40 (27.5%), poultry-based 9/32 (28.1%), salads/vegetables 6/28 (21.4%), dairy-based 7/25 (28.0%), and bakery/sweets 3/24 (12.5%). Recovery also varied by outlet type: restaurants/takeaways 13/60 (21.7%), supermarkets/delis 11/54 (20.4%), and street vendors/markets 12/35 (34.3%). Exact proportions with two-sided 95% confidence intervals are reported in Table 2. To illustrate precision, exact binomial 95% confidence intervals are shown alongside all proportions. For a total sample size of 149, half-widths are approximately 5% of points when prevalence is near 10%, with narrower or wider intervals at lower or higher prevalences, respectively.
Table 2.
Recovery of presumptive S. aureus from RTE foods by category and outlet type.
3.2. Confirmation of S. aureus, MRSA Prevalence, and MALDI Peak Signatures
Of the 36 presumptive isolates, 34 (94.4%) were confirmed as S. aureus by MALDI-TOF MS at species-level confidence (31 by direct smear; 3 after ethanol–formic acid extraction). Two isolates were reclassified as non-aureus staphylococci. Cefoxitin screening flagged 11 putative MRSA. Multiplex PCR detected mecA in 10 isolates and mecC in 0, yielding 10 MRSA and 24 MSSA overall. One cefoxitin-non-susceptible isolate was negative for mecA/mecC and PBP2a on repeat testing and was classified as MSSA. The corresponding MRSA prevalence among RTE items was 10/149 (6.7%; 95% CI, 3.3–11.8%). By food category, MRSA was detected in meat-based 4/40 (10.0%), poultry-based 3/32 (9.4%), salads/vegetables 1/28 (3.6%), dairy-based 1/25 (4.0%), and bakery/sweets 1/24 (4.2%). By outlet type, prevalence was 4/60 (6.7%) in restaurants/takeaways, 3/54 (5.6%) in supermarkets/delis, and 3/35 (8.6%) among street vendors/markets (Table 3).
Table 3.
Prevalence of MRSA among RTE food samples by category and outlet type.
In parallel, MALDI overlays revealed two reproducible low-mass peaks at m/z 3990 and 4125 that were present in all MRSA isolates (10/10, 100%) and absent in all MSSA isolates (0/24, 0%) (Figure 1). These signals were used strictly for preliminary triage. Final MRSA designations relied on cefoxitin-based screening with mecA/mecC confirmation, as detailed in the Methods.
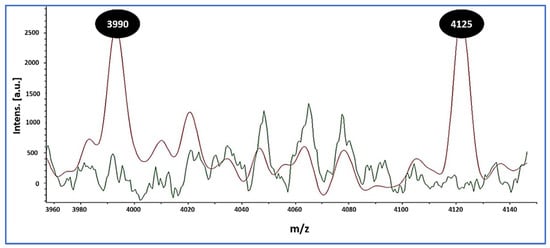
Figure 1.
MALDI-TOF MS overlay distinguishing MRSA from MSSA. MRSA spectra (red) show reproducible peaks at mass-to-charge ratio (m/z) 3990 and 4125, whereas these peaks are absent in MSSA spectra (green). Inset: per-isolate spectra/spot counts (median [IQR]) and the proportion of MRSA exhibiting both peaks across ≥2 independent spots (10/10, 100%), with 0/24 (0%) MSSA showing either peak. These low-mass signatures differentiate MRSA from MSSA in this dataset and were used for spectral triage; definitive MRSA calls relied on a cefoxitin screen with mecA/mecC PCR.
3.3. Antimicrobial Susceptibility Profiles and Agreement Metrics (Vitek 2 AST-GP71)
Using the Vitek 2 Compact AST-GP71 panel, 34 Staphylococcus aureus isolates were profiled (MRSA, n = 10; MSSA, n = 24). Unless noted, interpretive categories and agreement metrics are referenced to BMD for oxacillin/cefoxitin, ciprofloxacin, and vancomycin (Table 4). As can be seen from Table 4, the β-lactam split was stark: oxacillin and benzylpenicillin (penicillin G) were 0/10 (0.0%) susceptible in MRSA, whereas oxacillin was 24/24 (100%) susceptible in MSSA and benzylpenicillin showed 2/24 (8.3%) susceptible and 22/24 (91.7%) resistant. Among fluoroquinolones, ciprofloxacin susceptibility was 1/10 (10.0%) in MRSA versus 22/24 (91.7%) in MSSA. For macrolides, erythromycin was 3/10 (30.0%) susceptible in MRSA and 19/24 (79.2%) in MSSA. Clindamycin (lincosamide) remained broadly active—7/10 (70.0%) MRSA and 22/24 (91.7%) MSSA susceptible. Gentamicin activity was high in both groups—9/10 (90.0%) MRSA and 23/24 (95.8%) MSSA susceptible. Rifampin susceptibility was 8/10 (80.0%) in MRSA and 22/24 (91.7%) in MSSA.
Table 4.
Antimicrobial susceptibility of S. aureus from RTE foods (MRSA n = 10; MSSA n = 24) and agreement metrics for Vitek 2 AST-GP71 versus reference methods.
Several non-β-lactam agents retained uniform or near-uniform in vitro activity across the collection: linezolid, daptomycin, tigecycline, nitrofurantoin, quinupristin–dalfopristin, and vancomycin were 34/34 (100%) susceptible. Trimethoprim–sulfamethoxazole (TMP–SMX) was 33/34 (97.1%) susceptible overall, driven by a single resistant MSSA isolate (1/24, 4.2%). Agreement metrics were excellent. Categorical agreement (CA) reached 100.0% for most drugs (ciprofloxacin, daptomycin, linezolid, nitrofurantoin, oxacillin, quinupristin–dalfopristin, rifampin, tigecycline, TMP–SMX, vancomycin). CA was 97.1% for clindamycin and gentamicin (each with one minor error [mE] = 2.9%) and 97.1% for the cefoxitin screen (one major error [ME] = 4.2% among reference-susceptible MSSA). Benzylpenicillin showed 94.1% CA with two minor errors (5.9%). Very major errors (VME) were 0% for all agents. Essential agreement (EA) was not assessed for the cefoxitin screen and was not calculated for the remaining agents in this dataset.
These results confirm a clear MRSA–MSSA divide for β-lactams and ciprofloxacin, consistently high activity of multiple non-β-lactam agents against food-derived isolates, and robust categorical performance of the Vitek 2 AST-GP71 panel, with no VMEs and only isolated MEs/mEs.
3.4. In Vitro Activity of AgNPs: MICs and MBCs
Citrate-stabilized AgNPs demonstrated consistent inhibitory activity against all MRSA isolates (n = 10). MICs clustered within a narrow range of 8–32 µg/mL, with an MIC50 of 16 µg/mL and an MIC90 of 32 µg/mL (Table 5). Corresponding MBCs ranged from 16 to 128 µg/mL (median 32 µg/mL). In nine of ten isolates, the MBC/MIC ratio was ≤4, indicating bactericidal action at concentrations close to the MIC. One isolate required an MBC eightfold higher than its MIC (MBC/MIC = 8), consistent with a non-bactericidal effect under the test conditions.
Table 5.
In vitro activity of AgNPs against MRSA: MIC, MBC, killing classification, fold-change vs. MIC50, and baseline oxacillin/ciprofloxacin MICs (broth microdilution).
Replicate determinations were highly reproducible, agreeing within ±1 twofold dilution, and all assay controls performed within acceptable limits. No clear association between MIC distribution and food category or outlet type was observed, likely due to the modest sample size. In summary, across the MRSA collection (n = 10), AgNPs displayed MICs ranging from 8 to 32 µg/mL (MIC50 = 16 µg/mL; MIC90 = 32 µg/mL). Corresponding MBCs spanned 16–128 µg/mL, yielding a median MBC/MIC ratio of two. Using the conventional bactericidal criterion (MBC/MIC ≤ 4), nine of ten isolates (90.0%) exhibited bactericidal activity. The MIC was defined as the lowest concentration preventing visible growth after 18–24 h, and the MBC as the lowest concentration achieving ≥99.9% reduction in viable counts. These results confirm that the tested AgNP formulation exerted strong and predominantly bactericidal effects against food-derived MRSA isolates.
3.5. AgNP–Antibiotic Interactions in MRSA (Checkerboard FICI)
Checkerboard assays showed consistent potentiation of oxacillin by AgNPs against food-derived MRSA. Across ten isolates, the AgNP–oxacillin combination yielded synergy in 6/10 and additivity in 4/10, with no indifference or antagonism observed. The FICI had a median of 0.37 (IQR, 0.28–0.50), accompanied by 4–16-fold reductions in oxacillin MIC and 2–8-fold reductions in AgNP MIC compared with single-agent exposures. For AgNP–ciprofloxacin, synergy occurred in 3/10 isolates, additivity in 5/10, and indifference in 2/10 (no antagonism). The median FICI was 0.63 (IQR, 0.50–0.88), indicating an overall additive-to-synergistic effect. All interaction calls were reproducible across biological duplicates, with ≤1 twofold dilution difference between runs. Taken together, these data indicate that AgNPs re-sensitize a substantial fraction of MRSA to a β-lactam backbone and augment fluoroquinolone activity without detectable antagonism under the conditions tested (Figure 2; Supplementary Table S1).
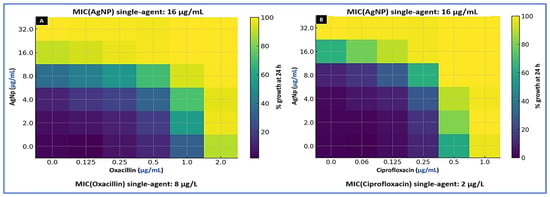
Figure 2.
Checkerboard interaction heatmaps for citrate-stabilized silver nanoparticles (AgNPs) combined with (A) oxacillin and (B) ciprofloxacin against representative MRSA isolates. Axes show two-fold dilution series of AgNP (y-axis, µg/mL) and antibiotic (x-axis, µg/mL). Heatmap values represent percent growth at 24 h (visual read); the color bar uses numeric ticks (0, 25, 50, 75, 100%). Single-agent MICs for the isolate depicted are MIC (AgNP) = 16 µg/mL, MIC (oxacillin) = 8 µg/mL, and MIC (ciprofloxacin) = 2 µg/mL. FICI interpretation: synergy (≤0.5), additivity (>0.5–1.0), indifference (>1.0–4.0), and antagonism (>4.0). No antagonism was observed.
3.6. Time–Kill Kinetics for AgNP Combinations
Time–kill assays were conducted on four MRSA isolates—two that showed synergy and two that showed additivity in checkerboard testing—to validate the dynamic effects of AgNP–antibiotic combinations using fixed concentrations rather than MIC multiples. For isolates with AgNP MICs of 8, 16, and 32 µg/mL, AgNPs were tested at 4 and 8 µg/mL, 8 and 16 µg/mL, and 16 and 32 µg/mL, respectively. Oxacillin was tested at 4 and 8 µg/mL, while ciprofloxacin was tested at 0.25 and 0.5 µg/mL (MIC 0.5 µg/mL), 0.5 and 1 µg/mL (MIC 1 µg/mL), or 1 and 2 µg/mL (MIC 2 µg/mL), matched to each isolate’s baseline susceptibility profile.
When AgNPs were combined with oxacillin, the regimen produced rapid and sustained bactericidal activity. At the higher combination dose (AgNP 8–32 µg/mL plus oxacillin 8 µg/mL), two isolates dropped below the limit of detection (10 CFU/mL) by 24 h, and the remaining two achieved ≥3-log10 CFU/mL reductions without regrowth. At the lower combination dose (AgNP 4–16 µg/mL plus oxacillin 4 µg/mL), three isolates showed reductions of approximately 2.3–3.8 log10 CFU/mL (synergy), and one isolate showed a 1.8 log10 CFU/mL reduction (additivity). In contrast, oxacillin alone (8 µg/mL) resulted in bacterial stasis or early regrowth, while AgNPs alone (8–32 µg/mL) produced only modest decreases of 0.5–1.2 log10 CFU/mL after 24 h.
When AgNPs were combined with ciprofloxacin (AgNP 4–32 µg/mL plus ciprofloxacin 0.25–2 µg/mL, dose-matched by isolate), the interaction was consistently additive-to-synergistic. Two isolates showed 1.5–2.7 log10 CFU/mL reductions, while two showed 1.0–1.4 log10 CFU/mL reductions, with no evidence of antagonism at any time point. Across all isolates, the combinations demonstrated an earlier onset of killing (within 4–8 h) and prevented regrowth more effectively than either agent alone. Representative kinetic curves are shown in Figure 3, while full isolate-level results—including 24 h Δlog10 CFU/mL changes, synergy/bactericidal adjudications, and limit-of-detection status—are detailed in Supplementary Table S2. All findings were reproducible across independent biological replicates, with pairwise differences ≤0.3 log10 CFU/mL at 24 h.
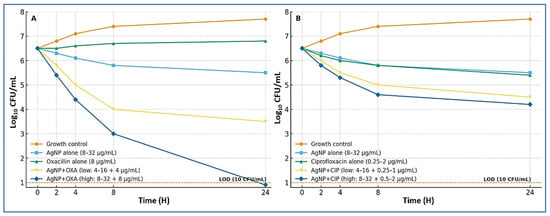
Figure 3.
Time–kill kinetics of AgNP combinations against MRSA. (A) AgNPs with oxacillin. Curves show the growth control, AgNPs alone (8–32 µg/mL), oxacillin alone (8 µg/mL), and two combination regimens: low dose (AgNPs 4–16 µg/mL plus oxacillin 4 µg/mL) and high dose (AgNPs 8–32 µg/mL plus oxacillin 8 µg/mL). The high-dose combination produced rapid, sustained bactericidal activity, with some isolates reaching the limit of detection (LOD; 10 CFU/mL, dashed line) by 24 h. (B) AgNPs with ciprofloxacin. Curves show the growth control, AgNPs alone (8–32 µg/mL), ciprofloxacin alone (0.25–2 µg/mL), and two combination regimens: low dose (AgNPs 4–16 µg/mL plus ciprofloxacin 0.25–1 µg/mL) and high dose (AgNPs 8–32 µg/mL plus ciprofloxacin 0.5–2 µg/mL). Combinations showed additive-to-synergistic killing without evidence of antagonism, with an earlier onset of killing (4–8 h) and reduced regrowth compared with single agents.
3.7. Experimental Evolution Under Sub-MIC AgNP Exposure
To model prolonged low-level pressure, isolates with baseline AgNP MICs of 8, 16, or 32 µg/mL were passaged once daily for 20 days in a CAMHB containing AgNPs at 4, 8, or 16 µg/mL, respectively. A parallel arm used the same AgNP levels plus oxacillin at 4 µg/mL. Both arms were followed by seven drug-free passages to assess stability of any changes. Under AgNP exposure alone, AgNP MICs increased modestly: the median change was twofold (IQR, 1–4-fold). Three isolates reached the pre-specified threshold for reduced susceptibility (≥4-fold rise), peaking at 4–8-fold above baseline. In two of these, the shift proved unstable and regressed to ≤2-fold after drug withdrawal; two isolates showed a durable increase and, together with their ancestors, were earmarked for targeted whole-genome sequencing in follow-up work.
Adding oxacillin constrained adaptation. In the AgNP-plus-oxacillin arm (AgNP 4–16 µg/mL with oxacillin 4 µg/mL), the median MIC change was 0-fold (IQR, 0–2-fold), and the single twofold elevation returned to baseline during drug-free passage. Across conditions, MBCs tended to rise in step with MICs, but the MBC/MIC ratio remained ≤4 for most endpoint cultures, indicating preserved bactericidal behavior despite modest shifts in susceptibility. No consistent cross-changes were detected in oxacillin or ciprofloxacin MICs. Overall, sustained sub-MIC AgNP exposure selected small, often reversible increases in AgNP MIC, whereas pairing AgNPs with a β-lactam limited adaptation under the conditions tested (Figure 4).
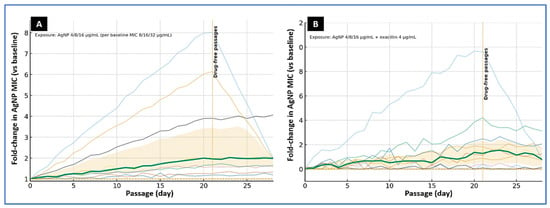
Figure 4.
Experimental evolution of foodborne MRSA under sub-MIC AgNP exposure. (A) AgNP alone. Ten isolates were serially passaged for 20 days in cation-adjusted Mueller–Hinton broth (CAMHB) containing AgNPs at 4, 8, or 16 µg/mL, matched to baseline AgNP MICs of 8, 16, or 32 µg/mL, respectively. (B) AgNP plus β-lactam. Parallel passages used AgNPs at 4, 8, or 16 µg/mL together with oxacillin at 4 µg/mL. Curves display individual isolate trajectories (thin lines) as fold-change in AgNP MIC relative to baseline; the bold line indicates the median, with the shaded band showing the interquartile range (IQR). The vertical dotted line at day 21 marks the start of drug-free passages. Under AgNP exposure alone, several isolates showed multi-fold MIC increases with partial regression during drug-free culture, whereas the AgNP–oxacillin combination constrained MIC drift.
4. Discussion
This study addresses a One Health priority by pairing real-world surveillance of RTE foods with rigorous confirmation of methicillin resistance and focused experiments on silver-based combination therapy. By integrating orthogonal identification methods—culture, MALDI-TOF MS, Vitek 2, and mecA/mecC PCR—with mechanistic assays (checkerboard synergy, time–kill kinetics, and experimental evolution), we move beyond prevalence reporting toward actionable intervention. We found that MRSA was present in 6.7% of RTE items, that citrate-stabilized AgNPs reliably restored or potentiated β-lactam activity with rapid, sustained bactericidal effects, and that prolonged sub-MIC exposure to silver produced only modest, often unstable MIC increases—further constrained when AgNPs were paired with oxacillin. These results support matrix-aware surveillance and combination-first strategies while cautioning against prolonged sub-therapeutic nanoparticle exposure.
From the 149 RTE items analyzed, S. aureus was recovered from 24.2% overall, with significantly higher recovery from animal-derived products and street vendors (≈1.7-fold higher than supermarkets/delis). This pattern mirrors global evidence implicating meat and dairy as high-risk matrices for staphylococcal contamination [5]. A recent meta-analysis estimated the global prevalence of MRSA in meats at ~3.7%, with the highest burden in the Eastern Mediterranean region [6]. Consistent findings have been reported in China, where Wang et al. [38] identified multidrug-resistant MRSA in retail foods and Wu et al. [3] documented notable contamination across diverse categories. In Europe, Basanisi et al. [39] reported 2.4% MRSA prevalence in southern Italian retail meats, while Normanno et al. [40] and Huber et al. [41] confirmed MRSA in dairy, meat, and RTE foods at variable rates. Beyond Europe, Weese et al. [42] detected MRSA in North American retail meats and linked isolates to both human- and livestock-associated clonal complexes, underscoring the zoonotic interface.
Regional studies support our outlet gradient: lapses in hygiene and infrastructure amplify contamination risk [43,44,45,46]. In Nigeria, Beshiru et al. [45] confirmed MRSA in RTE foods, raising consumer safety concerns, and in South Africa, Asiegbu et al. [47] found S. aureus in 31.8% of street-vended foods, highlighting informal markets as vulnerable nodes. In contrast, robust surveillance and cold-chain systems are associated with far lower rates. In Singapore, Aung et al. [48] detected MRSA in 2.2% of retail food isolates, and Zwe et al. [49] reported similarly low occurrence. Japan shows comparable patterns, with Kitai et al. [50] and Sato et al. [51] reporting consistently low MRSA prevalence in retail meats, attributed to strict hygiene and monitoring. These comparisons illustrate how infrastructural disparities shape exposure risk: informal or resource-limited markets remain persistent hotspots, whereas strong compliance frameworks can suppress MRSA in foods.
Biological plausibility for the category and outlet patterns is clear. Animal-derived foods often carry higher initial bioburden and undergo intensive handling, facilitating S. aureus survival and growth. By contrast, bakery items with low water activity and high sugar content naturally restrict S. aureus persistence. Operational factors—time to sale, temperature control, and surface sanitation—already shown to be inconsistent in Saudi food-service settings [43]—likely contribute to elevated contamination in street markets. These observations argue for matrix- and outlet-specific surveillance that prioritizes high-risk products and environments, with benchmarking against low-prevalence regions to demonstrate achievable suppression [49].
Orthogonal confirmation strengthens confidence in our workflow and highlights RTE foods as reservoirs of resistant staphylococci. The 6.7% MRSA prevalence observed here aligns with international reports: Somda et al. [52] found S. aureus in 70.8% of RTE foods in Burkina Faso with 14.3% MRSA; Islam et al. [53] identified MRSA in ~23% of Bangladeshi isolates; and Saber et al. [54] confirmed mecA-positive multidrug-resistant S. aureus in Egyptian RTE meats. MALDI-TOF MS provided rapid, accurate species confirmation and revealed two reproducible low-mass peaks (m/z 3990 and 4125) associated with MRSA in our dataset. Similar discriminatory features have been reported [55,56], and machine learning classifiers have achieved >87% MRSA–MSSA discrimination, though performance may vary by SCCmec type [57]. Practically, MALDI-based signatures are best used to triage isolates for targeted molecular confirmation, while mecA/mecC PCR remains indispensable.
AST profiles showed the classic β-lactam divide: MRSA remained non-susceptible to oxacillin/cefoxitin, whereas MSSA was oxacillin-susceptible but largely penicillin-resistant—consistent with MENA and global food-chain data [58]. To avoid clindamycin failure, routine screening for inducible macrolide–lincosamide–streptogramin B (MLSB) resistance by the CLSI-recommended D-test remains essential, given known discordance between erythromycin and clindamycin phenotypes [59]. Non-β-lactam trends also matched global reports: MRSA frequently resisted ciprofloxacin and macrolides, while glycopeptides and oxazolidinones were preserved [60]. Regionally, surveillance from Saudi Arabia and the UAE shows similar pressures in animal-derived RTE items and vendor settings [61]. Our automated system (Vitek 2, AST-GP71) demonstrated high categorical agreement, in line with multicenter evaluations (~98% CA; very-major-error ≤ 0.5%) [16]. Even so, confirmatory D-testing and mecA/mecC PCR remain prudent for critical decisions [62]. Accordingly, coupling automated AST with targeted molecular confirmation enables scalable food-chain surveillance and stewardship that reserves last-line agents for confirmed need.
Against confirmed food-derived MRSA, citrate-stabilized AgNPs showed consistent inhibitory potency (MIC 8–32 µg/mL; MIC50 16; MIC90 32) and were predominantly bactericidal (MBC/MIC ≤ 4 in 90%). Comparable ranges appear across diverse formulations: mycosynthesized particles (~27.7 nm) yielded MICs ~25 µg/mL [63]; green-synthesized Curcuma longa AgNPs produced low MICs [64]; and more resistant S. aureus required ~50/100 µg/mL MIC/MBC [65]. Because particle size, surface chemistry, and Ag+ release kinetics strongly determine potency, standardized reporting of size distributions, ζ-potential, and ion release is critical for cross-study comparison and risk assessment [66]. Our narrow MIC distribution suggests relatively consistent activity across strains, with a single isolate showing a higher MBC/MIC ratio of eight. This variability underscores the biological heterogeneity of S. aureus and the need to balance antimicrobial efficacy with safety and cost in translational applications [67].
Combination testing indicated that AgNPs can re-sensitize MRSA to a β-lactam scaffold. Checkerboards showed AgNP–oxacillin synergy in 60% and additivity in 40% of isolates (median FICI 0.37), with 4–16-fold reductions in oxacillin MIC and 2–8-fold reductions in AgNP MIC; AgNP–ciprofloxacin was additive-to-synergistic (median FICI 0.63) with no antagonism. Independent reports similarly describe AgNP–antibiotic potentiation against MRSA, e.g., nanosilver plus conventional agents yielding synergy and rapid killing [22], and enhanced effects in multimodality regimens (AgNPs + blue light + antibiotics) [24]. Mechanistically, silver’s multitarget action—inhibiting multiple essential proteins and pathways—provides a plausible basis for β-lactam re-sensitization and a higher barrier to stable resistance [21]. PK/PD-guided optimization of dosing ratios and schedules in food-relevant matrices is warranted to translate static synergy into robust operational interventions.
Time–kill assays confirmed that adding AgNPs converts oxacillin from static or failing monotherapy into a rapidly bactericidal regimen. At MIC-matched, fixed concentrations, AgNPs plus oxacillin achieved ≥3-log10 kills by 24 h in all four tested isolates (two reached the limit of detection), while monotherapies produced only stasis or ~0.5–1.2-log10 declines. AgNPs plus ciprofloxacin showed additive-to-synergistic trajectories with earlier onset of killing and suppressed regrowth, without antagonism—consistent with prior reports of accelerated kill and reduced rebound under combination exposure [22,24]. Together with the checkerboard data, these results support the practical value of combinations, contingent on dosing that maintains concentrations above synergy-enabling thresholds.
Experimental evolution under sub-MIC silver produced modest, often reversible, increases in AgNP MICs (median two-fold; IQR 1–4-fold), whereas co-exposure with oxacillin constrained drift and returned isolates to baseline after drug withdrawal. This pattern aligns with evidence that silver adaptation can emerge under specific exposure designs—sometimes via nanoparticle aggregation or altered envelope responses—but is not universally stable [26,68]. We detected no consistent collateral shifts in oxacillin or ciprofloxacin MICs; however, metal–antibiotic co-selection remains a credible risk in food and environmental settings. Longer-term surveillance should track ARG/HMRG linkages and fitness trade-offs [28,69].
Overall, our findings position RTE foods as credible MRSA exposure pathways—particularly animal-derived and street-vended items—and argue for matrix- and outlet-aware surveillance, strengthened hygiene/cold-chain controls, and scalable confirmation workflows (MALDI triage → automated AST → mecA/mecC PCR). AgNPs emerge as promising adjuvants that restore β-lactam efficacy and deliver rapid bactericidal activity, while combination use limits silver-specific MIC drift. Before field deployment, PK/PD optimization, standardized AgNP characterization, toxicity/residue assessments, and regulatory alignment are essential [21,66].
5. Limitations of the Study
This single-region, cross-sectional survey (n = 149) limits generalizability, seasonality assessment, and outlet-level inference; the MRSA subset (n = 10) constrains power for subgroup analyses. MALDI-TOF MRSA-associated peaks were identified in a small dataset without external or interlaboratory validation and should be treated as triage cues pending broader verification. AST relied primarily on Vitek 2 rather than comprehensive BMD across all agents, and SCCmec/clone-level correlates were not resolved. Whole-genome sequencing of evolved lines was not performed, limiting mechanistic insight into silver-response pathways and potential co-selection. Baseline and endpoint aliquots were preserved, and a follow-up is pre-specified to sequence the two isolates with the largest durable MIC increases alongside their ancestors. Formal fitness assays were beyond scope. Finally, safety, residue, and regulatory considerations (cytotoxicity, ecotoxicity, and compliance for food-contact materials) were not evaluated and require dedicated study.
6. Conclusions
Across 149 RTE foods from Al-Qassim, S. aureus occurred in 24.2% and MRSA in 6.7%, concentrated in animal-derived items and street-vendor outlets. An orthogonal workflow—culture, MALDI-TOF MS, Vitek 2, and mecA/mecC PCR—delivered rapid, reliable confirmation; reproducible low-mass MALDI peaks supported triage, while PCR provided definitive calls. Susceptibility testing showed the expected β-lactam split with preserved activity of last-line agents, supporting automated AST augmented by targeted D-testing and mec gene confirmation. Citrate-stabilized AgNPs were consistently potent (MIC 8–32 µg/mL; MIC50 16; MIC90 32) and predominantly bactericidal, and in combination produced frequent synergy with oxacillin and additive-to-synergistic effects with ciprofloxacin that translated into rapid, sustained killing without antagonism. Sub-MIC evolution selected only modest, often reversible increases in silver MIC that were curtailed by β-lactam co-exposure. These findings justify matrix- and outlet-aware surveillance, support MALDI-assisted triage integrated with automated AST and mecA/mecC PCR, and nominate standardized AgNP formulations as promising β-lactam adjuvants for food-adjacent risk reduction, pending in situ efficacy, safety, and residue evaluation. Future work will incorporate whole-genome sequencing of selected evolved pairs to clarify genetic drivers of silver susceptibility changes and interactions with β-lactam exposure.
Supplementary Materials
The following supporting information can be downloaded at: https://www.mdpi.com/article/10.3390/microorganisms13102393/s1, Table S1: Isolate-level checkerboard interactions of silver nanoparticles (AgNPs) with oxacillin and ciprofloxacin against MRSA; Table S2: Time–kill outcomes for silver nanoparticle (AgNP) combinations against MRSA (subset, n = 4).
Author Contributions
Conceptualization, A.A. and E.M.; Methodology, A.A. and E.M.; writing—original draft preparation, A.A. and E.M.; visualization, A.A. and E.M.; supervision, A.A. and E.M.; project administration, A.A. and E.M.; funding acquisition, A.A. and E.M. All authors have read and agreed to the published version of the manuscript.
Funding
The researchers would like to thank the Deanship of Graduate Studies and Scientific Research at Qassim University for financial support (QU-APC-2025).
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
The original contributions presented in this study are included in the article and Supplementary Material. Further inquiries can be directed to the corresponding author.
Conflicts of Interest
The authors declare no conflicts of interest.
References
- Roy, M.C.; Chowdhury, T.; Hossain, M.T.; Hasan, M.M.; Zahran, E.; Rahman, M.M.; Zinnah, K.M.A.; Rahman, M.M.; Hossain, F.M.A. Zoonotic linkage and environmental contamination of Methicillin-resistant Staphylococcus aureus (MRSA) in dairy farms: A one health perspective. One Health 2024, 18, 100680. [Google Scholar] [CrossRef] [PubMed]
- World Health Organization (WHO). WHO Bacterial Priority Pathogens List, 2024: Bacterial Pathogens of Public Health Importance to Guide Research, Development and Strategies to Prevent and Control Antimicrobial Resistance; World Health Organization: Geneva, Switzerland, 2024.
- Wu, S.; Huang, J.; Zhang, F.; Wu, Q.; Zhang, J.; Pang, R.; Zeng, H.; Yang, X.; Chen, M.; Wang, J. Prevalence and characterization of food-related methicillin-resistant Staphylococcus aureus (MRSA) in China. Front. Microbiol. 2019, 10, 304. [Google Scholar] [CrossRef]
- Esemu, S.N.; Njoh, S.T.; Ndip, L.M.; Keneh, N.K.; Kfusi, J.A.; Njukeng, A.P. Ready-to-Eat Foods: A Potential Vehicle for the Spread of Coagulase-Positive Staphylococci and Antimicrobial-Resistant Staphylococcus aureus in Buea Municipality, South West Cameroon. Can. J. Infect. Dis. Med. Microbiol. 2023, 2023, 9735319. [Google Scholar] [CrossRef] [PubMed]
- Léguillier, V.; Pinamonti, D.; Chang, C.-M.; Mukherjee, R.; Cossetini, A.; Manzano, M.; Anba-Mondoloni, J.; Malet-Villemagne, J.; Vidic, J. A review and meta-analysis of Staphylococcus aureus prevalence in foods. Microbe 2024, 4, 100131. [Google Scholar] [CrossRef]
- Xing, L.; Cheng, M.; Jiang, J.; Li, T.; Zhang, X.; Tian, Y.; Liu, W. Methicillin-resistant Staphylococcus aureus contamination in meat and meat products: A systematic review and meta-analysis. Front. Microbiol. 2025, 16, 1636622. [Google Scholar] [CrossRef]
- Liang, T.; Liang, Z.; Wu, S.; Ding, Y.; Wu, Q.; Gu, B. Global prevalence of Staphylococcus aureus in food products and its relationship with the occurrence and development of diabetes mellitus. Med. Adv. 2023, 1, 53–78. [Google Scholar] [CrossRef]
- EFSA/ECDC. The European Union summary report on antimicrobial resistance in zoonotic and indicator bacteria from humans, animals and food in 2022–2023. EFSA J. 2025, 23, e9237. [Google Scholar]
- Sergelidis, D.; Angelidis, A.S. Methicillin-resistant Staphylococcus aureus: A controversial food-borne pathogen. Lett. Appl. Microbiol. 2017, 64, 409–418. [Google Scholar] [CrossRef]
- Larsen, J.; Stegger, M.; Andersen, P.S.; Petersen, A.; Larsen, A.R.; Westh, H.; Agersø, Y.; Fetsch, A.; Kraushaar, B.; Käsbohrer, A. Evidence for human adaptation and foodborne transmission of livestock-associated methicillin-resistant Staphylococcus aureus. Clin. Infect. Dis. 2016, 63, 1349–1352. [Google Scholar] [CrossRef]
- Murray, C.J.; Ikuta, K.S.; Sharara, F.; Swetschinski, L.; Aguilar, G.R.; Gray, A.; Han, C.; Bisignano, C.; Rao, P.; Wool, E. Global burden of bacterial antimicrobial resistance in 2019: A systematic analysis. Lancet 2022, 399, 629–655, Erratum in Lancet 2022, 400, 1102. [Google Scholar] [CrossRef]
- Naghavi, M.; Vollset, S.E.; Ikuta, K.S.; Swetschinski, L.R.; Gray, A.P.; Wool, E.E.; Aguilar, G.R.; Mestrovic, T.; Smith, G.; Han, C. Global burden of bacterial antimicrobial resistance 1990–2021: A systematic analysis with forecasts to 2050. Lancet 2024, 404, 1199–1226. [Google Scholar] [CrossRef]
- Cassini, A.; Högberg, L.D.; Plachouras, D.; Quattrocchi, A.; Hoxha, A.; Simonsen, G.S.; Colomb-Cotinat, M.; Kretzschmar, M.E.; Devleesschauwer, B.; Cecchini, M. Attributable deaths and disability-adjusted life-years caused by infections with antibiotic-resistant bacteria in the EU and the European Economic Area in 2015: A population-level modelling analysis. Lancet Infect. Dis. 2019, 19, 56–66. [Google Scholar] [CrossRef]
- Hu, Y.; Huang, Y.; Lizou, Y.; Li, J.; Zhang, R. Evaluation of Staphylococcus aureus subtyping module for methicillin-resistant Staphylococcus aureus detection based on matrix-assisted laser desorption ionization time-of-flight mass spectrometry. Front. Microbiol. 2019, 10, 2504. [Google Scholar] [CrossRef]
- Yu, J.; Tien, N.; Liu, Y.-C.; Cho, D.-Y.; Chen, J.-W.; Tsai, Y.-T.; Huang, Y.-C.; Chao, H.-J.; Chen, C.-J. Rapid identification of methicillin-resistant Staphylococcus aureus using MALDI-TOF MS and machine learning from over 20,000 clinical isolates. Microbiol. Spectr. 2022, 10, e00483-22. [Google Scholar] [CrossRef]
- Bobenchik, A.M.; Hindler, J.A.; Giltner, C.L.; Saeki, S.; Humphries, R.M. Performance of Vitek 2 for antimicrobial susceptibility testing of Staphylococcus spp. and Enterococcus spp. J. Clin. Microbiol. 2014, 52, 392–397. [Google Scholar] [CrossRef]
- Elbehiry, A.; Al-Dubaib, M.; Marzouk, E.; Osman, S.; Edrees, H. Performance of MALDI biotyper compared with Vitek™ 2 compact system for fast identification and discrimination of Staphylococcus species isolated from bovine mastitis. MicrobiologyOpen 2016, 5, 1061–1070. [Google Scholar] [CrossRef]
- Lakhundi, S.; Zhang, K. Methicillin-resistant Staphylococcus aureus: Molecular characterization, evolution, and epidemiology. Clin. Microbiol. Rev. 2018, 31, e00020-18. [Google Scholar] [CrossRef]
- García-Álvarez, L.; Holden, M.T.; Lindsay, H.; Webb, C.R.; Brown, D.F.; Curran, M.D.; Walpole, E.; Brooks, K.; Pickard, D.J.; Teale, C. Meticillin-resistant Staphylococcus aureus with a novel mecA homologue in human and bovine populations in the UK and Denmark: A descriptive study. Lancet Infect. Dis. 2011, 11, 595–603. [Google Scholar] [CrossRef] [PubMed]
- CDC. Laboratory Testing for Methicillin (Oxacillin)-Resistant Staphylococcus aureus (MRSA). 2025. Available online: https://www.cdc.gov/mrsa/php/laboratories/index.html (accessed on 17 September 2025).
- Wang, H.; Wang, M.; Xu, X.; Gao, P.; Xu, Z.; Zhang, Q.; Li, H.; Yan, A.; Kao, R.Y.-T.; Sun, H. Multi-target mode of action of silver against Staphylococcus aureus endows it with capability to combat antibiotic resistance. Nat. Commun. 2021, 12, 3331, Erratum in Nat. Commun. 2021, 12, 4140. [Google Scholar] [CrossRef] [PubMed]
- Rahim, K.A.A.A.; Mohamed, A.M.A. Bactericidal and antibiotic synergistic effect of nanosilver against methicillin-resistant Staphylococcus aureus. Jundishapur J. Microbiol. 2015, 8, e25867. [Google Scholar] [CrossRef] [PubMed]
- Casals, E.; Gusta, M.F.; Bastus, N.; Rello, J.; Puntes, V. Silver Nanoparticles and Antibiotics: A Promising Synergistic Approach to Multidrug-Resistant Infections. Microorganisms 2025, 13, 952. [Google Scholar] [CrossRef]
- Akram, F.E.; El-Tayeb, T.; Abou-Aisha, K.; El-Azizi, M. A combination of silver nanoparticles and visible blue light enhances the antibacterial efficacy of ineffective antibiotics against methicillin-resistant Staphylococcus aureus (MRSA). Ann. Clin. Microbiol. Antimicrob. 2016, 15, 48. [Google Scholar] [CrossRef]
- Castro, I.M.d.; Antunes, C.; Valentim, C.C.; Spoladori, L.F.d.A.; Suzukawa, H.T.; Correia, G.F.; Silva-Rodrigues, G.; Borges, P.H.G.; Bartolomeu-Gonçalves, G.; Silva, M.L. Synergistic antibacterial interaction of geraniol and biogenic silver nanoparticles on methicillin-resistant Staphylococcus aureus. Plants 2025, 14, 1059. [Google Scholar] [CrossRef]
- Hochvaldová, L.; Panáček, D.; Válková, L.; Večeřová, R.; Kolář, M.; Prucek, R.; Kvítek, L.; Panáček, A.E. coli and S. aureus resist silver nanoparticles via an identical mechanism, but through different pathways. Commun. Biol. 2024, 7, 1552. [Google Scholar] [CrossRef]
- Panáček, D.; Hochvaldová, L.; Bakandritsos, A.; Malina, T.; Langer, M.; Belza, J.; Martincová, J.; Večeřová, R.; Lazar, P.; Poláková, K. Silver covalently bound to cyanographene overcomes bacterial resistance to silver nanoparticles and antibiotics. Adv. Sci. 2021, 8, 2003090. [Google Scholar] [CrossRef]
- Gillieatt, B.F.; Coleman, N.V. Unravelling the mechanisms of antibiotic and heavy metal resistance co-selection in environmental bacteria. FEMS Microbiol. Rev. 2024, 48, fuae017. [Google Scholar] [CrossRef] [PubMed]
- Clinical and Laboratory Standards Institute (CLSI). Performance Standards for Antimicrobial Susceptibility Testing; CLSI: Malvern, PA, USA, 2025. [Google Scholar]
- European Committee on Antimicrobial Susceptibility Testing (EUCAST). Breakpoint Tables for Interpretation of MICs and Zone Diameters; EUCAST: Växjö, Sweden, 2025. [Google Scholar]
- Stegger, á.; Andersen, P.; Kearns, A.; Pichon, B.; Holmes, M.; Edwards, G.; Laurent, F.; Teale, C.; Skov, R.; Larsen, A. Rapid detection, differentiation and typing of methicillin-resistant Staphylococcus aureus harbouring either mecA or the new mecA homologue mecALGA251. Clin. Microbiol. Infect. 2012, 18, 395–400. [Google Scholar] [CrossRef] [PubMed]
- DTU Food, National Food Institute (DTU/NFI). MRSA Multiplex PCR-2 Protocol; Technical University of Denmark: Kongens Lyngby, Denmark, 2024. [Google Scholar]
- Strommenger, B.; Braulke, C.; Heuck, D.; Schmidt, C.; Pasemann, B.; Nubel, U.; Witte, W. spa typing of Staphylococcus aureus as a frontline tool in epidemiological typing. J. Clin. Microbiol. 2008, 46, 574–581. [Google Scholar] [CrossRef] [PubMed]
- ISO 22412; Particle Size Analysis—Dynamic Light Scattering (DLS). ISO: Geneva, Switzerland, 2025.
- ISO 13099-1; Colloidal Systems—Methods for Zeta-Potential Determination. Part 1: Electroacoustic and Electrokinetic Phenomena. ISO: Geneva, Switzerland, 2012.
- ISO 17294-2; Water Quality—Application of Inductively Coupled Plasma Mass Spectrometry (ICP-MS). Part 2: Determination of Selected Elements Including Uranium Isotopes. ISO: Geneva, Switzerland, 2023.
- Wan Mat Khalir, W.K.A.; Shameli, K.; Jazayeri, S.D.; Othman, N.A.; Che Jusoh, N.W.; Hassan, N.M. Biosynthesized silver nanoparticles by aqueous stem extract of Entada spiralis and screening of their biomedical activity. Front. Chem. 2020, 8, 620. [Google Scholar] [CrossRef]
- Wang, W.; Baloch, Z.; Jiang, T.; Zhang, C.; Peng, Z.; Li, F.; Fanning, S.; Ma, A.; Xu, J. Enterotoxigenicity and antimicrobial resistance of Staphylococcus aureus isolated from retail food in China. Front. Microbiol. 2017, 8, 2256. [Google Scholar] [CrossRef] [PubMed]
- Basanisi, M.; La Bella, G.; Nobili, G.; Tola, S.; Cafiero, M.; La Salandra, G. Prevalence and characterization of methicillin-resistant Staphylococcus aureus (MRSA) isolates from retail meat in south Italy. Ital. J. Food Sci. 2020, 32, 410–419. [Google Scholar]
- Normanno, G.; Corrente, M.; La Salandra, G.; Dambrosio, A.; Quaglia, N.C.; Parisi, A.; Greco, G.; Bellacicco, A.; Virgilio, S.; Celano, G.V. Methicillin-resistant Staphylococcus aureus (MRSA) in foods of animal origin product in Italy. Int. J. Food Microbiol. 2007, 117, 219–222. [Google Scholar] [CrossRef] [PubMed]
- Huber, H. Prevalence and Characteristics of Meticillin-Resistant Staphylococcus aureus in Humans in Contact with Farm Animals, in Livestock, and in Food of Animal Origin, Switzerland, 2009; University of Zurich: Zurich, Switzerland, 2010. [Google Scholar]
- Weese, J.; Avery, B.; Reid-Smith, R. Detection and quantification of methicillin-resistant Staphylococcus aureus (MRSA) clones in retail meat products. Lett. Appl. Microbiol. 2010, 51, 338–342. [Google Scholar] [CrossRef] [PubMed]
- Bukhari, M.A.; Banasser, T.M.; El-Bali, M.; Bulkhi, R.A.; Qamash, R.A.; Trenganno, A.; Khayyat, M.; Kurdi, M.A.; Al Majrashi, A.; Bahewareth, F. Assessment of microbiological quality of food preparation process in some restaurants of Makkah city. Saudi J. Biol. Sci. 2021, 28, 5993–5997. [Google Scholar] [CrossRef] [PubMed]
- Moges, M.; Rodland, E.K.; Ambelu, A. Health risk assessment of Staphylococcus aureus and Salmonella from the consumption of street foods in Ethiopia. BMC Infect. Dis. 2025, 25, 576. [Google Scholar] [CrossRef]
- Beshiru, A.; Isichei-Ukah, B.O.; Uwhuba, K.E.; Igere, B.E.; Igbinosa, E.O. Prevalence, characterization, and implications of methicillin-resistant Staphylococcus aureus (MRSA) in ready-to-eat foods from Delta, Nigeria: A concern for consumer safety. Sustain. Microbiol. 2024, 1, qvae007. [Google Scholar] [CrossRef]
- Fusaro, C.; Miranda-Madera, V.; Serrano-Silva, N.; Bernal, J.E.; Ríos-Montes, K.; González-Jiménez, F.E.; Ojeda-Juárez, D.; Sarria-Guzmán, Y. Antibiotic-resistant bacteria isolated from street foods: A systematic review. Antibiotics 2024, 13, 481. [Google Scholar] [CrossRef] [PubMed]
- Asiegbu, C.; Lebelo, S.; Tabit, F. Microbial quality of ready-to-eat street vended food groups sold in the Johannesburg Metropolis, South Africa. J. Food Qual. Hazards Control 2020, 1, qvae007. [Google Scholar] [CrossRef]
- Aung, K.T.; Hsu, L.Y.; Koh, T.H.; Hapuarachchi, H.C.; Chau, M.L.; Gutiérrez, R.A.; Ng, L.C. Prevalence of methicillin-resistant Staphylococcus aureus (MRSA) in retail food in Singapore. Antimicrob. Resist. Infect. Control 2017, 6, 94. [Google Scholar] [CrossRef]
- Zwe, Y.H.; Mohan, R.D.; Lim, S.J.; Lai, W.C.; Sim, K.H.; Leyau, Y.L.; Lew, K.; Chua, J.M.C.; Aung, K.T.; Chng, K.R. Occurrence & characterization of Staphylococcus aureus from ready-to-eat (RTE), and cooked food in Singapore: A retrospective analysis. Int. J. Food Microbiol. 2025, 436, 111213. [Google Scholar]
- Kitai, S.; Shimizu, A.; Kawano, J.; Sato, E.; Nakano, C.; Uji, T.; Kitagawa, H. Characterization of methicillin-resistant Staphylococcus aureus isolated from retail raw chicken meat in Japan. J. Vet. Med. Sci. 2005, 67, 107–110. [Google Scholar] [CrossRef]
- Sato, T.; Usui, M.; Konishi, N.; Kai, A.; Matsui, H.; Hanaki, H.; Tamura, Y. Closely related methicillin-resistant Staphylococcus aureus isolates from retail meat, cows with mastitis, and humans in Japan. PLoS ONE 2017, 12, e0187319. [Google Scholar] [CrossRef]
- Somda, N.S.; Traoré, A.M.E.; Hien, D.F.d.S.; Bockarie, Y.; Tankoano, A.; Kaboré, D.; Bonkoungou, O.J.I.; Sawadogo-Lingani, H.; Savadogo, A. Molecular characterization of Methicillin-resistant Staphylococcus aureus isolated in ready-to-eat food sold in supermarkets in Bobo-Dioulasso: Case of charcuterie products. BMC Infect. Dis. 2024, 24, 722. [Google Scholar] [CrossRef]
- Islam, M.A.; Parveen, S.; Rahman, M.; Huq, M.; Nabi, A.; Khan, Z.U.M.; Ahmed, N.; Wagenaar, J.A. Occurrence and characterization of methicillin resistant Staphylococcus aureus in processed raw foods and ready-to-eat foods in an urban setting of a developing country. Front. Microbiol. 2019, 10, 503. [Google Scholar] [CrossRef]
- Saber, T.; Samir, M.; El-Mekkawy, R.M.; Ariny, E.; El-Sayed, S.R.; Enan, G.; Abdelatif, S.H.; Askora, A.; Merwad, A.M.; Tartor, Y.H. Methicillin-and vancomycin-resistant Staphylococcus aureus from humans and ready-to-eat meat: Characterization of antimicrobial resistance and biofilm formation ability. Front. Microbiol. 2022, 12, 735494. [Google Scholar] [CrossRef] [PubMed]
- Kim, J.-M.; Kim, I.; Chung, S.H.; Chung, Y.; Han, M.; Kim, J.-S. Rapid discrimination of methicillin-resistant Staphylococcus aureus by MALDI-TOF MS. Pathogens 2019, 8, 214. [Google Scholar] [CrossRef]
- Wang, Y.R.; Qian, C.; Cui, S.H.; Li, F.Q. Characterization of Staphylococcus aureus isolated from clinical specimens by matrix assisted laser desorption/ionization time-of-flight mass spectrometry. Biomed. Environ. Sci. 2013, 26, 430–436. [Google Scholar] [PubMed]
- Jeon, K.; Kim, J.-M.; Rho, K.; Jung, S.H.; Park, H.S.; Kim, J.-S. Performance of a machine learning-based methicillin resistance of Staphylococcus aureus identification system using MALDI-TOF MS and comparison of the accuracy according to SCC mec types. Microorganisms 2022, 10, 1903. [Google Scholar] [CrossRef] [PubMed]
- Alkuraythi, D.M.; Alkhulaifi, M.M. Methicillin-resistant Staphylococcus aureus prevalence in food-producing animals and food products in Saudi Arabia: A review. Vet. World 2024, 17, 1753. [Google Scholar] [CrossRef]
- Thapa, D.; Pyakurel, S.; Thapa, S.; Lamsal, S.; Chaudhari, M.; Adhikari, N.; Shrestha, D. Staphylococcus aureus with inducible clindamycin resistance and methicillin resistance in a tertiary hospital in Nepal. Trop. Med. Health 2021, 49, 99. [Google Scholar] [CrossRef]
- Islam, S.; Nasrin, N.; Tithi, N.S.; Khatun, F.; Asaduzzaman, M.; Topa, A.F.; Kabir, M.F.; Haque, F.K.M.; Jubair, M.; Rahman, M. Antimicrobial Susceptibility and Genomic Profiles of Multidrug-Resistant Staphylococcus aureus from Nasopharynx of Asymptomatic Children in Dhaka, Bangladesh. Life 2024, 14, 971. [Google Scholar] [CrossRef]
- Raji, M.A.; Garaween, G.; Ehricht, R.; Monecke, S.; Shibl, A.M.; Senok, A. Genetic characterization of Staphylococcus aureus isolated from retail meat in Riyadh, Saudi Arabia. Front. Microbiol. 2016, 7, 911. [Google Scholar] [CrossRef]
- Majhi, S.; Dash, M.; Mohapatra, D.; Mohapatra, A.; Chayani, N. Detection of inducible and constitutive clindamycin resistance among Staphylococcus aureus isolates in a tertiary care hospital, Eastern India. Avicenna J. Med. 2016, 6, 75–80. [Google Scholar] [CrossRef]
- Ali, E.M.; Rajendran, P.; Abdallah, B.M. Mycosynthesis of silver nanoparticles from endophytic Aspergillus parasiticus and their antibacterial activity against methicillin-resistant Staphylococcus aureus in vitro and in vivo. Front. Microbiol. 2024, 15, 1483637. [Google Scholar] [CrossRef]
- Do, B.L.; Bui, T.H.; Ho, T.G.-T.; Duong, N.L.; Nguyen, V.M.; Dang-Bao, T.; Nguyen, T.; Phuong, P.H. Green synthesis of nano-silver and its antibacterial activity against methicillin-resistant Staphylococcus aureus. J. Saudi Chem. Soc. 2023, 27, 101722. [Google Scholar]
- Elghazaly, E.M.; Torky, H.A.; Tawfik, R.G. Effect of silver nanoparticles and REP-PCR typing of Staphylococcus aureus isolated from various sources. Sci. Rep. 2024, 14, 21997. [Google Scholar] [CrossRef] [PubMed]
- He, X.; Hwang, H.-M. Nanotechnology in food science: Functionality, applicability, and safety assessment. J. Food Drug Anal. 2016, 24, 671–681. [Google Scholar] [CrossRef] [PubMed]
- Ansari, M.A.; Khan, H.M.; Khan, A.A.; Sultan, A.; Azam, A.; Shahid, M.; Shujatullah, F. Antibacterial activity of silver nanoparticles dispersion against MSSA and MRSA isolated from wounds in a tertiary care hospital of north India. Int. J. Appl. Biol. Pharm. Technol. 2011, 2, 34–42. [Google Scholar]
- Panáček, A.; Kvítek, L.; Smékalová, M.; Večeřová, R.; Kolář, M.; Röderová, M.; Dyčka, F.; Šebela, M.; Prucek, R.; Tomanec, O. Bacterial resistance to silver nanoparticles and how to overcome it. Nat. Nanotechnol. 2018, 13, 65–71. [Google Scholar] [CrossRef] [PubMed]
- Xu, Z.; Lin, X. Metal-regulated antibiotic resistance and its implications for antibiotic therapy. Microb. Biotechnol. 2024, 17, e14537. [Google Scholar] [CrossRef] [PubMed]




Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).